

Lactobacillus murinus Reduces Susceptibility to Secondary MRSA Infection in IAV-Infected Mice Through Promoting a T Cell-Independent IgA Response

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus and Bacteria
2.2. Animal Experiments
2.3. Viral and Bacterial Load Detection
2.4. Hematoxylin and Eosin Staining
2.5. Full Length 16S rRNA Gene Sequencing and Bioinformatics Pipeline
2.6. Flow Cytometry and Enzyme-Linked Immunosorbent Assays
2.7. Statistical Analyses
3. Results
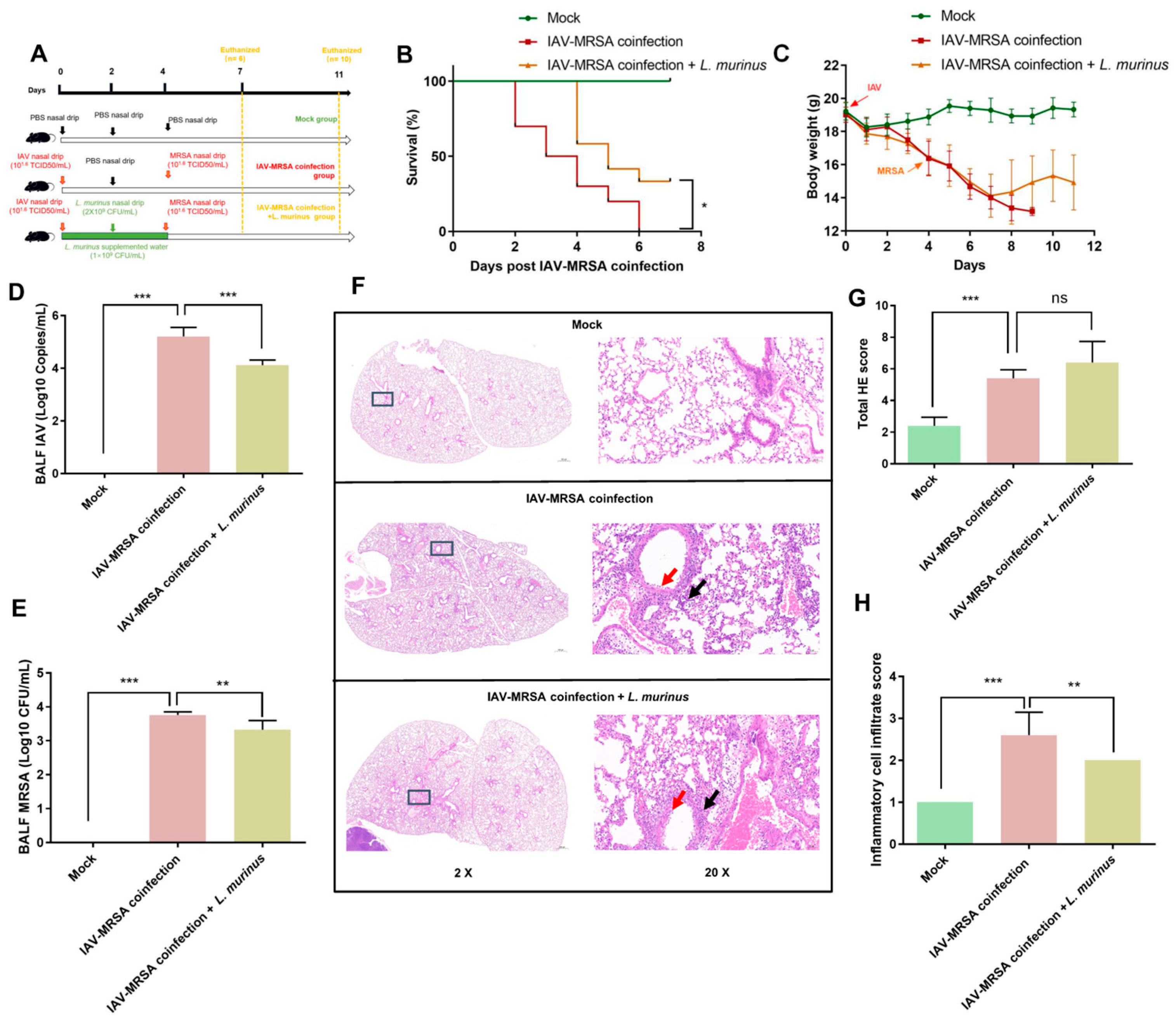
3.1. L. murinus Supplementation Reduces Susceptibility to Secondary MRSA Infection in IAV-Infected Mice
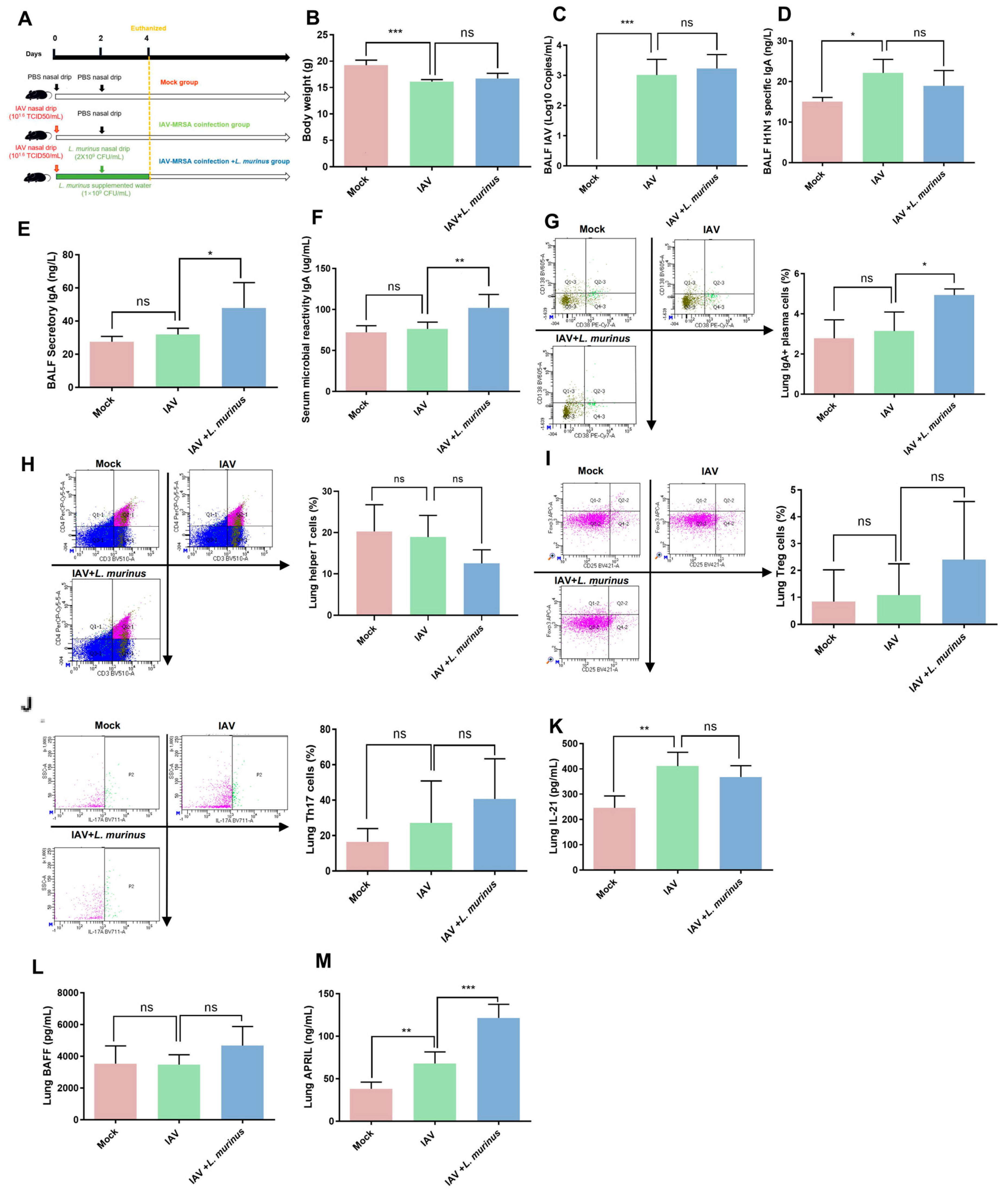
3.2. L. murinus Supplementation Enhanced Lung IgA Response and the Levels of TI IgA Pathway Markers in IAV Infected Mice
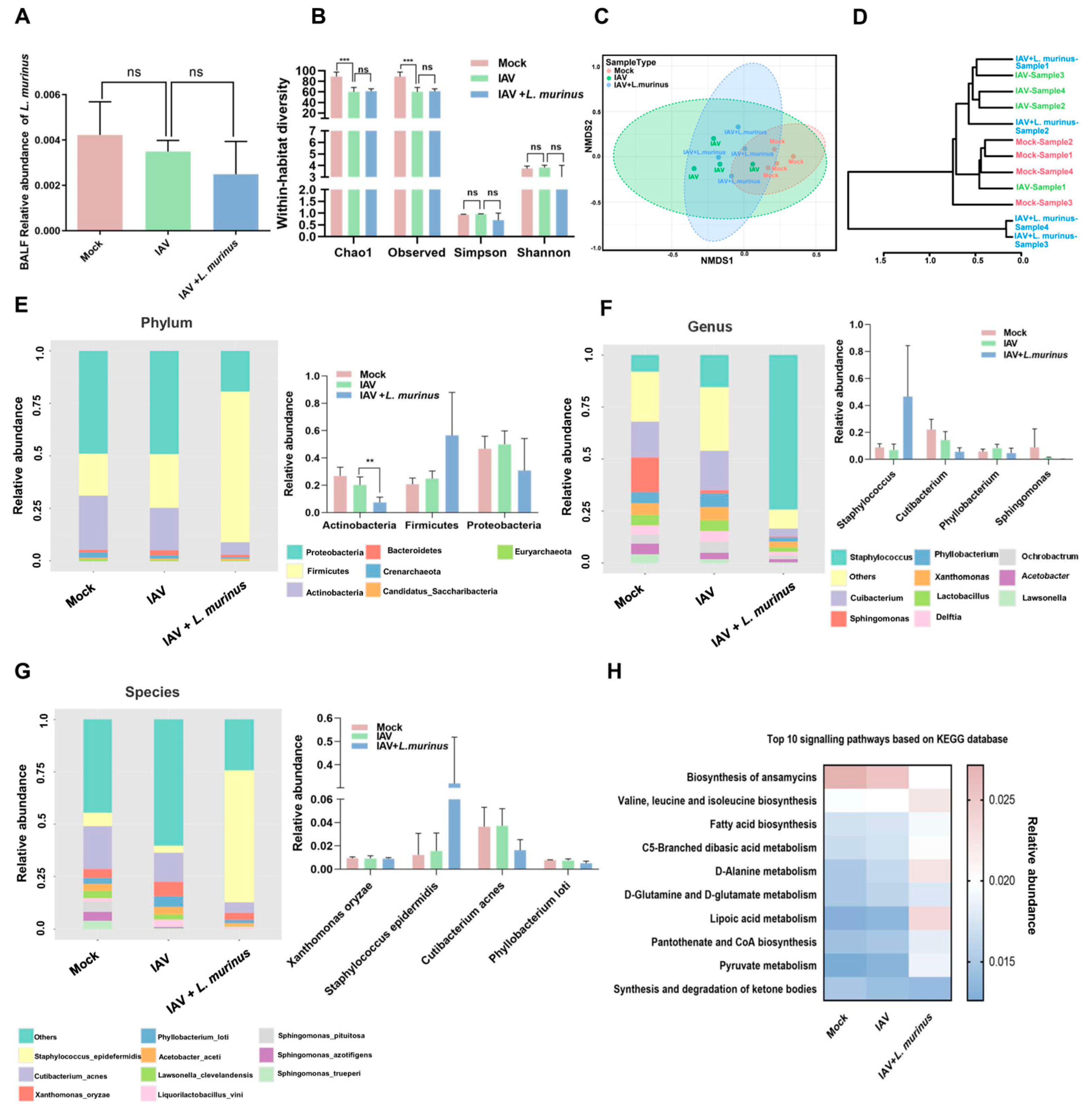
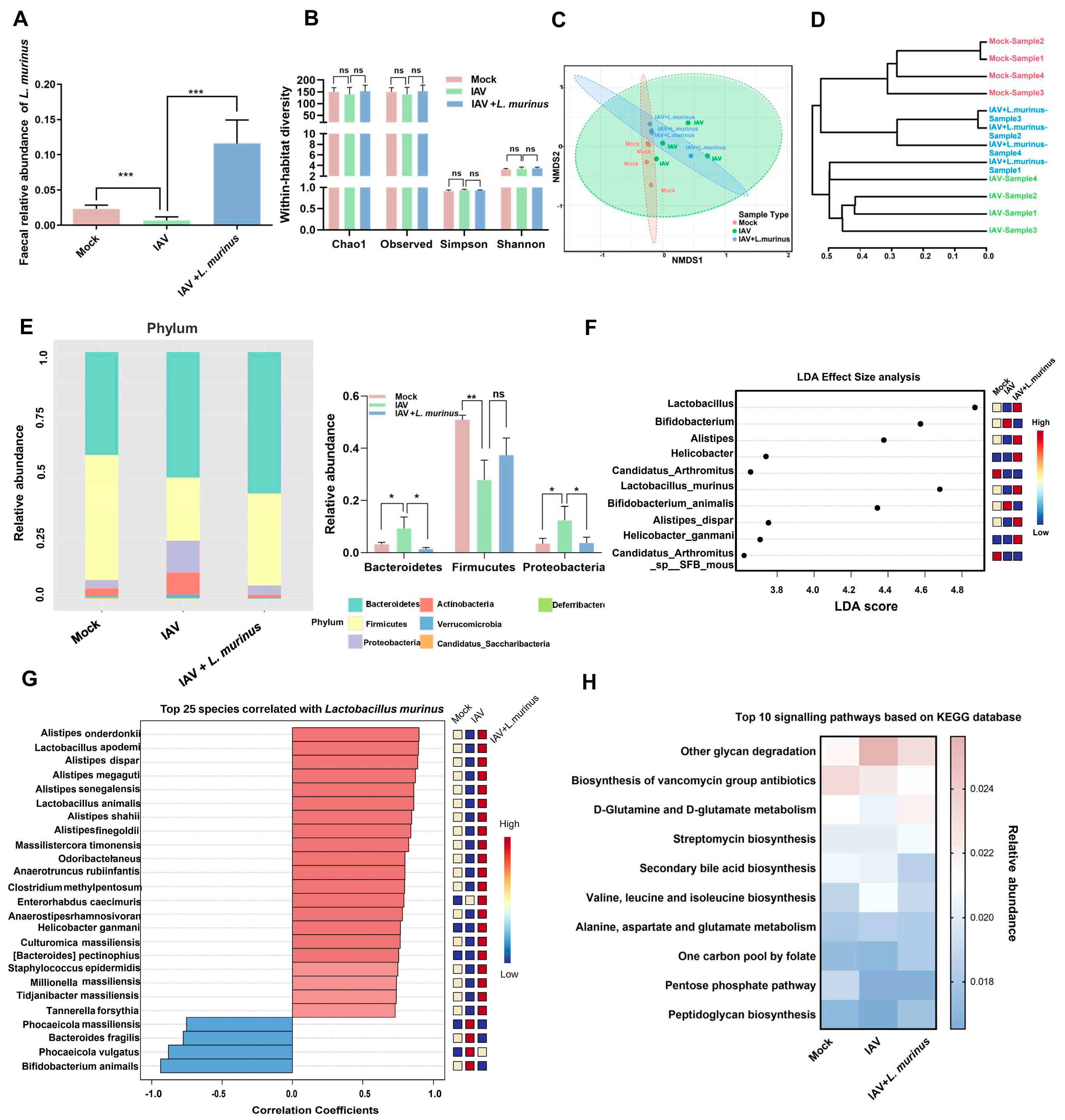
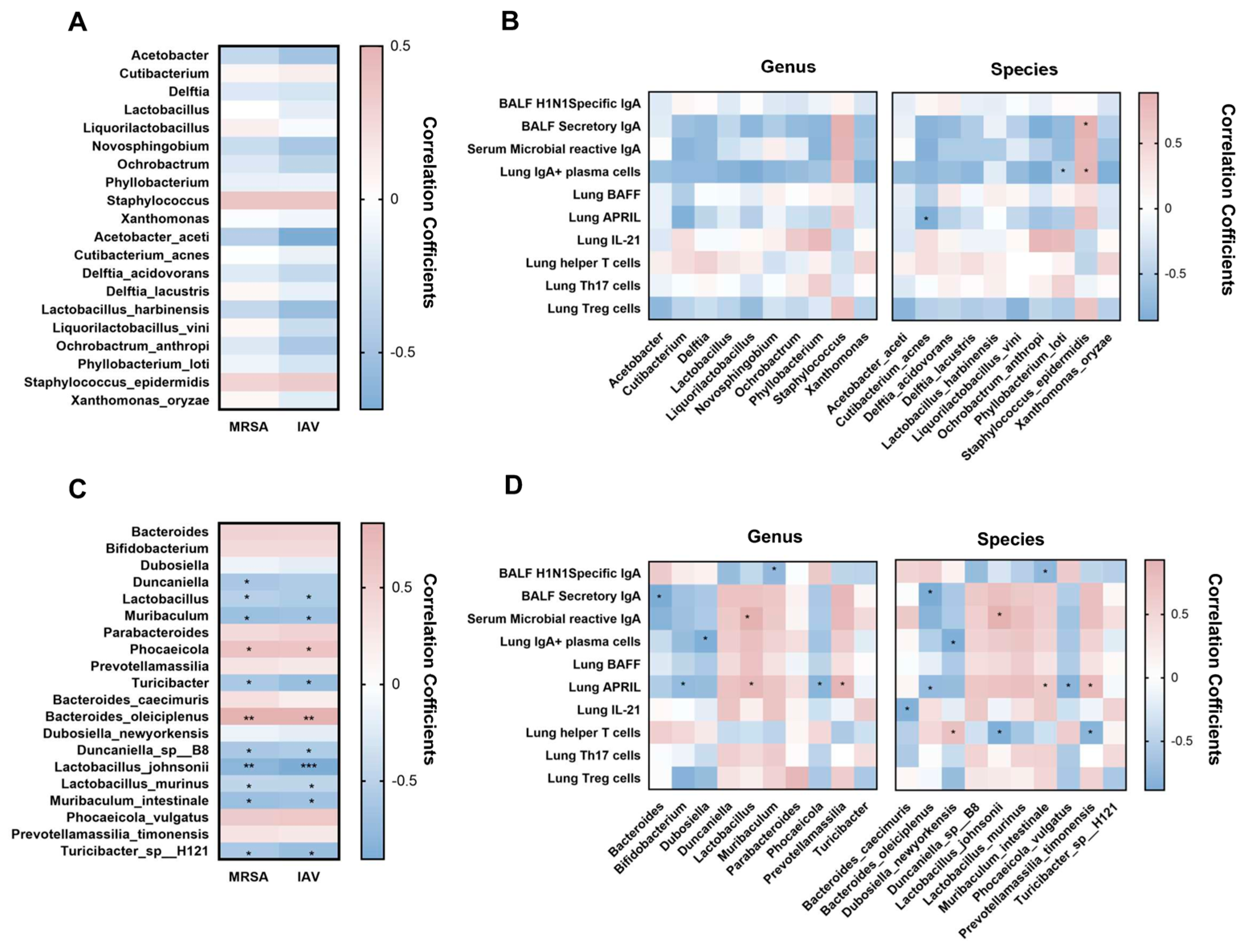
3.3. L. murinus Supplementation Reversed Gut Microbiota Dysbiosis, but Had No Effect on LRT Microbiota Dysbiosis in IAV-Infected Mice
3.4. Gut Microbiota and Lung IgA Response Show Correlation in Mice
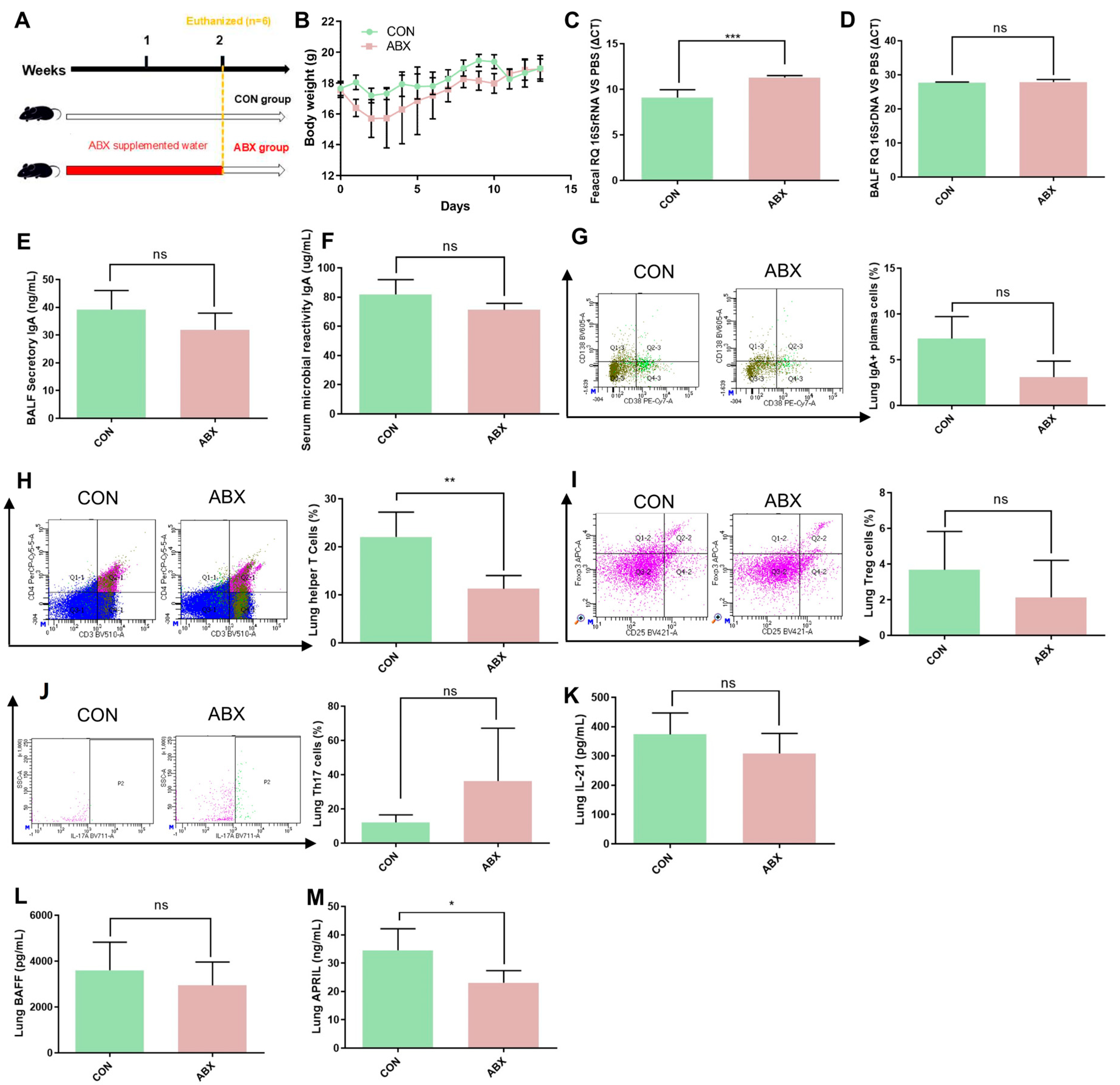
3.5. TI-IgA Response in the Lung Is Dependent on Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MRSA | Methicillin-resistant Staphylococcus aureus |
| IAV | Influenza A virus |
| BALF | Bronchoalveolar lavage fluid |
| SIgA | Secretory IgA |
| TD | T cell-dependent |
| TI | T cell-independent |
| BAFF | B-cell activating factor |
| APRIL | A proliferation-inducing ligand |
| PR8 | Influenza virus strain A/Puerto Rico/8/34 (H1N1) |
| TCID50 | 50% tissue culture infective dose |
| CFU | Colony forming units |
| SPF | Specific pathogen free |
| ABX | Antibiotic cocktails |
| H&E | Hematoxylin-eosin |
References
- Ciminski, K.; Chase, G.P.; Beer, M.; Schwemmle, M. Influenza A Viruses: Understanding Human Host Determinants. Trends Mol. Med. 2021, 27, 104–112. [Google Scholar] [CrossRef] [PubMed]
- WHO. Influenza (Seasonal). WHO. 2025. Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 28 February 2025).
- Klein, E.Y.; Monteforte, B.; Gupta, A.; Jiang, W.; May, L.; Hsieh, Y.H.; Dugas, A. The frequency of influenza and bacterial coinfection: A systematic review and meta-analysis. Influenza Other Respi. Viruses 2016, 10, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Randolph, A.G.; Xu, R.; Novak, T.; Newhams, M.M.; Wardenburg, J.B.; Weiss, S.L.; Sanders, R.C.; Thomas, N.J.; Hall, M.W.; Tarquinio, K.M.; et al. Vancomycin monotherapy may be insufficient to treat methicillin-resistant Staphylococcus aureus coinfection in children with influenza-related critical illness. Clin. Infect. Dis. 2019, 68, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, N.; Kitano, S.; Puah, G.R.Y.; Kittelmann, S.; Hwang, I.Y.; Chang, M.W. Microbiome and Human Health: Current Understanding, Engineering, and Enabling Technologies. Chem. Rev. 2023, 123, 31–72. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, M.; Guo, H.; Wang, K.; Liu, J.; Wang, Y.; Lin, Y.; Li, J.; Li, P.; Yang, L.; et al. Altered Respiratory Microbiomes, Plasma Metabolites, and Immune Responses in Influenza A Virus and Methicillin-Resistant Staphylococcus aureus Coinfection. Microbiol. Spectr. 2023, 11, e0524722. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, M.; Lin, Y.; Wang, K.; Li, J.; Li, P.; Yang, L.; Jia, L.; Zhang, B.; Guo, H.; et al. Topography of respiratory tract and gut microbiota in mice with influenza A virus infection. Front. Microbiol. 2023, 14, 1129690. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, S.; Bonifacio Lopes, J.P.P.; Bergé, M.; González-Ruiz, V.; Baud, D.; Kloehn, J.; Boal-Carvalho, I.; Schaeren, O.P.; Schotsaert, M.; Hathaway, L.J.; et al. Respiratory tissue-associated commensal bacteria offer therapeutic potential against pneumococcal colonization. Elife 2020, 9, e53581. [Google Scholar] [CrossRef] [PubMed]
- Bernard-Raichon, L.; Colom, A.; Monard, S.C.; Namouchi, A.; Cescato, M.; Garnier, H.; Leon-Icaza, S.A.; Métais, A.; Dumas, A.; Corral, D.; et al. A Pulmonary Lactobacillus murinus Strain Induces Th17 and RORγt+ Regulatory T Cells and Reduces Lung Inflammation in Tuberculosis. J. Immunol. 2021, 207, 1857–1870. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Deng, F.; Zhao, B.; Lin, Z.; Sun, Q.; Yang, X.; Wu, M.; Qiu, S.; Chen, Y.; Yan, Z.; et al. Lactobacillus murinus alleviate intestinal ischemia/reperfusion injury through promoting the release of interleukin-10 from M2 macrophages via Toll-like receptor 2 signaling. Microbiome 2022, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Corthésy, B. Role of secretory IgA in infection and maintenance of homeostasis. Autoimmun. Rev. 2013, 12, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A. The regulation of IgA class switching. Nat. Rev. Immunol. 2008, 8, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hu, S.; Yin, J.; Peng, X.; King, L.; Li, L.; Xu, Z.; Zhou, L.; Peng, Z.; Ze, X.; et al. Effect of synbiotic supplementation on immune parameters and gut microbiota in healthy adults: A double-blind randomized controlled trial. Gut Microbes 2023, 15, 2247025. [Google Scholar] [CrossRef] [PubMed]
- Vaisberg, M.; Paixão, V.; Almeida, E.B.; Santos, J.M.B.; Foster, R.; Rossi, M.; Pithon-Curi, T.C.; Gorjão, R.; Momesso, C.M.; Andrade, M.S.; et al. Daily intake of fermented milk containing lactobacillus casei shirota (lcs) modulates systemic and upper airways immune/inflammatory responses in marathon runners. Nutrients 2019, 11, 1678. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Cui, H.; Zhang, C.; Chen, Z.; Jiang, X.; Liu, J.; Wan, Z.; Li, J.; Liu, J.; Gao, Y.; et al. Aerosol Transmission of the Pandemic SARS-CoV-2 and Influenza A Virus Was Blocked by Negative Ions. Front. Cell. Infect. Microbiol. 2022, 12, 897416. [Google Scholar] [CrossRef] [PubMed]
- Papić, B.; Pate, M.; Henigman, U.; Zajc, U.; Gruntar, I.; Biasizzo, M.; Ocepek, M.; Kušar, D. New approaches on quantification of Campylobacter jejuni in poultry samples: The use of digital PCR and real-time PCR against the ISO standard plate count method. Front. Microbiol. 2017, 8, 331. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Gong, J.; Liu, F.; Li, B.; Li, Z.; You, J.; He, J.; Wu, S. Evaluation of an Antibiotic Cocktail for Fecal Microbiota Transplantation in Mouse. Front. Nutr. 2022, 9, 918098. [Google Scholar] [CrossRef] [PubMed]
- Robak, O.H.; Heimesaat, M.M.; Kruglov, A.A.; Prepens, S.; Ninnemann, J.; Gutbier, B.; Reppe, K.; Hochrein, H.; Suter, M.; Kirschning, C.J.; et al. Antibiotic treatment-induced secondary IgA deficiency enhances susceptibility to Pseudomonas aeruginosa pneumonia. J. Clin. Investig. 2018, 128, 3535–3545. [Google Scholar] [CrossRef] [PubMed]
- Renne, R.; Brix, A.; Harkema, J.; Herbert, R.; Kittel, B.; Lewis, D.; March, T.; Nagano, K.; Pino, M.; Rittinghausen, S.; et al. Proliferative and Nonproliferative Lesions of the Rat and Mouse Respiratory Tract. Toxicol. Pathol. 2009, 37, 5S–73S. [Google Scholar] [CrossRef] [PubMed]
- Curry, K.D.; Wang, Q.; Nute, M.G.; Tyshaieva, A.; Reeves, E.; Soriano, S.; Wu, Q.; Graeber, E.; Finzer, P.; Mendling, W.; et al. Emu: Species-Level Microbial Community Profiling for Full-Length Nanopore 16S Reads. Nat. Methods 2022, 19, 845. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhou, G.; Ewald, J.; Pang, Z.; Shiri, T.; Xia, J. MicrobiomeAnalyst 2.0: Comprehensive statistical, functional and integrative analysis of microbiome data. Nucleic Acids Res. 2023, 51, W310–W318. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, A.T.; Lyon De Ana, C.; Arafa, E.I.; Salwig, I.; Barker, K.A.; Korkmaz, F.T.; Ramanujan, A.; Etesami, N.S.; Soucy, A.M.; Martin, I.M.C.; et al. Antigen presentation by lung epithelial cells directs CD4+ TRM cell function and regulates barrier immunity. Nat. Commun. 2021, 12, 5834. [Google Scholar] [CrossRef] [PubMed]
- Djukovic, A.; Garzón, M.J.; Canlet, C.; Cabral, V.; Lalaoui, R.; García-Garcerá, M.; Rechenberger, J.; Tremblay-Franco, M.; Peñaranda, I.; Puchades-Carrasco, L.; et al. Lactobacillus supports Clostridiales to restrict gut colonization by multidrug-resistant Enterobacteriaceae. Nat. Commun. 2022, 13, 5617. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Colón, G.J.; Warheit-Niemi, H.; Gurczynski, S.J.; Taylor, Q.M.; Wilke, C.A.; Podsiad, A.B.; Crespo, J.; Bhan, U.; Moore, B.B. Influenza-induced immune suppression to methicillin-resistant Staphylococcus aureus is mediated by TLR9. PLoS Pathog. 2019, 15, e1007560. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.; Han, S.; Jang, I.H.; Ryu, J.; Rha, M.S.; Cho, H.J.; Yoon, S.S.; Nam, K.T.; Kim, C.H.; Park, M.S.; et al. Airway epithelial CD47 plays a critical role in inducing influenza virus-mediated bacterial super-infection. Nat. Commun. 2024, 15, 3666. [Google Scholar] [CrossRef] [PubMed]
- Møller, K.V.; Nguyen, H.T.T.; Mørch, M.G.M.; Hesselager, M.O.; Mulder, F.A.A.; Fuursted, K.; Olsen, A. A Lactobacilli diet that confers MRSA resistance causes amino acid depletion and increased antioxidant levels in the C. elegans host. Front. Microbiol. 2022, 13, 886206. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A.; Castro-Dopico, T.; Clatworthy, M.R. B cell class switching in intestinal immunity in health and disease. Scand. J. Immunol. 2022, 95, e13139. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Magri, G.; Grasset, E.K.; Cerutti, A. Rethinking mucosal antibody responses: IgM, IgG and IgD join IgA. Nat. Rev. Immunol. 2020, 20, 427–441. [Google Scholar] [CrossRef] [PubMed]
- Myette, J.R.; Kano, T.; Suzuki, H.; Sloan, S.E.; Szretter, K.J.; Ramakrishnan, B.; Adari, H.; Deotale, K.D.; Engler, F.; Shriver, Z.; et al. A Proliferation Inducing Ligand (APRIL) targeted antibody is a safe and effective treatment of murine IgA nephropathy. Kidney Int. 2019, 96, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Huus, K.E.; Petersen, C.; Finlay, B.B. Diversity and dynamism of IgA−microbiota interactions. Nat. Rev. Immunol. 2021, 21, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Mulder, M.; Radjabzadeh, D.; Kiefte-de Jong, J.C.; Uitterlinden, A.G.; Kraaij, R.; Stricker, B.H.; Verbon, A. Long-term effects of antimicrobial drugs on the composition of the human gut microbiota. Gut Microbes 2020, 12, 1795492. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Liu, X.; Cui, Z.; Zheng, Y.; Jiang, H.; Zhang, Y.; Li, Z.; Liang, Y.; Zhu, S.; Chu, P.K.; et al. Probiotic-based nanoparticles for targeted microbiota modulation and immune restoration in bacterial pneumonia. Natl. Sci. Rev. 2023, 10, nwac221. [Google Scholar] [CrossRef] [PubMed]
- Woodworth, M.H.; Conrad, R.E.; Haldopoulos, M.; Pouch, S.M.; Babiker, A.; Mehta, A.K.; Sitchenko, K.L.; Wang, C.H.; Strudwick, A.; Ingersoll, J.M.; et al. Fecal microbiota transplantation promotes reduction of antimicrobial resistance by strain replacement. Sci. Transl. Med. 2023, 15, eabo2750. [Google Scholar] [CrossRef] [PubMed]
- Rajeeve, K.; Vollmuth, N.; Janaki-Raman, S.; Wulff, T.F.; Baluapuri, A.; Dejure, F.R.; Huber, C.; Fink, J.; Schmalhofer, M.; Schmitz, W.; et al. Reprogramming of host glutamine metabolism during Chlamydia trachomatis infection and its key role in peptidoglycan synthesis. Nat. Microbiol. 2020, 5, 1390–1402. [Google Scholar] [CrossRef] [PubMed]
- Tsujii, A.; Takahashi, K.; Harada, H.; Kawashima, S.; Oikawa, H.; Fukushima, H.; Hayakawa, Y.; Koizumi, J.; Inoue, N.; Koshizuka, T. Evaluation of the protective effect of the intranasal vaccines adjuvanted with bacterium-like particles against intestinal infection. Vaccine 2024, 42, 125975. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, S.; Zhou, Y.; Disoma, C.; Dong, Z.; Du, A.; Zhang, Y.; Chen, Y.; Huang, W.; Chen, J.; et al. Microbiome Profiling Using Shotgun Metagenomic Sequencing Identified Unique Microorganisms in COVID-19 Patients with Altered Gut Microbiota. Front. Microbiol. 2021, 12, 712081. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, X.; Nie, C.; Wu, Y.; Luo, R.; Chen, C.; Niu, J.; Zhang, W. Effects of two strains of Lactobacillus isolated from the feces of calves after fecal microbiota transplantation on growth performance, immune capacity, and intestinal barrier function of weaned calves. Front. Microbiol. 2023, 14, 1249628. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q.; Lin, Y.; Wang, K.; Li, J.; Li, P.; Song, H. Lactobacillus murinus Reduces Susceptibility to Secondary MRSA Infection in IAV-Infected Mice Through Promoting a T Cell-Independent IgA Response. Microorganisms 2025, 13, 1709. https://doi.org/10.3390/microorganisms13071709
Chen Q, Lin Y, Wang K, Li J, Li P, Song H. Lactobacillus murinus Reduces Susceptibility to Secondary MRSA Infection in IAV-Infected Mice Through Promoting a T Cell-Independent IgA Response. Microorganisms. 2025; 13(7):1709. https://doi.org/10.3390/microorganisms13071709
Chicago/Turabian StyleChen, Qichao, Yanfeng Lin, Kaiying Wang, Jinhui Li, Peng Li, and Hongbin Song. 2025. "Lactobacillus murinus Reduces Susceptibility to Secondary MRSA Infection in IAV-Infected Mice Through Promoting a T Cell-Independent IgA Response" Microorganisms 13, no. 7: 1709. https://doi.org/10.3390/microorganisms13071709
APA StyleChen, Q., Lin, Y., Wang, K., Li, J., Li, P., & Song, H. (2025). Lactobacillus murinus Reduces Susceptibility to Secondary MRSA Infection in IAV-Infected Mice Through Promoting a T Cell-Independent IgA Response. Microorganisms, 13(7), 1709. https://doi.org/10.3390/microorganisms13071709