
Cyanobacterial Assemblages Inhabiting the Apatity Thermal Power Plant Fly Ash Dumps in the Russian Arctic


Abstract
1. Introduction
2. Materials and Methods
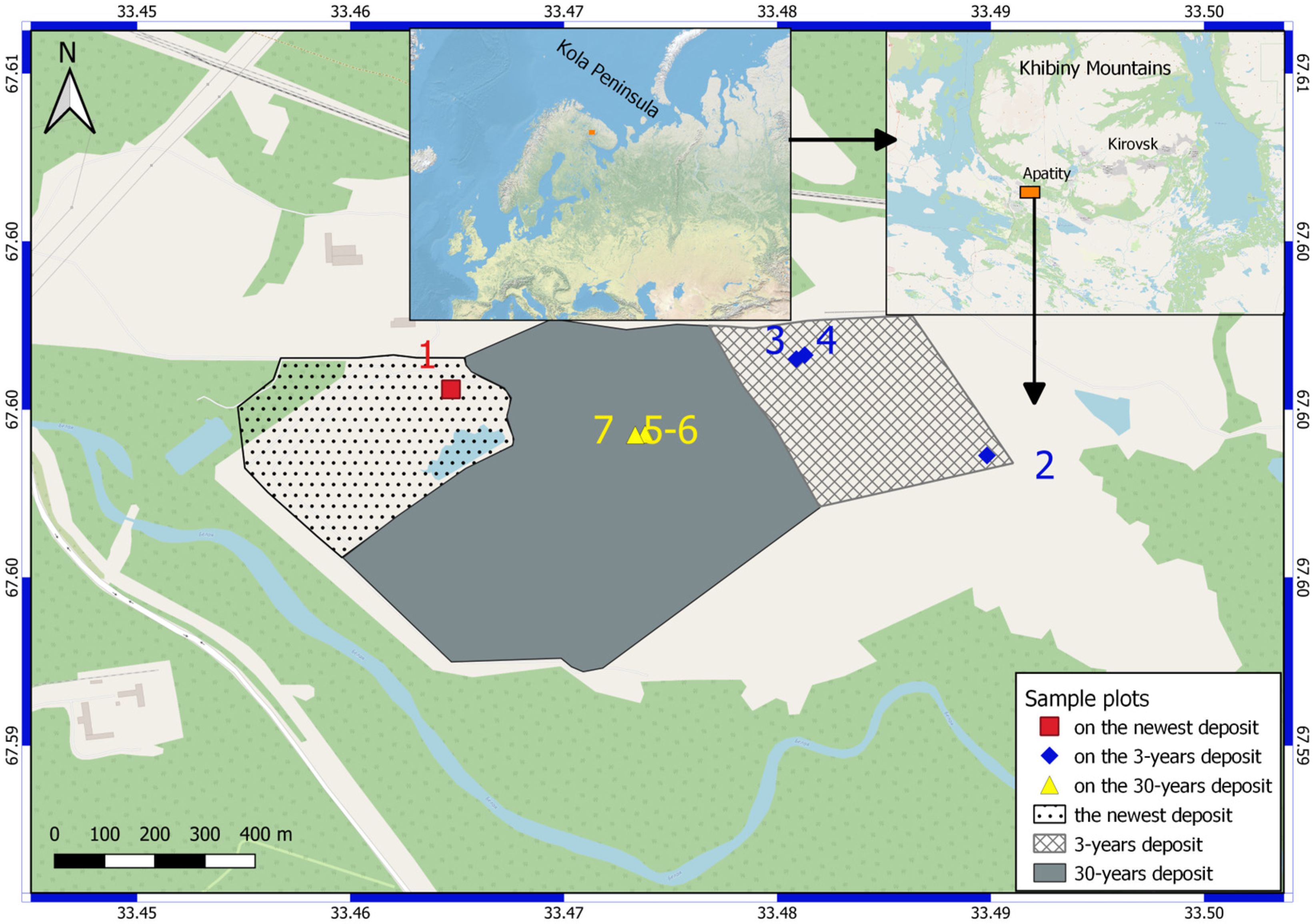
2.1. Description of Study Sites
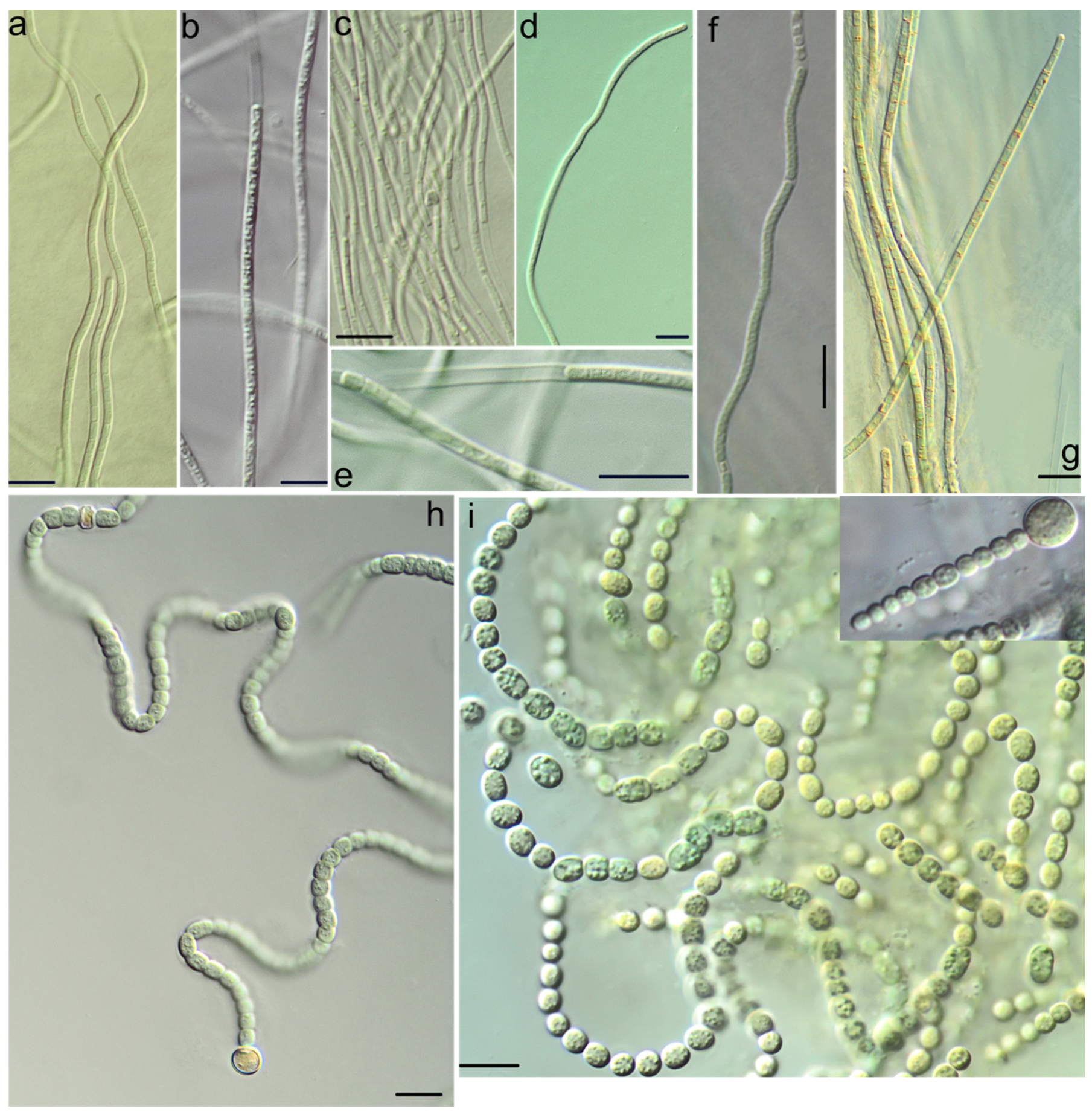
2.2. Culture Condition and Morphological Study
2.3. DNA Extraction and Sequencing
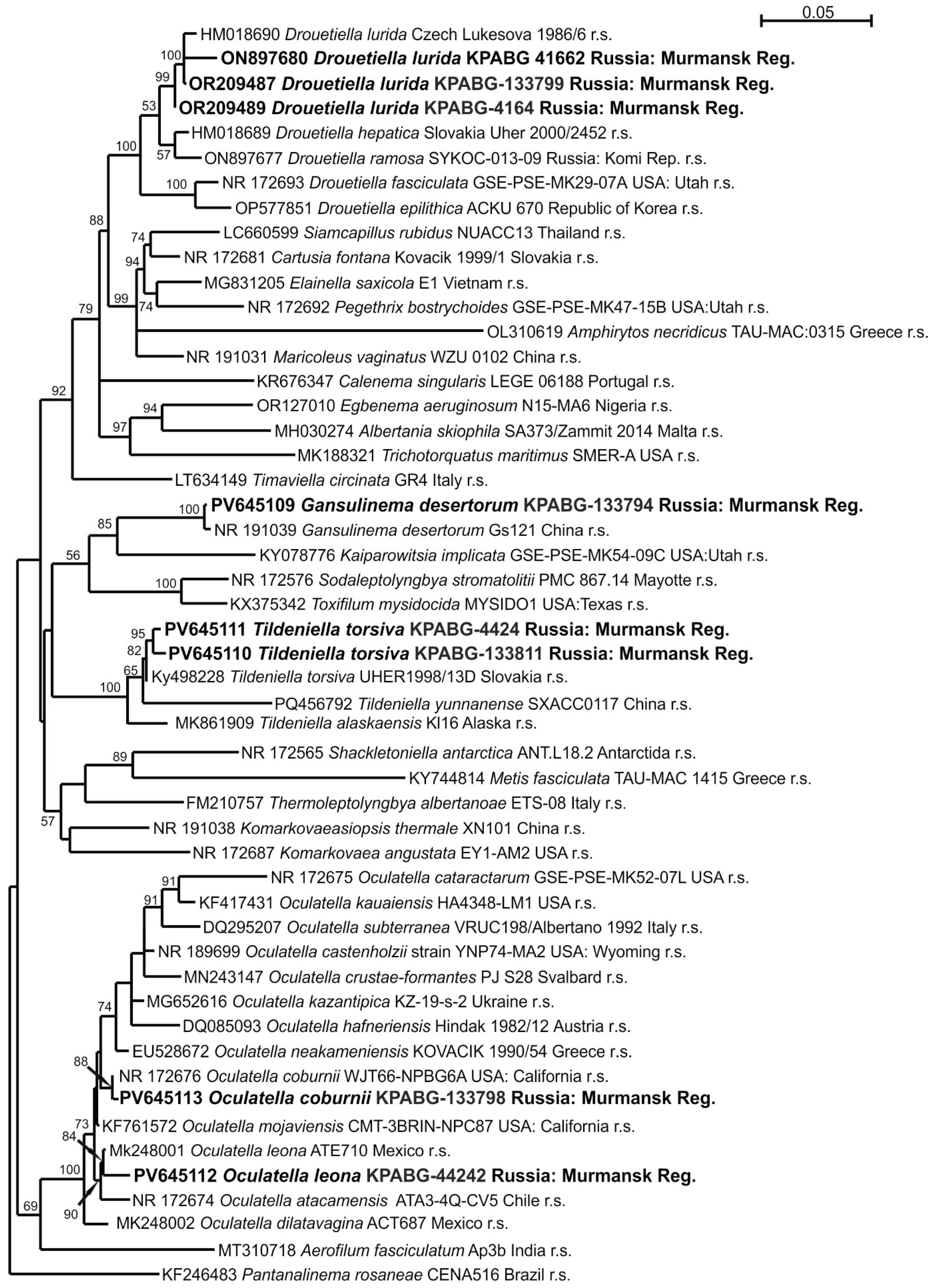
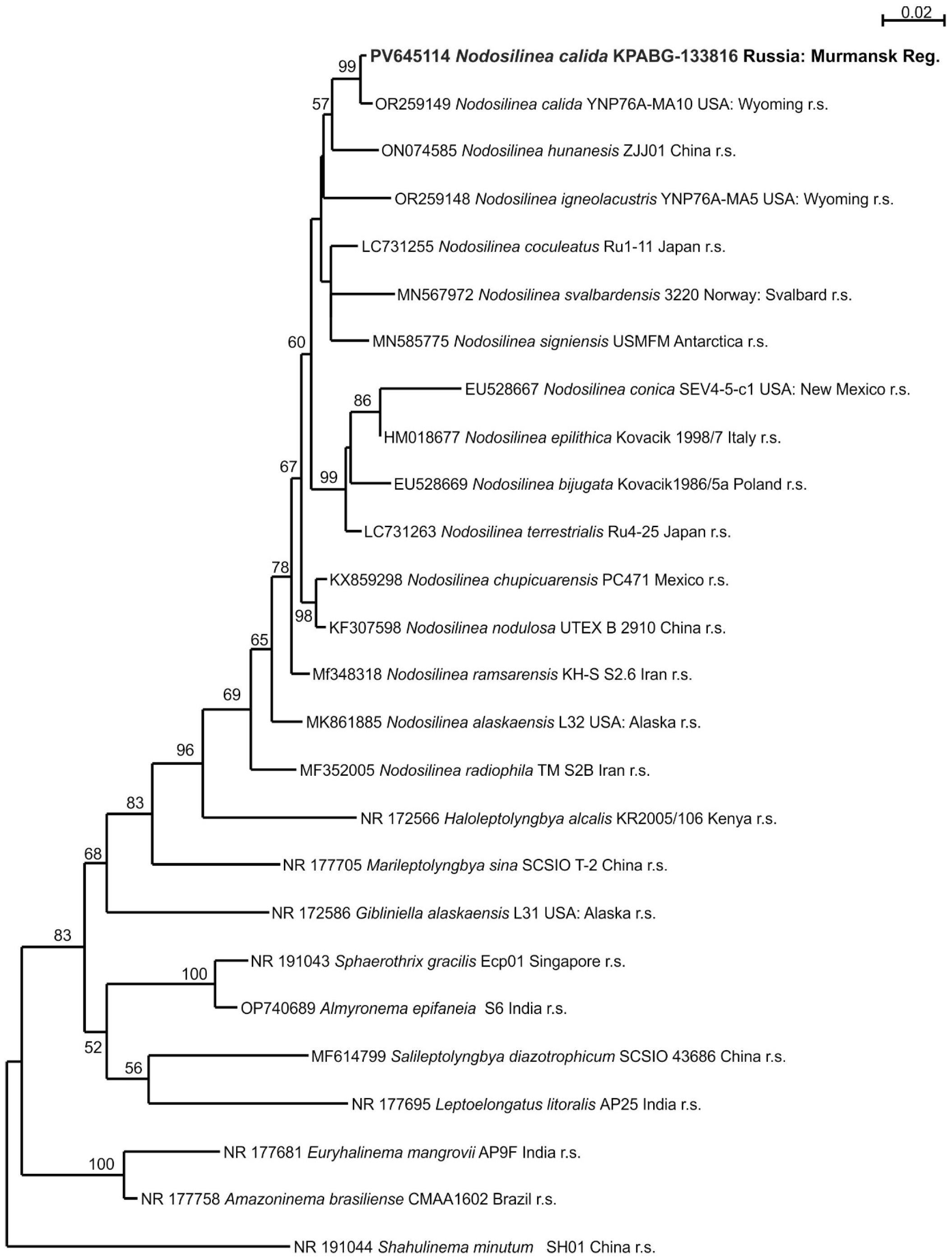
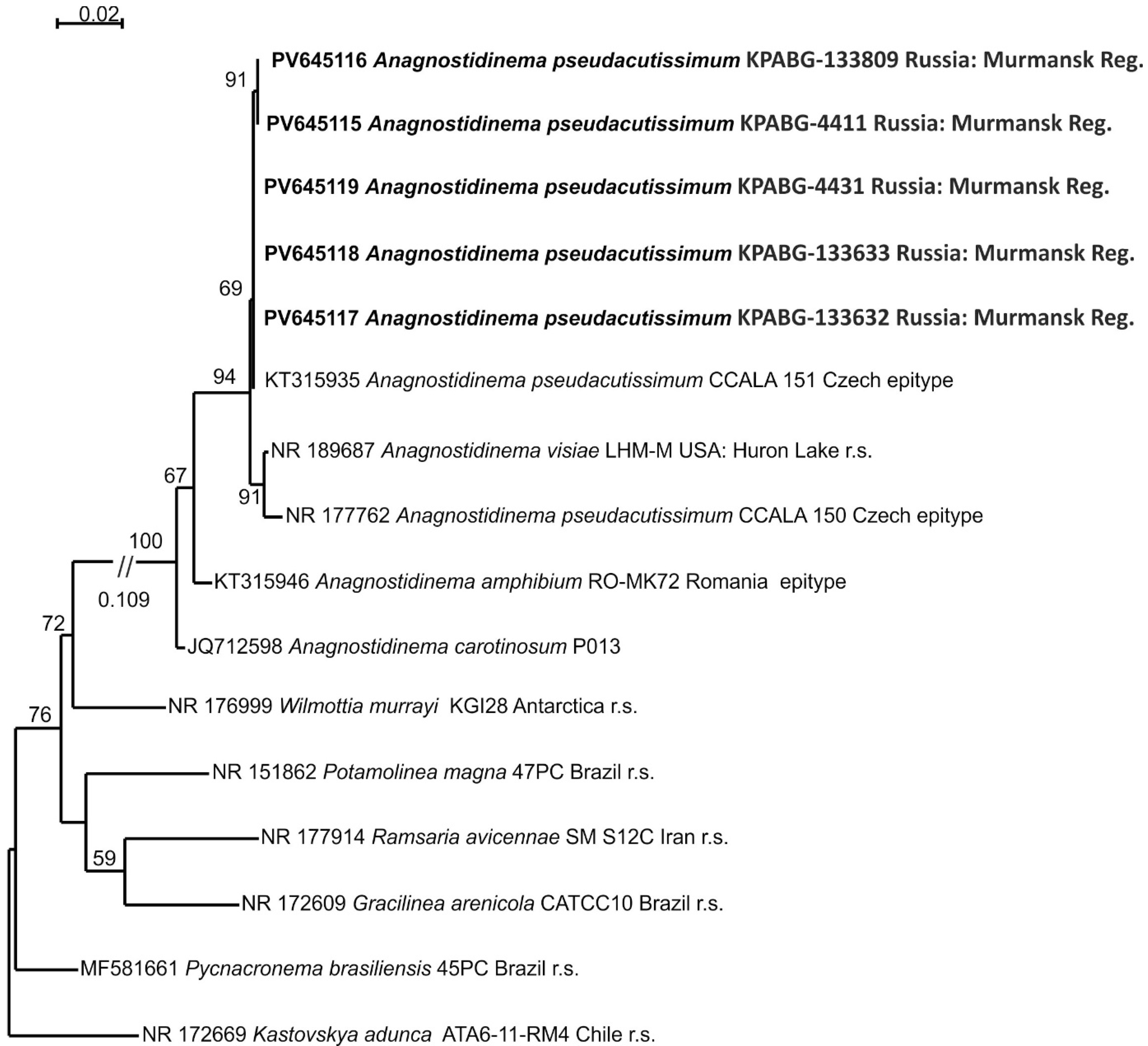
2.4. Molecular Phylogenetic Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Economic Forum. Available online: https://www.weforum.org/agenda/2019/02/these-countries-are-driving-global-demand-for-coal/ (accessed on 9 April 2025).
- Delić, D.; Stajković-Srbinović, O.; Buntić, A. Hazards and Usability of Coal Fly Ash. In Advances in Understanding Soil Degradation; Saljnikov, E., Mueller, L., Lavrishchev, A., Eulenstein, F., Eds.; Innovations in Landscape Research; Springer International Publishing: Cham, Switzerland, 2022; pp. 571–608. ISBN 978-3-030-85681-6. [Google Scholar]
- Menshov, P.V.; Khlupin, Y.V.; Nalesnik, O.I.; Makarovskikh, A.V. Ash and Slag Waste as a Secondary Raw Material. Procedia Chem. 2014, 10, 184–191. [Google Scholar] [CrossRef]
- Kryuchkov, V.V. Extreme Anthropogenic Loads and the Northern Ecosystem Condition. Ecol. Appl. 1993, 3, 622–630. [Google Scholar] [CrossRef]
- Slukovskaya, M.V.; Ivanova, L.A.; Kremenetskaya, I.P.; Gorbacheva, T.T.; Drogobuzhskaya, S.V.; Lashchuk, V.V.; Markovskaya, E.F. Rehabilitation of Industrial Barren in Arctic Region Using Mining Wastes. Open Ecol. J. 2018, 11, 1–13. [Google Scholar] [CrossRef]
- Chatterjee, S.; Kumari, S.; Rath, S.; Das, S. Prospects and Scope of Microbial Bioremediation for the Restoration of the Contaminated Sites. In Microbial Biodegradation and Bioremediation; Elsevier: Amsterdam, The Netherlands, 2022; pp. 3–31. ISBN 978-0-323-85455-9. [Google Scholar]
- Dudeja, C.; Masroor, S.; Mishra, V.; Kumar, K.; Sansar, S.; Yadav, P.; Chaturvedi, N.; Singh, R.; Kumar, A. Cyanobacteria-Based Bioremediation of Environmental Contaminants: Advances and Computational Insights. Discov. Agric. 2025, 3, 42. [Google Scholar] [CrossRef]
- Hassan, S.; Bhadwal, S.S.; Khan, M.; Sabreena; Nissa, K.-U.; Shah, R.A.; Bhat, H.M.; Bhat, S.A.; Lone, I.M.; Ganai, B.A. Revitalizing Contaminated Lands: A State-of-the-Art Review on the Remediation of Mine-Tailings Using Phytoremediation and Genomic Approaches. Chemosphere 2024, 356, 141889. [Google Scholar] [CrossRef]
- Kumar, A. Inorganic Soil Contaminants and Their Biological Remediation. In Plant Responses to Soil Pollution; Singh, P., Singh, S.K., Prasad, S.M., Eds.; Springer: Singapore, 2020; pp. 133–153. ISBN 978-981-15-4963-2. [Google Scholar]
- Rossi, F.; Mugnai, G.; De Philippis, R. Cyanobacterial Biocrust Induction: A Comprehensive Review on a Soil Rehabilitation-Effective Biotechnology. Geoderma 2022, 415, 115766. [Google Scholar] [CrossRef]
- Rossi, F.; Li, H.; Liu, Y.; De Philippis, R. Cyanobacterial Inoculation (Cyanobacterisation): Perspectives for the Development of a Standardized Multifunctional Technology for Soil Fertilization and Desertification Reversal. Earth-Sci. Rev. 2017, 171, 28–43. [Google Scholar] [CrossRef]
- Zanganeh, F.; Heidari, A.; Sepehr, A.; Rohani, A. Bioaugmentation and Bioaugmentation—Assisted Phytoremediation of Heavy Metal Contaminated Soil by a Synergistic Effect of Cyanobacteria Inoculation, Biochar, and Purslane (Portulaca oleracea L.). Environ. Sci. Pollut. Res. 2022, 29, 6040–6059. [Google Scholar] [CrossRef]
- Seiderer, T.; Venter, A.; Van Wyk, F.; Levanets, A.; Jordaan, A. Growth of Soil Algae and Cyanobacteria on Gold Mine Tailings Material. S. Afr. J. Sci. 2017, 113, 6. [Google Scholar] [CrossRef] [PubMed]
- Ssenku, J.E.; Ntale, M.; Backeus, I.; Lehtila, K.; Oryem-Origa, H. Dynamics of Plant Species during Phytostabilisation of Copper Mine Tailings and Pyrite Soils, Western Uganda. J. Environ. Eng. Ecol. Sci. 2014, 3, 4. [Google Scholar] [CrossRef]
- Davydov, D.A. Cyanoprokaryota and Their Role in the Process of Nitrogen Fixation in Terrestrial Ecosystems of the Murmansk Region; Geos: Moscow, Russia, 2010. [Google Scholar]
- Coppens, J.; Grunert, O.; Van Den Hende, S.; Vanhoutte, I.; Boon, N.; Haesaert, G.; De Gelder, L. The Use of Microalgae as a High-Value Organic Slow-Release Fertilizer Results in Tomatoes with Increased Carotenoid and Sugar Levels. J. Appl. Phycol. 2016, 28, 2367–2377. [Google Scholar] [CrossRef]
- Silva, G.H.R.; Sueitt, A.P.E.; Haimes, S.; Tripidaki, A.; Van Zwieten, R.; Fernandes, T.V. Feasibility of Closing Nutrient Cycles from Black Water by Microalgae-Based Technology. Algal Res. 2019, 44, 101715. [Google Scholar] [CrossRef]
- Kumar, J.; Singh, D.; Tyagi, M.B.; Kumar, A. Cyanobacteria: Applications in Biotechnology. In Cyanobacteria; Elsevier: Amsterdam, The Netherlands, 2019; pp. 327–346. ISBN 978-0-12-814667-5. [Google Scholar]
- FGBU “REA” Minenergo Rossii. Integrated Investment Project, Modernization of the Heat Supply System of the Murmansk Region for 2015–2030; FGBU “REA” Minenergo Rossii: Moscow, Russia, 2014. [Google Scholar]
- Pak, A.A. On the Issue of Using Technogenic Waste from Enterprises in the Murmansk Region in Cellular Concrete. Available online: https://helion-ltd.ru/use-of-tech-waste (accessed on 23 May 2020).
- Pak, A.A.; Sukhorukova, N.R. Use of Technogenic Waste from the Murmansk Region in Cellular Concrete. Proc. Fersman Sci. Sess. Kola Sci. Cent. Russ. Acad. Sci. 2017, 14, 254–257. [Google Scholar]
- Kozhukova, N.I.; Zhernovsky, I.V.; Fomina, E.V. Phase Formation in Geopolymer Systems Based on Fly Ash from the Apatite CHPP. Build. Mater. 2015, 85–88. [Google Scholar]
- Krasheninnikov, O.N.; Pak, A.A.; Bastrygina, S.V. Use of Ash Waste for the Production of Concrete. Ecol. Ind. Prod. 2007, 2, 48–56. [Google Scholar]
- VNIIG. Recommendations for the Design of Ash and Slag Dumps at Thermal Power Plants; VNIIG: St Petersburg, Russia, 1986. [Google Scholar]
- Kotai, J. Instructions for Preparation of Modified Nutrient Solution Z8 for Algae. Nor. Inst. Water Res. 1972, 11, 5. [Google Scholar]
- Komárek, J. Cyanoprokaryota 3. Teil: Heterocytous Genera; Süßwasserflora von Mitteleuropa; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 978-3-8274-0932-4. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Chroococcales; Cyanoprokaryota/Komárek, Jiří; Unaltered repr.; Spektrum Akad. Verl: Heidelberg, Germany, 2008; ISBN 978-3-7482-2111-1. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Oscillatoriales; Cyanoprokaryota/Komárek, Jiří; Unaltered repr., 2. print.; Spektrum Akad. Verl: Heidelberg, Germany, 2008; ISBN 978-3-8274-1914-9. [Google Scholar]
- Boyer, S.L.; Flechtner, V.R.; Johansen, J.R. Is the 16S–23S rRNA Internal Transcribed Spacer Region a Good Tool for Use in Molecular Systematics and Population Genetics? A Case Study in Cyanobacteria. Mol. Biol. Evol. 2001, 18, 1057–1069. [Google Scholar] [CrossRef] [PubMed]
- Neilan, B.A.; Stuart, J.L.; Goodman, A.E.; Cox, P.T.; Hawkins, P.R. Specific Amplification and Restriction Polymorphisms of the Cyanobacterial rRNA Operon Spacer Region. Syst. Appl. Microbiol. 1997, 20, 612–621. [Google Scholar] [CrossRef]
- Nübel, U.; Garcia-Pichel, F.; Muyzer, G. PCR Primers to Amplify 16S rRNA Genes from Cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. World-Wide Electronic Publication; AlgaeBase: Galway, Ireland, 2025. [Google Scholar]
- Strunecký, O.; Ivanova, A.P.; Mareš, J. An Updated Classification of Cyanobacterial Orders and Families Based on Phylogenomic and Polyphasic Analysis. J. Phycol. 2023, 59, 12–51. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.-H.; Whitman, W.B.; Euzéby, J.; Amann, R.; Rosselló-Móra, R. Uniting the Classification of Cultured and Uncultured Bacteria and Archaea Using 16S rRNA Gene Sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef]
- Strunecký, O.; Bohunická, M.; Johansen, J.R.; Čapková, K.; Raabová, L.; Dvořák, P.; Komárek, J. A Revision of the Genus Geitlerinema and a Description of the Genus Anagnostidinema Gen. Nov. (Oscillatorio phycidae, Cyanobacteria). Fottea 2017, 17, 114–126. [Google Scholar] [CrossRef]
- McGovern, C.A.; Norwich, A.R.; Thomas, A.L.; Hamsher, S.E.; Biddanda, B.A.; Weinke, A.D.; Casamatta, D.A. Unbiased Analyses of ITS Folding Motifs in a Taxonomically Confusing Lineage: Anagnostidinema visiae Sp. Nov. (Cyanobacteria). J. Phycol. 2023, 59, 619–634. [Google Scholar] [CrossRef]
- Hašler, P.; Dvořák, P.; Johansen, J.R.; Kitner, M.; Vladan, O.; Poulíčková, A. Morphological and Molecular Study of Epipelic Filamentous Genera Phormidium, Microcoleus and Geitlerinema (Oscillatoriales, Cyanophyta/Cyanobacteria). Fottea 2012, 12, 341–356. [Google Scholar] [CrossRef]
- Stanojković, A.; Skoupý, S.; Johannesson, H.; Dvořák, P. The Global Speciation Continuum of the Cyanobacterium Microcoleus. Nat. Commun. 2024, 15, 2122. [Google Scholar] [CrossRef]
- Strunecký, O.; Komárek, J.; Johansen, J.; Lukešová, A.; Elster, J. Molecular and Morphological Criteria for Revision of the Genus M Icrocoleus (Oscillatoriales, Cyanobacteria). J. Phycol. 2013, 49, 1167–1180. [Google Scholar] [CrossRef]
- Mishra, D.; Saraf, A.; Kumar, N.; Pal, S.; Singh, P. Issues in Cyanobacterial Taxonomy: Comprehensive Case Study of Unbranched, False Branched and True Branched Heterocytous Cyanobacteria. FEMS Microbiol. Lett. 2021, 368, fnab005. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Saraf, A.; Kumar, N.; Singh, P. Igniting Taxonomic Curiosity: The Amazing Story of Amazonocrinis with the Description of a New Genus Ahomia Gen. Nov. and Novel Species of Ahomia, Amazonocrinis, and Dendronalium from the Biodiversity-rich Northeast Region of India. J. Phycol. 2024, 60, 387–408. [Google Scholar] [CrossRef] [PubMed]
- Carmona Jiménez, J.; Caro Borrero, A.; Becerra-Absalón, I.; Perona Urizar, E.; Márquez Santamaría, K.; Mateo Ortega, P. Description of Two New Species of Nostoc (Nostocales, Cyanobacteria) from Central Mexico, Using Morphological, Ecological, and Molecular Attributes. J. Phycol. 2023, 59, 1237–1257. [Google Scholar] [CrossRef]
- Wang, J.; Wang, S. Preparation, Modification and Environmental Application of Biochar: A Review. J. Clean. Prod. 2019, 227, 1002–1022. [Google Scholar] [CrossRef]
- Fattahi, S.M.; Soroush, A.; Huang, N. Wind Erosion Control Using Inoculation of Aeolian Sand with Cyanobacteria. Land Degrad. Dev. 2020, 31, 2104–2116. [Google Scholar] [CrossRef]
- Xu, B.; Zhang, J.; Huang, N.; Gong, K.; Liu, Y. Characteristics of Turbulent Aeolian Sand Movement Over Straw Checkerboard Barriers and Formation Mechanisms of Their Internal Erosion Form. J. Geophys. Res. Atmos. 2018, 123, 6907–6919. [Google Scholar] [CrossRef]
- Rahmonov, O.; Cabala, J.; Bednarek, R.; Rozek, D.; Florkiewicz, A. Role of Soil Algae on the Initial Stages of Soil Formation in Sandy Polluted Areas. Ecol. Chem. Eng. S 2015, 22, 675–690. [Google Scholar] [CrossRef]
- Keqiang, Z.; Zijia, Z.; Cui, Z.; Ling, X.; Delong, M.; Li, W.; Shaoxian, S.; Sancheze, R.M.T.; Farias, M.E. Rapid Artificial Biocrust Development by Cyanobacterial Inoculation and Clay Amendment. Land Degrad. Dev. 2023, 34, 3728–3743. [Google Scholar] [CrossRef]
- Mugnai, G.; Rossi, F.; Martin Noah Linus Felde, V.J.; Colesie, C.; Büdel, B.; Peth, S.; Kaplan, A.; De Philippis, R. The Potential of the Cyanobacterium Leptolyngbya Ohadii as Inoculum for Stabilizing Bare Sandy Substrates. Soil Biol. Biochem. 2018, 127, 318–328. [Google Scholar] [CrossRef]
- Kumar, V.; Kharwar, S. Cyanoremediation: An Overview. In Phytoremediation; Madhav, S., Gupta, G.P., Yadav, R.K., Mishra, R., Hullebusch, E.V., Eds.; Springer: Cham, Switzerland, 2024; pp. 1–35. ISBN 978-3-031-60760-8. [Google Scholar]
- Acea, M. Cyanobacterial Inoculation of Heated Soils: Effect on Microorganisms of C and N Cycles and on Chemical Composition in Soil Surface. Soil Biol. Biochem. 2003, 35, 513–524. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, F.; Yang, T.; Liu, X.; Jiang, Q.; Shang, H.; Zheng, C. Wind Erosion Control of Bare Surface Soil in Arid Mining Area by Cyanobacterial Inoculation and Biochar Amendment. CATENA 2025, 250, 108765. [Google Scholar] [CrossRef]
- Wu, Y.; Rao, B.; Wu, P.; Liu, Y.; Li, G.; Li, D. Development of Artificially Induced Biological Soil Crusts in Fields and Their Effects on Top Soil. Plant Soil 2013, 370, 115–124. [Google Scholar] [CrossRef]
- Patova, E.N.; Kulyugina, E.E.; Deneva, S.V. Processes of Natural Soil and Vegetation Recovery on a Worked-out Open Pit Coal Mine (Bol’shezemel’skaya Tundra). Russ. J. Ecol. 2016, 47, 228–233. [Google Scholar] [CrossRef]
- Zimonina, N.M. Algae in technogenic landscapes of the Far North. In Vozobnovimye Resursy Vodoemov Bol’shezemel’skoi Tundry [Renewable Resources of Water Bodies in the Bol’shezemel’skaya Tundra]; Komi Science Centre of Russian Academy of Science: Syktyvkar, Russia, 2002; pp. 25–32. [Google Scholar]
- Zheng, Y.; Xu, M.; Zhao, J.; Bei, S.; Hao, L. Effects of Inoculated Microcoleus Vaginatus on the Structure and Function of Biological Soil Crusts of Desert. Biol. Fertil. Soils 2011, 47, 473–480. [Google Scholar] [CrossRef]
- Bowker, M.A. Biological Soil Crust Rehabilitation in Theory and Practice: An Underexploited Opportunity. Restor. Ecol. 2007, 15, 13–23. [Google Scholar] [CrossRef]
- Pandey, V.C.; Singh, N. Impact of Fly Ash Incorporation in Soil Systems. Agric. Ecosyst. Environ. 2010, 136, 16–27. [Google Scholar] [CrossRef]
- Domracheva, L.I.; Dabakh, E.V.; Kondakova, L.V.; Varaksina, A.I. Algal–Mycological Complexes in Soils upon Their Chemical Pollution. Eurasian Soil Sci. 2006, 39, 91–97. [Google Scholar] [CrossRef]
- Davydov, D.A.; Redkina, V.V. Algae and Cyanoprokaryotes on Naturally Overgrowing Ash Dumps of the Apatity Thermal Power Station (Murmansk Region). Proc. Karelian Res. Cent. Russ. Acad. Sci. 2021, 1, 51. [Google Scholar] [CrossRef]
- Pereverzev, V.N. Soil Formation in the Forest Zone of the Kola Peninsula. Bull. Kola Sci. Cent. Russ. Acad. Sci. 2011, 74–82. [Google Scholar]
- Geitler, L. Oscillatoria Carotinosa n. Sp. and O. Pseudoacutissima n. Sp., Zwei Arten Mit Lokalisierter Karotinoidbildung. Österr. Bot. Z. 1956, 103, 342–345. [Google Scholar] [CrossRef]
- Graveland, J. NDFF Rijkswaterstaat; NDFF: Leiden, The Netherlands, 2018. [Google Scholar]
- Iglesias, M.J.; Soengas, R.; Martins, C.B.; Correia, M.J.; Ferreira, J.D.; Santos, L.M.A.; Ortiz, F.L. Chemotaxonomic Profiling Through NMR1. J. Phycol. 2020, 56, 521–539. [Google Scholar] [CrossRef]
- Yim, B.C.; Kim, J.H.; Yoon, S.R.; Lee, N.J.; Lee, O.M. A Study of Seven Unrecorded Species of Benthic and Subaerophytic Cyanobacteria (Cyanophyceae, Cyanophyta) in Korea. Environ. Biol. Res. 2018, 36, 291–298. [Google Scholar] [CrossRef]
- Nikulin, A.Y.; Nikulin, V.Y.; Bagmet, V.B.; Allaguvatova, R.Z.; Abdullin, S.R. New Data on Cyanobacteria and Algae in the Russian Far East. Part II. Biota Environ. Nat. Areas 2022, 10, 5–15. [Google Scholar] [CrossRef]
- Davydov, D.; Vilnet, A.; Novakovskaya, I.; Patova, E. Terrestrial Species of Drouetiella (Cyanobacteria, Oculatellaceae) from the Russian Arctic and Subarctic Regions and Description of Drouetiella ramosa sp. Nov. Diversity 2023, 15, 132. [Google Scholar] [CrossRef]
- Cai, F.; Li, S.; Chen, J.; Li, R. Gansulinema Gen. Nov. and Komarkovaeasiopsis Gen. Nov.: Novel Oculatellacean Genera (Cyanobacteria) Isolated from Desert Soils and Hot Spring. J. Phycol. 2024, 60, 432–446. [Google Scholar] [CrossRef]
- Kaštovský, J.; Johansen, J.R.; Hauerová, R.; Akagha, M.U. Hot Is Rich—An Enormous Diversity of Simple Trichal Cyanobacteria from Yellowstone Hot Springs. Diversity 2023, 15, 975. [Google Scholar] [CrossRef]
- Mai, T.; Johansen, J.R.; Pietrasiak, N.; Bohunická, M.; Martin, M.P. Revision of the Synechococcales (Cyanobacteria) through Recognition of Four Families Including Oculatellaceae Fam. Nov. and Trichocoleaceae Fam. Nov. and Six New Genera Containing 14 Species. Phytotaxa 2018, 365, 1. [Google Scholar] [CrossRef]
- Kim, S.-W.; Lee, N.-J.; Kim, D.-H.; Song, J.-H.; Wang, H.-R.; Lee, O.M. Five Newly Recorded Species of Cyanobacteria in Korea. J. Species Res. 2022, 11, 296–309. [Google Scholar] [CrossRef]
- Osorio-Santos, K.; Pietrasiak, N.; Bohunická, M.; Miscoe, L.H.; Kováčik, L.; Martin, M.P.; Johansen, J.R. Seven New Species of Oculatella (Pseudanabaenales, Cyanobacteria): Taxonomically Recognizing Cryptic Diversification. Eur. J. Phycol. 2014, 49, 450–470. [Google Scholar] [CrossRef]
- Becerra-Absalón, I.; Johansen, J.R.; Osorio-Santos, K.; Montejano, G. Two New Oculatella (Oculatellaceae, Cyanobacteria) Species in Soil Crusts from Tropical Semi-Arid Uplands of México. Fottea 2020, 20, 160–170. [Google Scholar] [CrossRef]
- Chamizo, S.; Adessi, A.; Torzillo, G.; De Philippis, R. Exopolysaccharide Features Influence Growth Success in Biocrust-Forming Cyanobacteria, Moving From Liquid Culture to Sand Microcosms. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Rossi, F.; De Philippis, R. Role of Cyanobacterial Exopolysaccharides in Phototrophic Biofilms and in Complex Microbial Mats. Life 2015, 5, 1218–1238. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.D.; Shukla, P.N.; Giri, D.D.; Kashyap, A.K. Cyanobacteria in Alkaline Soil and the Effect of Cyanobacteria Inoculation with Pyrite Amendments on Their Reclamation. Biol. Fertil. Soils 2005, 41, 451–457. [Google Scholar] [CrossRef]
- Yadav, P.; Gupta, R.K.; Singh, R.P.; Yadav, P.K.; Jalaluddin; Patel, A.K.; Pandey, K.D. Chapter 9—Role of Cyanobacteria in Green Remediation. In Sustainable Environmental Clean-Up; Kumar Mishra, V., Kumar, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 187–210. ISBN 978-0-12-823828-8. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site | Number of Sample Plots | LongitudeE | LatitudeN | Elevation (m a.s.l.) |
|---|---|---|---|---|
| The newest dump | 1 | 33.464710 | 67.599360 | 180 |
| 3-year-old dump | 2 | 33.489820 | 67.598180 | 180 |
| 3 | 33.480890 | 67.599900 | 180 | |
| 4 | 33.481280 | 67.599972 | 202 | |
| 30-year-old dump | 5 | 33.473850 | 67.598560 | 188 |
| 6 | 33.473800 | 67.598520 | 189 | |
| 7 | 33.473342 | 67.598548 | 189 |
| Taxon | Number of a Strain | Number of Sample Plots | Genbank Accession Number (16S–23S rRNA) |
|---|---|---|---|
| Oculatellaceae | |||
| Drouetiella lurida | KPABG-133799 | 3 | OR209487 |
| D. lurida | KPABG-4164 | 4 | OR209489 |
| D. lurida | KPABG-41662 | 5 | ON897680 |
| Gansulinema desertorum | KPABG-133794 | 5 | PV645109 |
| Tildeniella torsiva | KPABG-133811 | 2 | PV645110 |
| T. torsiva | KPABG-4424 | 5 | PV645111 |
| Oculatella leonae | KPABG-44242 | 5 | PV645112 |
| O. coburnii | KPABG-133798 | 3 | PV645113 |
| Nodosilineaceae | |||
| Nodosilinea calida | KPABG-133816 | 1 | PV645114 |
| Wilmottiaceae | |||
| Anagnostidinema pseudacutissimum | KPABG-4411 | 1 | PV645115 |
| A. pseudacutissimum | KPABG-133809 | 1 | PV645116 |
| A. pseudacutissimum | KPABG-133632 | 1 | PV645117 |
| A. pseudacutissimum | KPABG-133633 | 1 | PV645118 |
| A. pseudacutissimum | KPABG-4431 | 5 | PV645119 |
| Microcoleaceae | |||
| Microcoleus sp. | KPABG-133810 | 1 | PV645120 |
| Nostocaceae | |||
| Nostoc commune | KPABG-236 | 7 | PV645121 |
| N. commune | KPABG-237 | 7 | PV645122 |
| N. commune | KPABG-238 | 7 | PV645123 |
| N. commune | KPABG-239 | 7 | PV645124 |
| N. commune | KPABG-240 | 7 | PV645125 |
| N. commune | KPABG-241 | 7 | PV645126 |
| Nostocalean cyanobacterium 1 | KPABG-4166 | 5 | PV645127 |
| Nodulariaceae | |||
| Nostocalean cyanobacterium 2 | KPABG-133801 | 3 | PV645128 |
| Nostocalean cyanobacterium 2 | KPABG-133804 | 2 | PV645129 |
| Species | Number of Sample Plots | |||||
|---|---|---|---|---|---|---|
| The Newest Dump | 3-Year-Old Dump | 30-Year-Old Dump | ||||
| Field Sample | Culture | Field Sample | Culture | Field Sample | Culture | |
| Anagnostidinema pseudacutissimum (Geitler) Strunecký et al. | 1 | 5 | 5 | |||
| Aphanocapsa sp. | 4 | |||||
| Aphanothece pallida (Kütz.) Rabenh. | 5, 6 | |||||
| A. saxicola Näg. | 7 | |||||
| Calothrix sp. | 7 | |||||
| Drouetiella lurida (Gom.) Mai et al. | 3, 4 | 3, 4 | 5 | 5 | ||
| Gansulinema desertorum S. Li et F. Cai | 5 | |||||
| Microchaete tenera Thur. ex Born. et Flah. | 6 | |||||
| Microcoleus sp. (cf. autumnalis) | 1 | 1 | 5 | |||
| Nodosilinea calida J.R. Johansen et al. | 1 | 1 | ||||
| Nostoc cf. pruniforme C. Ag. ex Born. et Flah. | 1 | |||||
| Nostoc commune Vauch. ex Born. et Flah. | 4 | 7 | 7 | |||
| Nostocalean cyanobacterium 1 | 5 | 5 | ||||
| Nostocalean cyanobacterium 2 | 2, 3 | 2, 3 | ||||
| Oculatella leonae Becerra-Absalón et J.R. Johansen | 5 | |||||
| Oculatella coburnii N. Pietrasiak et J.R. Johansen | 3 | |||||
| Synechococcus elongatus (Näg.) Näg. | 4 | |||||
| Tildeniella torsiva Mai et al. | 2 | 2 | 5 | 5 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davydov, D.; Vilnet, A. Cyanobacterial Assemblages Inhabiting the Apatity Thermal Power Plant Fly Ash Dumps in the Russian Arctic. Microorganisms 2025, 13, 1762. https://doi.org/10.3390/microorganisms13081762
Davydov D, Vilnet A. Cyanobacterial Assemblages Inhabiting the Apatity Thermal Power Plant Fly Ash Dumps in the Russian Arctic. Microorganisms. 2025; 13(8):1762. https://doi.org/10.3390/microorganisms13081762
Chicago/Turabian StyleDavydov, Denis, and Anna Vilnet. 2025. "Cyanobacterial Assemblages Inhabiting the Apatity Thermal Power Plant Fly Ash Dumps in the Russian Arctic" Microorganisms 13, no. 8: 1762. https://doi.org/10.3390/microorganisms13081762
APA StyleDavydov, D., & Vilnet, A. (2025). Cyanobacterial Assemblages Inhabiting the Apatity Thermal Power Plant Fly Ash Dumps in the Russian Arctic. Microorganisms, 13(8), 1762. https://doi.org/10.3390/microorganisms13081762