
Analysis of Bacterial Community During Cow Manure and Wheat Straw Composting and the Isolation of Lignin-Degrading Bacteria from the Compost

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Composting Process and Sampling
2.2. Compost Physicochemical Properties Analysis
2.3. DNA Extraction and High-Throughput Sequencing
2.4. Bioinformatics and Data Analysis
2.5. Isolation of Lignin-Degrading Bacteria from Compost
2.6. 16S rRNA Sequencing Determination and Physiological Characteristics
2.7. Determination of the Biodegradation of Alkali Lignin
2.8. Lignolytic Enzyme Activity
2.9. Statistical Analysis
3. Results
3.1. Changes in Physicochemical Parameters During Composting
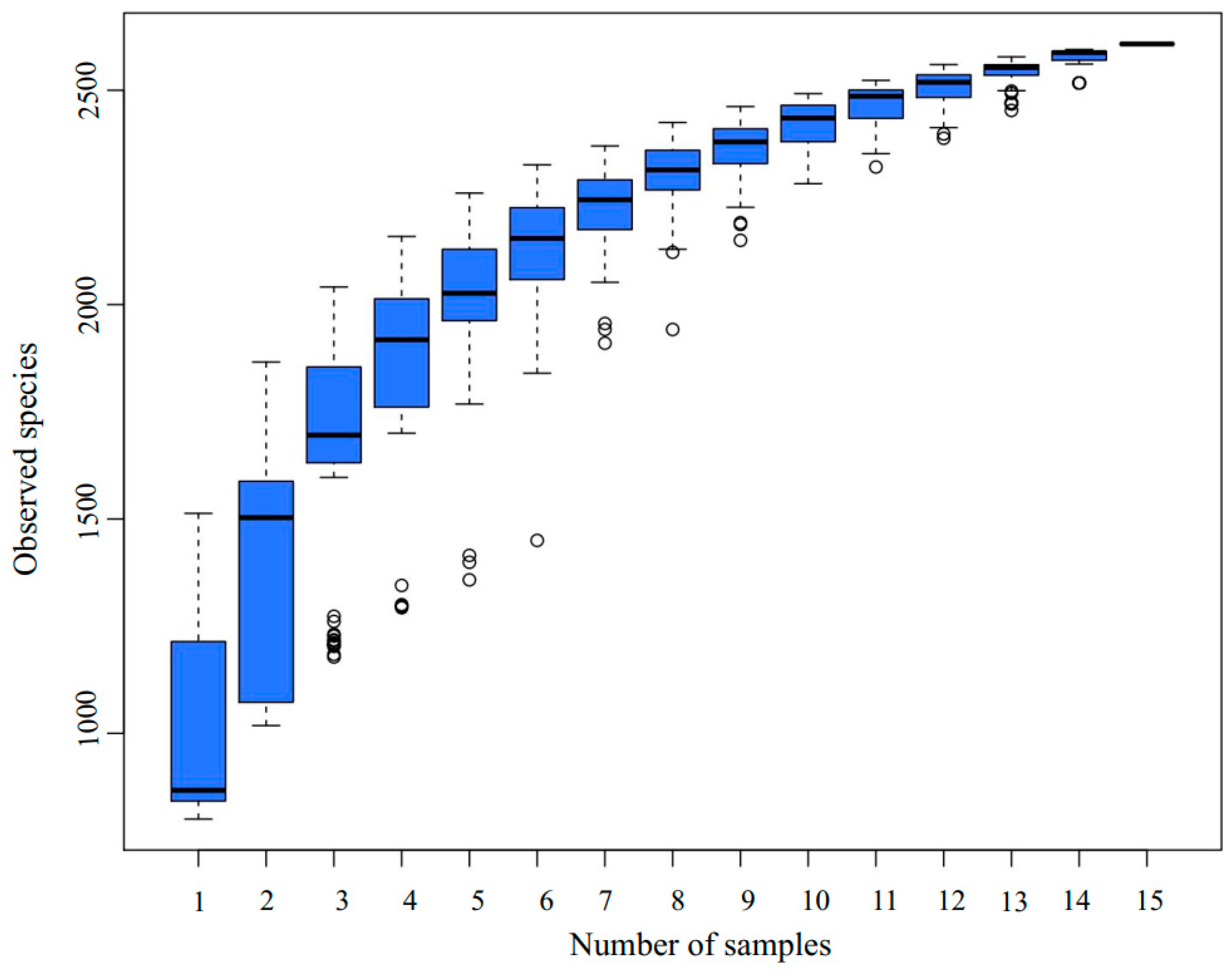
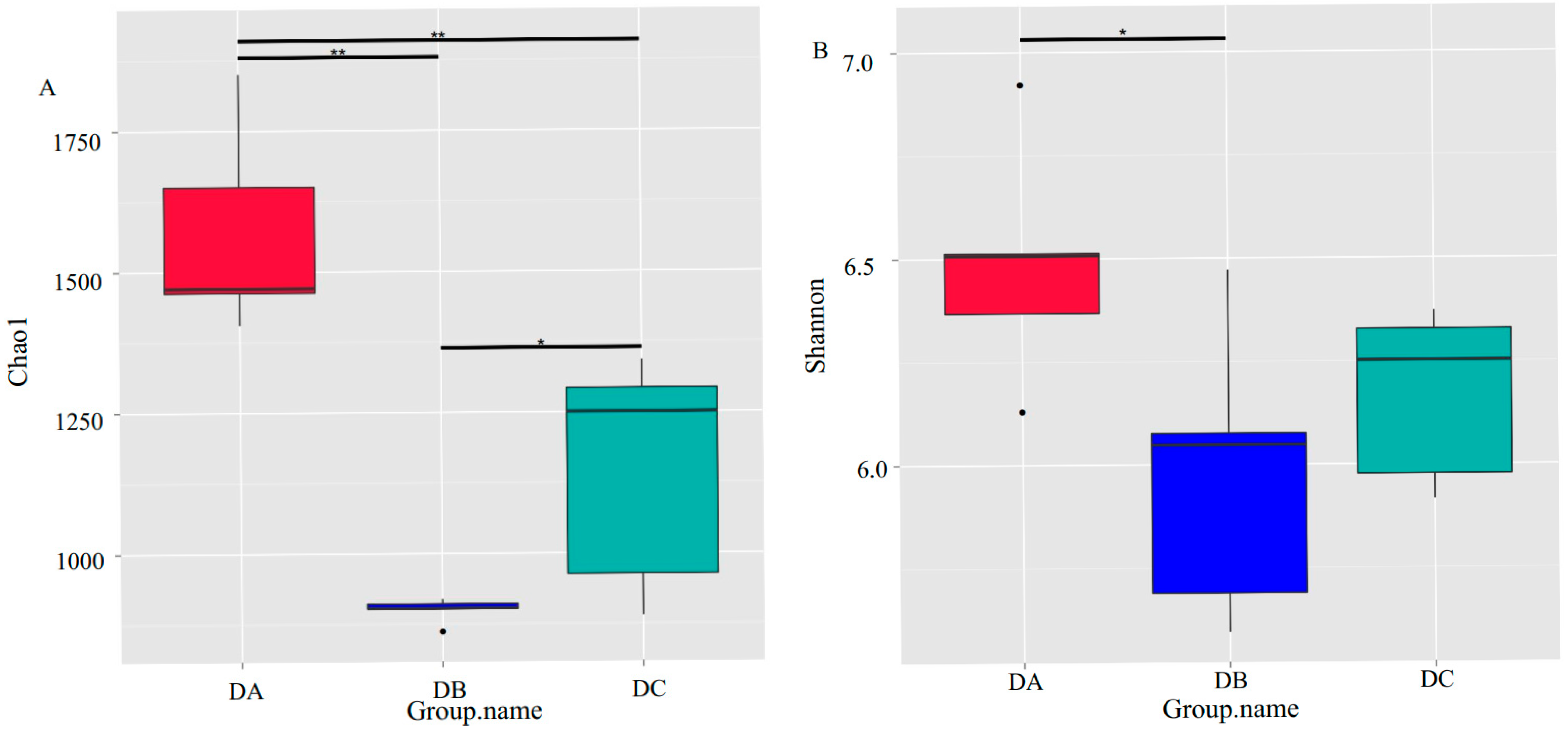
3.2. 16S Amplicon High-Throughput Sequencing
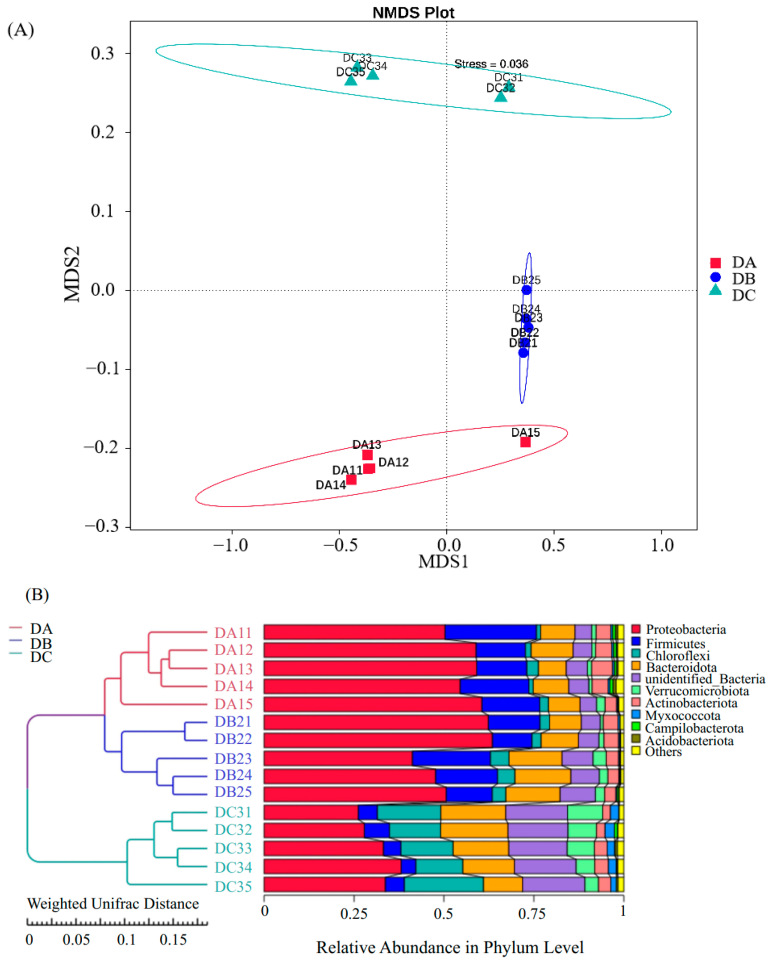
3.3. Evolution of Bacterial Community During Composting
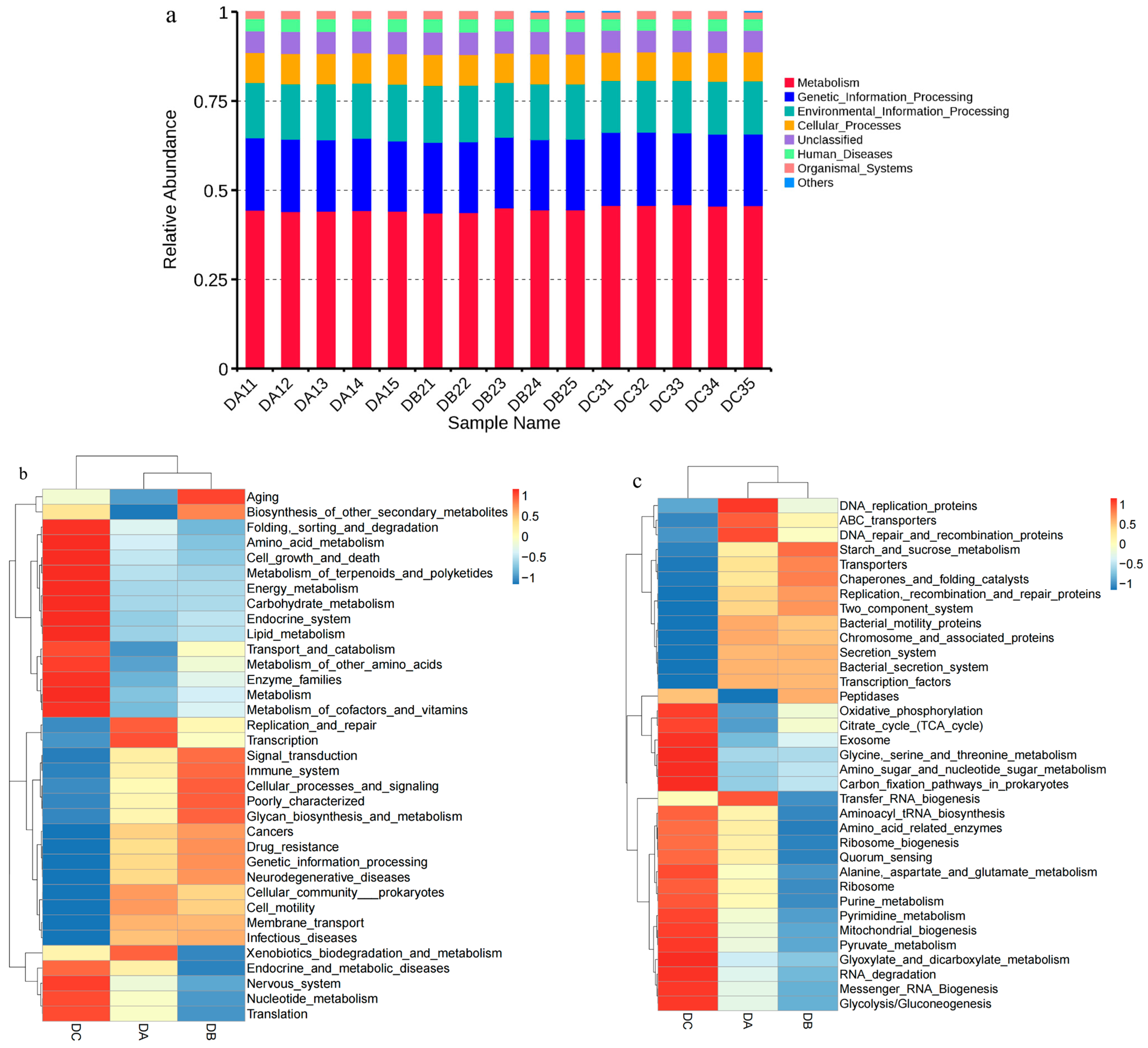
3.4. Function Prediction
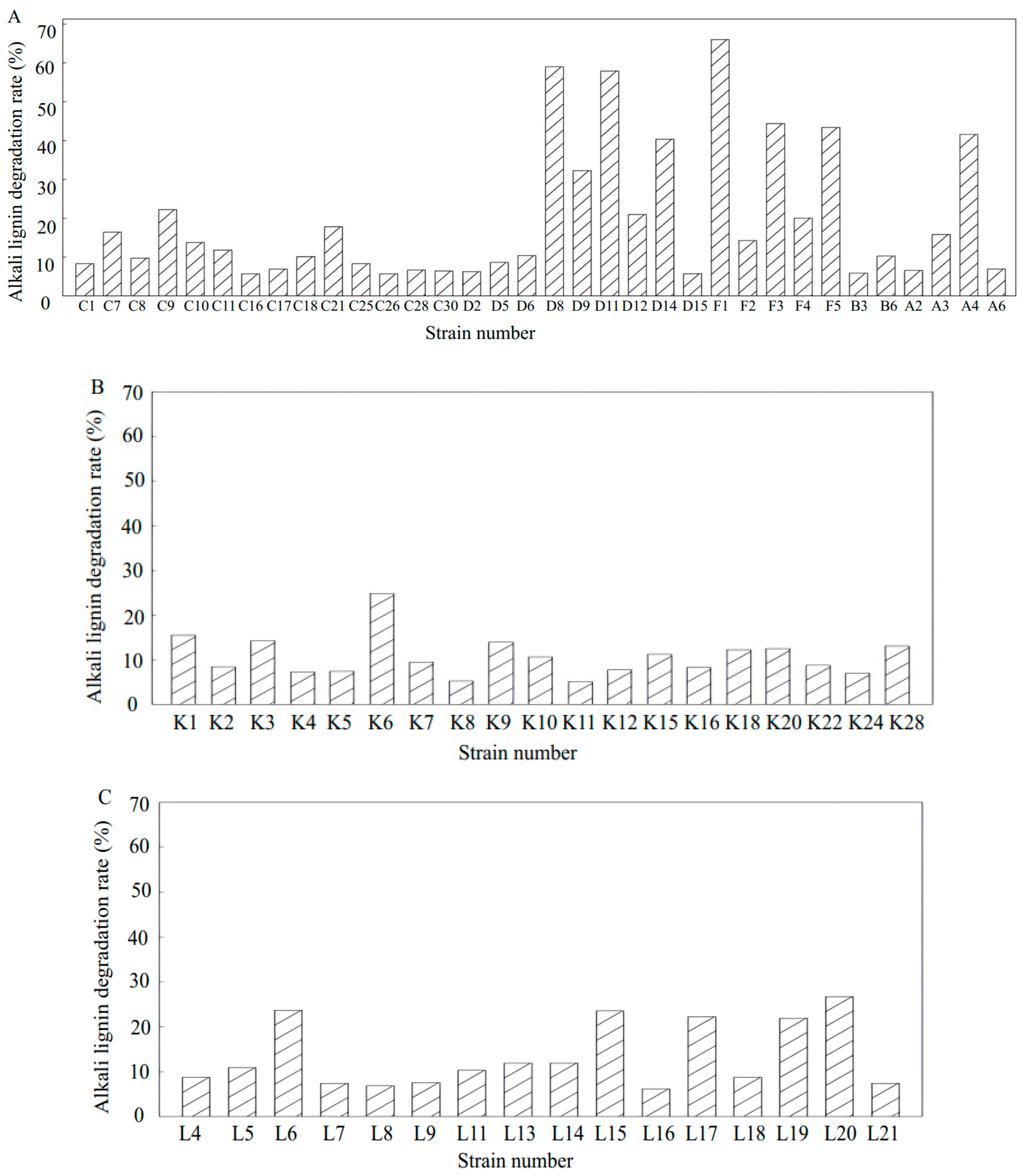
3.5. Screening of High-Efficiency Lignin-Degrading Strains
3.6. Degradation of Alkali Lignin by Lignin-Degrading Strains
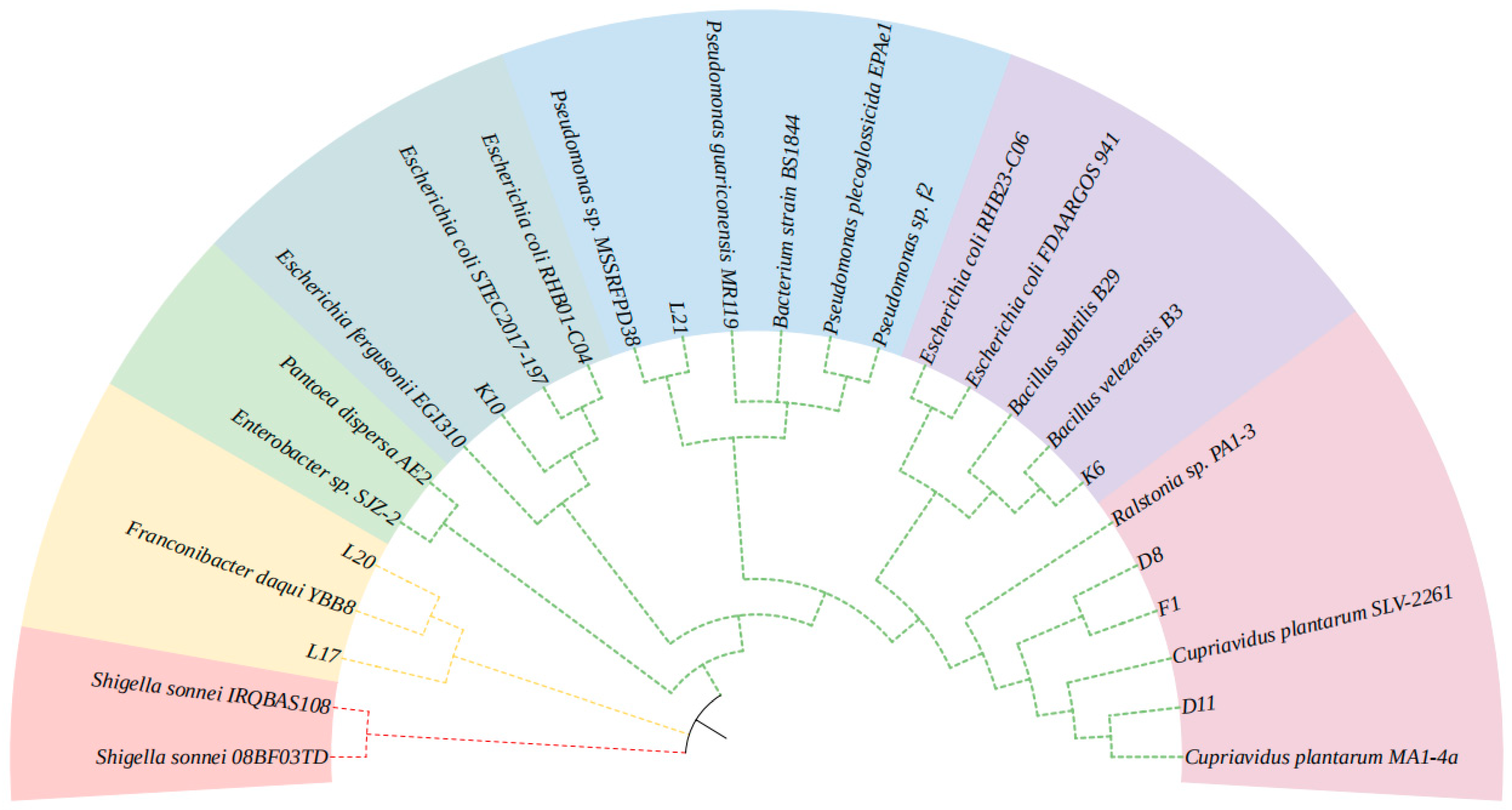
3.7. Identification of Isolated Lignin-Degrading Strains
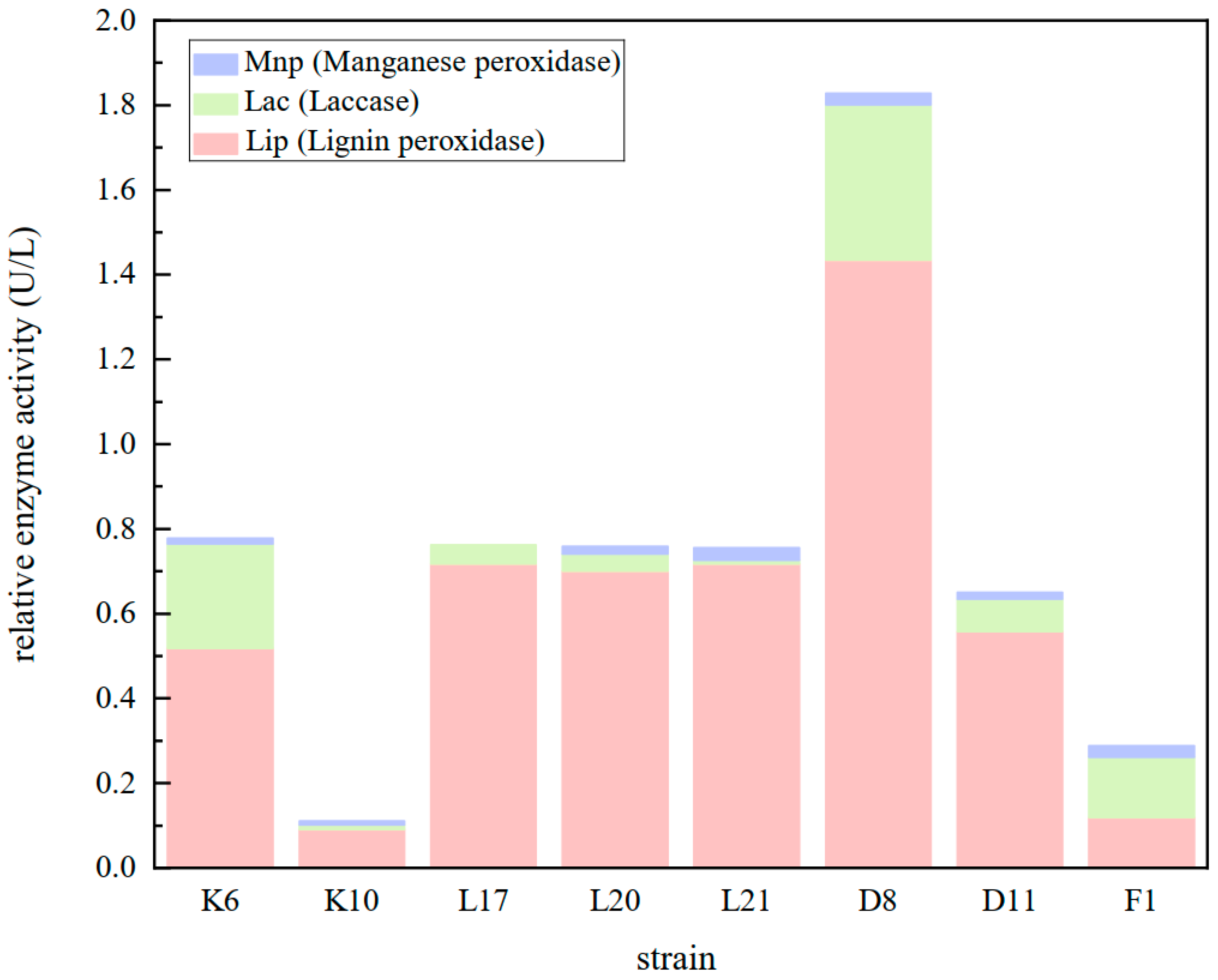
3.8. Enzyme Activity of Isolated Lignin-Degrading Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brodin, M.; Vallejos, M.; Opedal, M.T.; Area, M.C.; Chinga-Carrasco, G. Lignocellulosics as sustainable resources for production of bioplastics—A review. J. Clean. Prod. 2017, 162, 646–664. [Google Scholar] [CrossRef]
- Yadav, M.; Paritosh, K.; Pareek, N.; Vivekanand, V. Coupled treatment of lignocellulosic agricultural residues for augmented biomethanation. J. Clean. Prod. 2019, 213, 75–88. [Google Scholar] [CrossRef]
- Yang, W.; Li, X.; Zhang, Y. Research Progress and the development trend of the utilization of crop straw biomass resources in China. Front. Chem. 2022, 10, 904660. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Sun, R. Recent advances in lignocellulose prior-fractionation for biomaterials, biochemicals, and bioenergy. Carbohydr. Polym. 2021, 261, 117884. [Google Scholar] [CrossRef] [PubMed]
- Hu, N.; Liu, X.; Wei, S.; Yao, J.; Wang, W.; Liu, B.; Tang, T.; Jiang, J.; Wang, L. Current status and future prospects of pretreatment for tobacco stalk lignocellulose. Front. Bioeng. Biotechnol. 2024, 12, 1465419. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, C.; Wu, X.; Tsang, C.W.; Mou, J.; Yan, J.; Liu, Y.; Lin, C.S.K. Recent advancement in lignin biorefinery: With special focus on enzymatic degradation and valorization. Bioresour. Technol. 2019, 291, 121898. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, C.; Sun, X.; Su, S.; Li, Q.; Linhardt, R.J. Efficient, environmentally-friendly and specific valorization of lignin: Promising role of non-radical lignolytic enzymes. World J. Microbiol. Biotechnol. 2017, 33, 125. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Han, D.; Yang, S. Combinations of mild chemical and bacterial pretreatment for improving enzymatic saccharification of corn stover. Biotechnol. Biotechnol. Equip. 2022, 36, 598–608. [Google Scholar] [CrossRef]
- Zhao, X.; Zheng, Z.; Cai, Y.; Zhao, Y.; Zhang, Y.; Gao, Y.; Cui, Z.; Wang, X. Accelerated biomethane production from lignocellulosic biomass: Pretreated by mixed enzymes secreted by Trichoderma viride and Aspergillus sp. Bioresour. Technol. 2020, 309, 123378. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Li, Y. Fungal pretreatment of lignocellulosic biomass. Biotechnol. Adv. 2012, 30, 1447–1457. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Ko, Y.; Hyeon, J.; Han, S. Studies of advanced lignin valorization based on various types of lignolytic enzymes and microbes. Bioresour. Technol. 2019, 289, 121728. [Google Scholar] [CrossRef] [PubMed]
- Kainthola, J.; Podder, A.; Fechner, M.; Goel, R. An overview of fungal pretreatment processes for anaerobic digestion: Applications, bottlenecks and future needs. Bioresour. Technol. 2021, 321, 124397. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Liu, X.; Zhang, J.; Sui, W.; Jang, J.; Si, C. One-pot lignin depolymerization and activation by solid acid catalytic phenolation for lightweight phenolic foam preparation. Ind. Crops Prod. 2018, 124, 216–225. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, L.; Liu, F.; Fan, X.; Sun, R.C. One-step process of hydrothermal and alkaline treatment of wheat straw for improving the enzymatic saccharification. Biotechnol. Biofuels 2018, 11, 137. [Google Scholar] [CrossRef] [PubMed]
- Hao, N.; Lu, K.; Ben, H.; Adhikari, S.; Lacerda, T.B.; Ragauskas, A.J. Effect of autohydrolysis pretreatment conditions on sugarcane bagasse structures and product distribution resulting from pyrolysis. Energy Technol. 2018, 6, 640–648. [Google Scholar] [CrossRef]
- Mazarji, M.; Alvarado-Morales, M.; Tsapekos, P.; Nabi-Bidhendi, G.; Mahmoodi, N.M.; Angelidaki, I. Graphene based ZnO nanoparticles to depolymerize lignin-rich residues via UV/iodide process. Environ. Int. 2019, 125, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Wang, X.; Deng, Y.; Tian, Z.; Huang, C.; Fang, G. Comprehensive insights of pretreatment strategies on the structures and bioactivities variation of lignin-carbohydrate complexes. Front. Bioeng. Biotechnol. 2024, 12, 1465328. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Wei, L.; Pan, X.; Liu, C.; Jiang, J.; Wang, K. The pretreatment of lignocelluloses with green solvent as biorefinery preprocess: A minor review. Front. Plant Sci. 2021, 12, 670061. [Google Scholar] [CrossRef] [PubMed]
- Picart, P.; Wiermans, L.; Pérez-Sánchez, M.; Grande, P.M.; Schallmey, A.; Domínguez de María, P. Assessing lignin types to screen novel biomass-degrading microbial strains: Synthetic lignin as useful carbon source. ACS Sustain. Chem. Eng. 2016, 4, 651–655. [Google Scholar] [CrossRef]
- Wang, H.; Peng, X.; Li, H.; Giannis, A.; He, C. Recent biotechnology advances in bio-conversion of lignin to lipids by bacterial cultures. Front. Chem. 2022, 10, 894593. [Google Scholar] [CrossRef] [PubMed]
- Singhania, R.R.; Patel, A.K.; Raj, T.; Chen, C.W.; Ponnusamy, V.K.; Tahir, N.; Kim, S.H.; Dong, C.D. Lignin valorisation via enzymes: A sustainable approach. Fuel 2022, 311, 122608. [Google Scholar] [CrossRef]
- Sanchez, C. Lignocellulosic residues: Biodegradation and bioconversion by fungi. Biotechnol. Adv. 2009, 27, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, D.; Liu, Y. Study on comprehensive energy-saving of shading and photovoltaics of roof added pv module. Energy Procedia 2017, 132, 598–603. [Google Scholar] [CrossRef]
- De Gonzalo, G.; Colpa, D.I.; Habib, M.H.; Fraaije, M.W. Bacterial enzymes involved in lignin degradation. J. Biotechnol. 2016, 236, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kang, M.; Bae, J.H.; Sohn, J.H.; Sung, B.H. Bacterial valorization of lignin: Strains, enzymes, conversion pathways, biosensors, and perspectives. Front. Bioeng. Biotechnol. 2019, 7, 209. [Google Scholar] [CrossRef] [PubMed]
- Aston, J.E.; Apel, W.A.; Lee, B.D.; Thompson, D.N.; Lacey, J.A.; Newby, D.T.; Reed, D.W.; Thompson, V.S. Degradation of phenolic compounds by the lignocellulose deconstructing thermoacidophilic bacterium Alicyclobacillus acidocaldarius. J. Ind. Microbiol. Biotechnol. 2016, 43, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Dar, M.A.; Shaikh, A.A.; Pawar, K.D.; Pandit, R.S. Exploring the gut of Helicoverpa armigera for cellulose degrading bacteria and evaluation of a potential strain for lignocellulosic biomass deconstruction. Process Biochem. 2018, 73, 142–153. [Google Scholar] [CrossRef]
- Li, X.; Li, M.; Pu, Y.; Ragauskas, A.J.; Zheng, Y. Simultaneous depolymerization and fermentation of lignin into value-added products by the marine protist, Thraustochytrium striatum. Algal Res. 2020, 46, 101773. [Google Scholar] [CrossRef]
- Li, X.; He, Y.; Zhang, L.; Xu, Z.; Ben, H.; Gaffrey, M.J.; Yang, Y.; Yang, S.; Yuan, J.S.; Qian, W.J.; et al. Discovery of potential pathways for biological conversion of poplar wood into lipids by co-fermentation of Rhodococci strains. Biotechnol. Biofuels 2019, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- Rashid, G.M.; Taylor, C.R.; Liu, Y.; Zhang, X.; Rea, D.; Fulop, V.; Bugg, T.D. Identification of manganese superoxide dismutase from Sphingobacterium sp. T2 as a novel bacterial enzyme for lignin oxidation. ACS Chem. Biol. 2015, 10, 2286–2294. [Google Scholar] [CrossRef] [PubMed]
- Chong, G.; Huang, X.; Di, J.; Xu, D.; He, Y.; Pei, Y.; Tang, Y.; Ma, C. Biodegradation of alkali lignin by a newly isolated Rhodococcus pyridinivorans CCZU-B16. Bioprocess Biosyst. Eng. 2018, 41, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Wilkinson, R.C.; Rashid, G.M.M.; Brown, D.; Fulop, V.; Bugg, T.D.H. Characterization of thiamine diphosphate-dependent 4-hydroxybenzoylformate decarboxylase enzymes from Rhodococcus jostii RHA1 and pseudomonas fluorescens Pf-5 Involved in degradation of aryl c2 lignin degradation fragments. Biochemistry 2019, 58, 5281–5293. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhao, J.; Jiang, J.; Xu, H.; Zhang, N.; Xie, J.; Wei, M. Isolation and characterization of Bacillus sp. capable of degradating alkali lignin. Front. Energy Res. 2021, 9, 807286. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhao, Y.; Ni, K.; Shi, Y.; Xu, Q. Characterization of ligninolytic bacteria and analysis of alkali-lignin biodegradation products. Pol. J. Microbiol. 2020, 69, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Shen, X.; Gang, L.; Xu, H.; Wu, F.; Sheng, L. A novel lignin degradation bacteria-Bacillus amyloliquefaciens SL-7 used to degrade straw lignin efficiently. Bioresour. Technol. 2020, 310, 123445. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Verma, S.; Gazara, R.K.; Kumar, M.; Pandey, A.; Verma, P.K.; Thakur, I.S. Genomic and proteomic analysis of lignin degrading and polyhydroxyalkanoate accumulating beta-proteobacterium Pandoraea sp. ISTKB. Biotechnol. Biofuels 2018, 11, 154. [Google Scholar] [CrossRef] [PubMed]
- Riyadi, F.A.; Tahir, A.A.; Yusof, N.; Sabri, N.S.A.; Noor, M.J.M.M.; Akhir, F.N.M.D.; Othman, N.A.; Zakaria, Z.; Hara, H. Enzymatic and genetic characterization of lignin depolymerization by Streptomyces sp. S6 isolated from a tropical environment. Sci. Rep. 2020, 10, 7813. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Taylor, C.R.; Pink, D.; Burton, K.; Eastwood, D.; Bending, G.D.; Bugg, T.D. Development of novel assays for lignin degradation: Comparative analysis of bacterial and fungal lignin degraders. Mol. Biosyst. 2010, 6, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Bugg, T.D.; Ahmad, M.; Hardiman, E.M.; Singh, R. The emerging role for bacteria in lignin degradation and bio-product formation. Curr. Opin. Biotechnol. 2011, 22, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Ogawa, Y. In vitro protein digestibility and biochemical characteristics of soaked, boiled and fermented soybeans. Sci. Rep. 2021, 11, 14257. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Wang, X.; Yang, T.; Wei, Z.; Banerjee, S.; Friman, V.P.; Mei, X.; Xu, Y.; Shen, Q. Livestock manure type affects microbial community composition and assembly during composting. Front. Microbiol. 2021, 12, 621126. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Chen, Q.; Qin, Y.; Yang, Y.; Yang, Q.; Wang, Y.; Cheng, Z.; Cao, N.; Zhang, G. Succession of the microbial communities and function prediction during short-term peach sawdust-based composting. Bioresour. Technol. 2021, 332, 125079. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Liu, X.Y.; Liu, M.Y.; Liu, W.X.; Xu, J.Z.; Li, Y.W. Comparative evaluation of 16S rRNA primer pairs in identifying nitrifying guilds in soils under long-term organic fertilization and water management. Front. Microbiol. 2024, 15, 1424795. [Google Scholar] [CrossRef] [PubMed]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.H.; Xu, X.; Xu, F.J.; Zhang, Y.; Su, P.; Shen, C.F. Developing the script “degenerate primer 111” to enhance the coverage of universal primers for the small subunit rRNA gene on target microorganisms. Front. Microbiol. 2024, 15, 1394303. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Wang, T.; Gao, L.; Yin, H.; Lu, X. Isolation, identification and characterization of lignin-degrading bacteria from Qinling, China. J. Appl. Microbiol. 2017, 123, 1447–1460. [Google Scholar] [CrossRef] [PubMed]
- Camarero, S.; Ibarra, D.; Martínez, M.J.; Martínez, Á.T. Lignin-derived compounds as efficient laccase mediators for decolorization of different types of recalcitrant dyes. Appl. Environ. Microbiol. 2005, 71, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xiao, J.; Wang, G.; Chen, G. Enzymatic hydrolysis of lignin by ligninolytic enzymes and analysis of the hydrolyzed lignin products. Bioresour. Technol. 2020, 304, 122975. [Google Scholar] [CrossRef] [PubMed]
- Morales, G.M.; Ali, S.S.; Si, H.; Zhang, W.; Zhang, R.; Hosseini, K.; Sun, J.; Zhu, D. Acidic versus alkaline bacterial degradation of lignin through engineered strain E. coli BL21 (Lacc): Exploring the differences in chemical structure, morphology, and degradation products. Front. Bioeng. Biotechnol. 2020, 8, 671. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Zhang, P.; Xie, C.; Zhang, W.; Sun, J.; Qian, W.; Yang, B. Biodegradation of alkaline lignin by Bacillus ligniniphilus L1. Biotechnol. Biofuels 2017, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Sukhikh, S.; Kalashnikova, O.; Ivanova, S.; Prosekov, A.; Krol, O.; Kriger, O.; Fedovskikh, N.; Babich, O. Evaluating the influence of microbial fermentation on the nutritional value of soybean meal. Fermentation 2022, 8, 458. [Google Scholar] [CrossRef]
- Tahmasian, A.; Drew, R.; Broadbent, J.A.; Juhász, A.; Nye-Wood, M.; Colgrave, M.L. Conventional solid-state fermentation impacts the white lupin proteome reducing the abundance of allergenic peptides. Food Chem. 2023, 426, 136622. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Z.; Xia, J.; Chen, Y. Effect of microbial inoculation on physicochemical properties and bacterial community structure of citrus peel composting. Bioresour. Technol. 2019, 291, 121843. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Yang, W.; Men, M.; Bello, A.; Xu, X.; Xu, B.; Deng, L.; Jiang, X.; Sheng, S.; Wu, X.; et al. Microbial community succession and response to environmental variables during cow manure and corn straw composting. Front. Microbiol. 2019, 10, 529. [Google Scholar] [CrossRef] [PubMed]
- Sundberg, C.; Smars, S.; Jonsson, H. Low pH as an inhibiting factor in the transition from mesophilic to thermophilic phase in composting. Bioresour. Technol. 2004, 95, 145. [Google Scholar] [CrossRef] [PubMed]
- Huhe; Jiang, C.; Wu, Y.P.; Cheng, Y.X. Bacterial and fungal communities and contribution of physicochemical factors during cattle farm waste composting. Microbiologyopen 2017, 6, e518. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.F.; Wan, J.X.; Jiang, G.F.; Yang, T.J.; Banerjee, S.; Wei, Z.; Mei, X.L.; Friman, V.P.; Xu, Y.C.; Shen, Q.R. Compositional and functional succession of bacterial and fungal communities is associated with changes in abiotic properties during pig manure composting. Waste Manag. 2021, 131, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Pan, T.; Fu, D.F.; Fotidis, I.A.; Moulogianni, C.; Yan, Y.X.; Singh, R.P. Pilot-scale membrane-covered composting of food waste: Initial moisture, mature compost addition, aeration time and rate. Sci. Total Environ. 2024, 926, 171797. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chu, C.; Li, X.K.; Wang, W.; Ren, N.Q. Succession of bacterial community function in cow manure composing. Bioresour. Technol. 2018, 267, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhao, J.; Feng, W.; Zhang, Z.; Gu, Y.; Wang, Y. Microbial community succession of cow manure and tobacco straw composting. Front. Microbiomes 2024, 3, 1301156. [Google Scholar] [CrossRef]
- Jain, M.S.; Daga, M.; Kalamdhad, A.S. Physical parameters evaluation during production of soil conditioner from aquatic waste: Hydrilla verticillata (L.f.) Royle. Environ. Technol. Innov. 2018, 11, 64–73. [Google Scholar] [CrossRef]
- Zhong, X.-Z.; Li, X.-X.; Zeng, Y.; Wang, S.-P.; Sun, Z.-Y.; Tang, Y.-Q. Dynamic change of bacterial community during dairy manure composting process revealed by high-throughput sequencing and advanced bioinformatics tools. Bioresour. Technol. 2020, 306, 123091. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, J.; Che, Z.; Xue, L. Succession of the Bacterial communities and functional characteristics in sheep manure composting. Biology 2022, 11, 1181. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; He, X.; Tang, J.; Li, X.; Zhao, R.; Tao, Y.; Wang, C.; Qiu, Z. Influence of moisture content on chicken manure stabilization during microbial agent-enhanced composting. Chemosphere 2021, 264, 128549. [Google Scholar] [CrossRef] [PubMed]
- Qiao, C.; Ryan Penton, C.; Liu, C.; Shen, Z.; Ou, Y.; Liu, Z.; Xu, X.; Li, R.; Shen, Q. Key extracellular enzymes triggered high-efficiency composting associated with bacterial community succession. Bioresour. Technol. 2019, 288, 121576. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Wang, Y.Y.; Li, L.; Ma, L.T.; Deng, Y.Q.; Xu, Z. Adjusting pH of the secondary composting materials to further enhance the lignocellulose degradation and promote the humification process. Sustainability 2023, 15, 9032. [Google Scholar] [CrossRef]
- Ge, M.; Shen, Y.; Ding, J.; Meng, H.; Zhou, H.; Zhou, J.; Cheng, H.; Zhang, X.; Wang, J.; Wang, H.; et al. New insight into the impact of moisture content and pH on dissolved organic matter and microbial dynamics during cattle manure composting. Bioresour. Technol. 2022, 344, 126236. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ren, L.H.; Luo, L.; Zhang, J.C.; Zhang, L.H.; Huang, H.L. Bacterial and fungal community dynamics and shaping factors during agricultural waste composting with zeolite and biochar addition. Sustainability 2020, 12, 7082. [Google Scholar] [CrossRef]
- Kausar, H.; Ismail, M.R.; Saud, H.M.; Othman, R.; Habib, S. Use of lignocellulolytic microbial consortium and pH amendment on composting efficacy of rice straw. Compos. Sci. Util. 2013, 21, 121–133. [Google Scholar] [CrossRef]
- Cai, S.Y.; Ma, Y.; Bao, Z.K.; Yang, Z.Y.; Niu, X.Y.; Meng, Q.Z.; Qin, D.S.; Wang, Y.; Wan, J.F.; Guo, X.Y. The impacts of the C/N ratio on hydrogen sulfide emission and microbial community characteristics during chicken manure composting with wheat straw. Agriculture 2024, 14, 948. [Google Scholar] [CrossRef]
- Guo, R.; Li, G.X.; Jiang, T.; Schuchardt, F.; Chen, T.B.; Zhao, Y.Q.; Shen, Y.J. Effect of aeration rate, C/N ratio and moisture content on the stability and maturity of compost. Bioresour. Technol. 2012, 112, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.F.; Wong, J.W.C.; Wu, Q.T.; Nagar, B.B. Effect of C/N on composting of pig manure with sawdust. Waste Manag. 2004, 24, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.C.; Liang, F.Y.; Yu, A.; Li, B.; Yang, L.J. Evaluation of stability and maturity during forced-aeration composting of chicken manure and sawdust at different C/N ratios. Chemosphere 2010, 78, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Qiao, C.C.; Penton, C.R.; Liu, C.; Tao, C.Y.; Deng, X.H.; Ou, Y.N.; Liu, H.J.; Li, R. Patterns of fungal community succession triggered by C/N ratios during composting. J. Hazard. Mater. 2021, 401, 123344. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhang, Y.; Qu, M.; Yin, Y.; Fan, K.; Hu, B.; Zhang, H.; Wei, M.; Ma, C. Effects of biochar on the microbial activity and community structure during sewage sludge composting. Bioresour. Technol. 2019, 272, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, L.; Hassan, M.; Xie, B. Succession of the functional microbial communities and the metabolic functions in maize straw composting process. Bioresour. Technol. 2018, 256, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.Z.; Ma, S.C.; Wang, S.P.; Wang, T.T.; Sun, Z.Y.; Tang, Y.Q.; Deng, Y.; Kida, K. A comparative study of composting the solid fraction of dairy manure with or without bulking material: Performance and microbial community dynamics. Bioresour. Technol. 2018, 247, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.P.; Martins, L.F.; Pereira, R.V.; Thomas, A.M.; Barbosa, D.; Lemos, L.N.; Silva, G.M.; Moura, L.M.; Epamino, G.W.; Digiampietri, L.A.; et al. Microbial community structure and dynamics in thermophilic composting viewed through metagenomics and metatranscriptomics. Sci. Rep. 2016, 6, 38915. [Google Scholar] [CrossRef] [PubMed]
- Toledo, M.; Gutiérrez, M.C.; Siles, J.A.; García-Olmo, J.; Martín, M.A. Chemometric analysis and NIR spectroscopy to evaluate odorous impact during the composting of different raw materials. J. Clean. Prod. 2017, 167, 154–162. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Hu, J.; Zhang, B.; Li, Y.; Yang, C.; Chen, F.; Zhu, T.; Xin, Y. Analysis of Bacterial Community During Cow Manure and Wheat Straw Composting and the Isolation of Lignin-Degrading Bacteria from the Compost. Microorganisms 2025, 13, 1716. https://doi.org/10.3390/microorganisms13081716
Yang H, Hu J, Zhang B, Li Y, Yang C, Chen F, Zhu T, Xin Y. Analysis of Bacterial Community During Cow Manure and Wheat Straw Composting and the Isolation of Lignin-Degrading Bacteria from the Compost. Microorganisms. 2025; 13(8):1716. https://doi.org/10.3390/microorganisms13081716
Chicago/Turabian StyleYang, Hanxiang, Jianguo Hu, Bingxin Zhang, Yan Li, Chenxian Yang, Fusheng Chen, Tingwei Zhu, and Ying Xin. 2025. "Analysis of Bacterial Community During Cow Manure and Wheat Straw Composting and the Isolation of Lignin-Degrading Bacteria from the Compost" Microorganisms 13, no. 8: 1716. https://doi.org/10.3390/microorganisms13081716
APA StyleYang, H., Hu, J., Zhang, B., Li, Y., Yang, C., Chen, F., Zhu, T., & Xin, Y. (2025). Analysis of Bacterial Community During Cow Manure and Wheat Straw Composting and the Isolation of Lignin-Degrading Bacteria from the Compost. Microorganisms, 13(8), 1716. https://doi.org/10.3390/microorganisms13081716