
New Findings on the Survival of Durusdinium glynnii Under Different Acclimation Methods to Low Salinities

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Experimental Design and Cultivation Conditions
2.3. Growth Parameters
2.4. Cell Morphotype
2.5. Statistical Analyses
3. Results
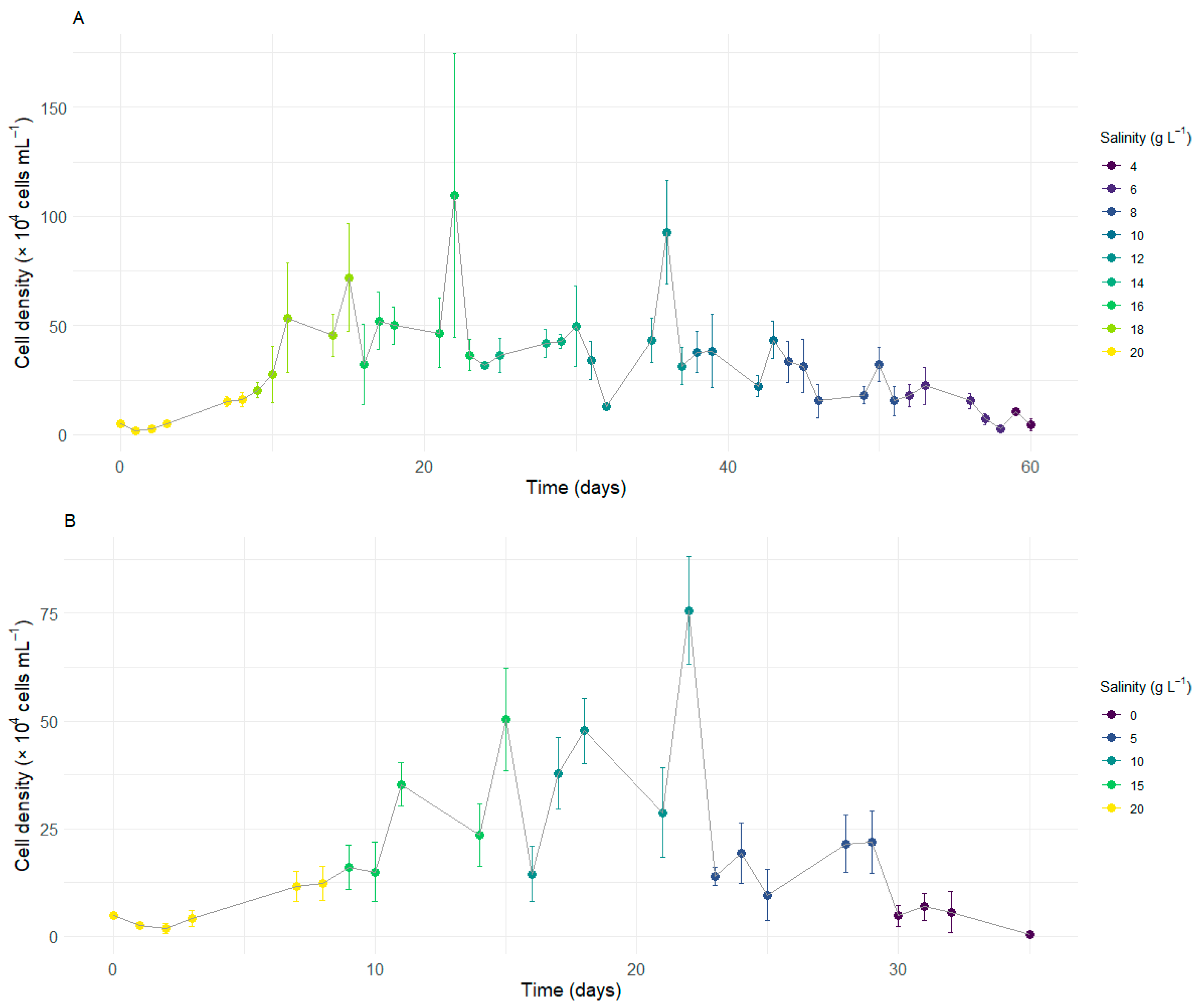
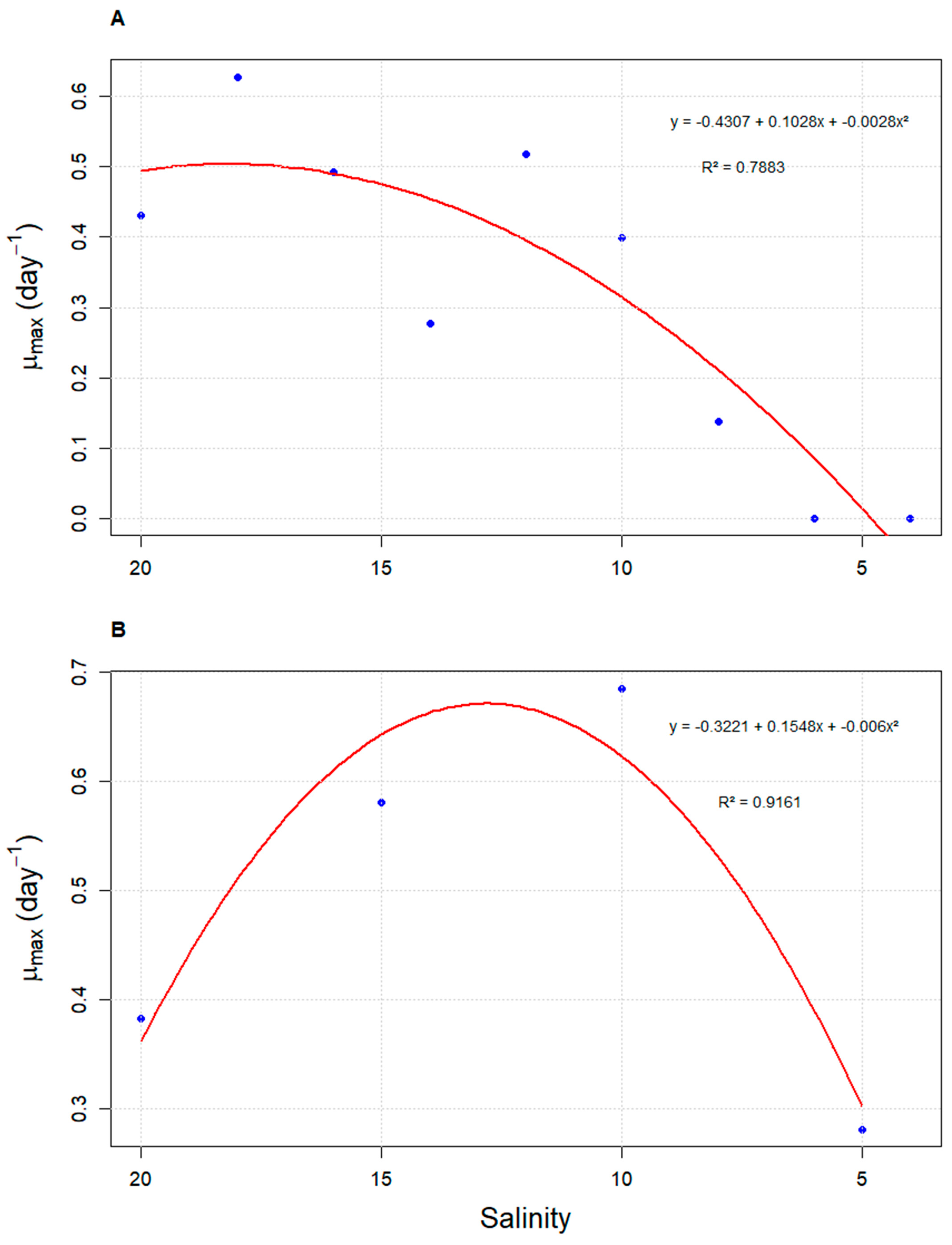
3.1. Growth Kinetics of Durusdinium glynnii Under Different Salinities
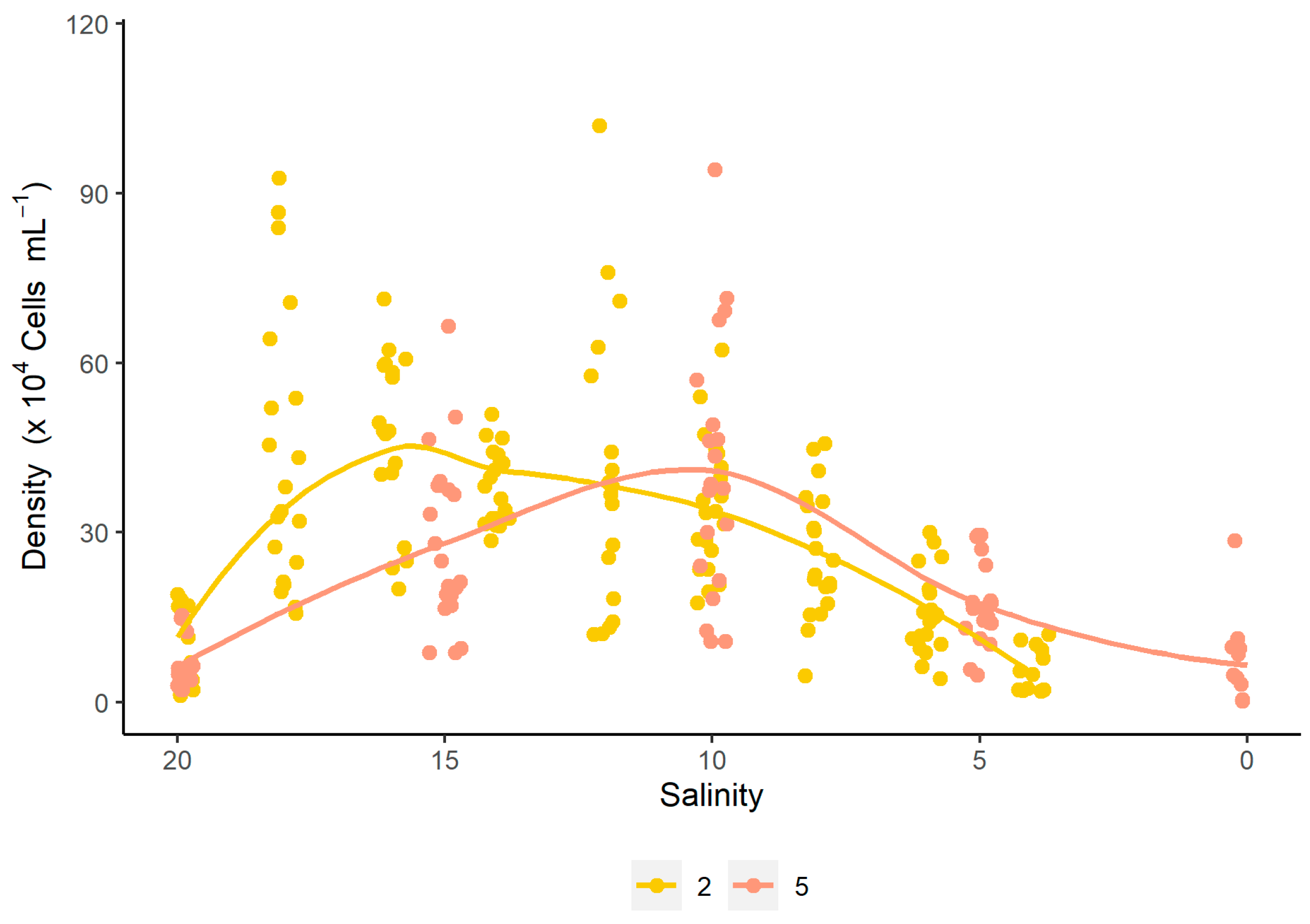
3.2. Growth of D. glynnii Under Strategies of Gradual Salinity Reduction
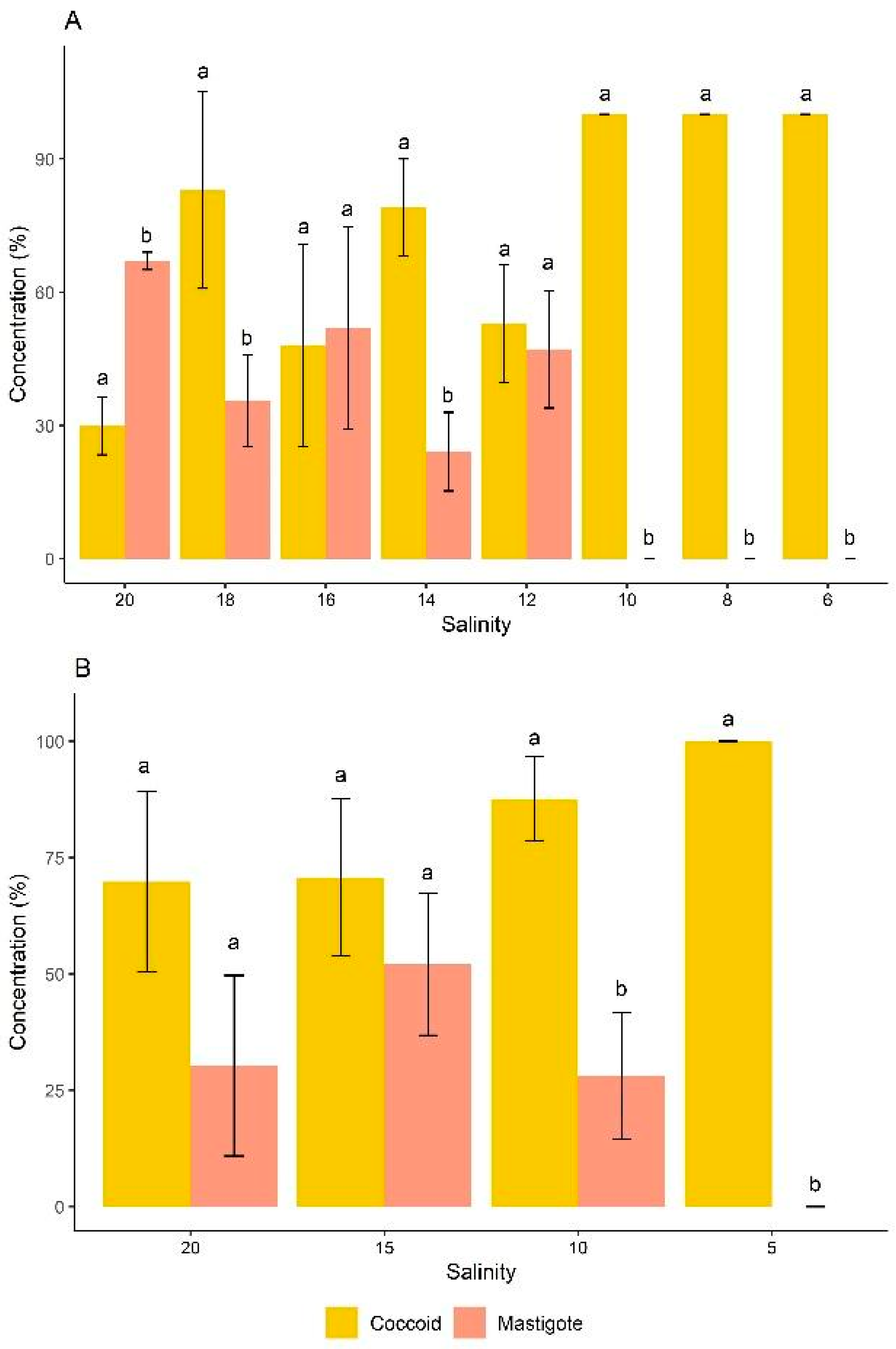
3.3. Effects of Salinite on the Cell Morphotype
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carty, S.; Parrow, M.W. Dinoflagellates. In Freshwater Algae of North America, 2nd ed.; Wehr, J.D., Sheath, R.G., Kociolek, J.P., Eds.; Elsevier: Ann Arbor, MI, USA, 2015; pp. 773–807. [Google Scholar]
- Oliveira, C.Y.B.; Oliveira, C.D.L.; Müller, M.N.; Santos, E.P.; Dantas, D.M.; Gálvez, A.O. A scientometric overview of global dinoflagellate research. Publications 2020, 8, 50. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Curr. Biol. 2018, 28, 2570–2580.e6. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.-H.; Jang, J.Y.; Kim, J.; Kim, S.; Keshavmurthy, S.; Agostini, S.; Reimer, J.D.; Chen, C.A.; Choi, K.-S.; Park, S.R.; et al. The Origin of the Subtropical Coral Alveopora japonica (Scleractinia: Acroporidae) in High-Latitude Environments. Front. Ecol. Evol. 2020, 8, 12. [Google Scholar] [CrossRef]
- Kannan, M.R.; Balakrishnan, R.; Thillaichidambaram, M.; Natesan, S.; Paramasamy, G.; Prakash, S.; Muthiah, R.C. Probing the Thermo-Tolerant Endosymbiont Genus Durusdinium (Clade D) in the Scleractinian Corals of Palk Bay, Southeast Coast of India. Biologia 2023, 78, 255–264. [Google Scholar] [CrossRef]
- Gordon, B.R.; Leggat, W. Symbiodinium–Invertebrate Symbioses and the Role of Metabolomics. Mar. Drugs 2010, 8, 2546–2568. [Google Scholar] [CrossRef]
- Ren, Y.; Sun, H.; Deng, J.; Huang, J.; Chen, F. Carotenoid Production from Microalgae: Biosynthesis, Salinity Responses and Novel Biotechnologies. Mar. Drugs 2021, 19, 713. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Galal, H.R.; Mousa, A.S.H.; Farghl, A.A.M. Impact of Macronutrients and Salinity Stress on Biomass and Biochemical Constituents in Monoraphidium braunii to Enhance Biodiesel Production. Sci. Rep. 2024, 14, 2725. [Google Scholar] [CrossRef]
- Röthig, T.; Trevathan-Tackett, S.M.; Voolstra, C.R.; Ross, C.; Chaffron, S.; Durack, P.J.; Warmuth, L.M.; Sweet, M. Human-induced salinity changes impact marine organisms and ecosystems. Glob. Chang. Biol. 2023, 29, 4731–4749. [Google Scholar] [CrossRef]
- Abadie, E.; Muguet, A.; Berteaux, T.; Chomérat, N.; Hess, P.; D’OrbCastel, E.R.; Masseret, E.; Laabir, M. Toxin and Growth Responses of the Neurotoxic Dinoflagellate Vulcanodinium rugosum to Varying Temperature and Salinity. Toxins 2016, 8, 136. [Google Scholar] [CrossRef]
- Čalasan, A.Ž.; Kretschmann, J.; Gottschling, M. They Are Young, and They Are Many: Dating Freshwater Lineages in Unicellular Dinophytes. Environ. Microbiol. 2019, 21, 4125–4135. [Google Scholar] [CrossRef]
- Annenkova, N.V.; Hansen, G.; Rengefors, K. Closely Related Dinoflagellate Species in Vastly Different Habitats—An Example of a Marine–Freshwater Transition. Eur. J. Phycol. 2020, 55, 478–489. [Google Scholar] [CrossRef]
- Tawong, W.; Yoshimatsu, T.; Yamaguchi, H.; Adachi, M. Effects of Temperature, Salinity and Their Interaction on Growth of Benthic Dinoflagellates Ostreopsis spp. from Thailand. Harmful Algae 2015, 44, 37–45. [Google Scholar] [CrossRef]
- Heyerhoff, B.; Nguyen, T.; Hillebrand, H.; Rhiel, E. On the Morphology and Predatory Behavior of the Dinoflagellate Oxyrrhis marina Exposed to Reduced Salinity. Eur. J. Protistol. 2019, 68, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Kholssi, R.; Lougraimzi, H.; Moreno-Garrido, I. Effects of Global Environmental Change on Microalgal Photosynthesis, Growth and Their Distribution. Mar. Environ. Res. 2023, 184, 105877. [Google Scholar] [CrossRef]
- Chaloub, R.M.; Monteiro, V.H.S.; Giordano, M. Physiological Responses of the Marine Microalga Isochrysis galbana (Prymnesiophyceae) to Hyposalinity. J. Appl. Phycol. 2023, 35, 2789–2800. [Google Scholar] [CrossRef]
- Honarparvar, S.; Zhang, X.; Chen, T.; Na, C.; Reible, D. Modeling technologies for desalination of brackish water—Toward a sustainable water supply. Curr. Opin. Chem. Eng. 2019, 26, 104–111. [Google Scholar] [CrossRef]
- Oliveira, C.Y.B.; Abreu, J.L.; Santos, E.P.; Matos, Â.P.; Tribuzi, G.; Oliveira, C.D.L.; Veras, B.O.; Bezerra, R.S.; Müller, M.N.; Gálvez, A.O. Light induces peridinin and docosahexaenoic acid accumulation in the dinoflagellate Durusdinium glynnii. Appl. Microbiol. Biotechnol. 2022, 106, 6263–6276. [Google Scholar] [CrossRef]
- Oliveira, C.Y.B.; Abreu, J.L.; Brandão, B.C.; Oliveira, D.W.S.; de Sena, P.R.; da Silva, W.A.; Araújo, E.S.; Rörig, L.R.; Costa, G.K.A.; Silva, S.M.B.C.; et al. A Holistic Approach to Producing Anti-Vibrio Metabolites by an Endosymbiotic Dinoflagellate Using Wastewater from Shrimp Rearing. Microorganisms 2024, 12, 1598. [Google Scholar] [CrossRef]
- Walne, P.R. Experiments in the Large-Scale Culture of the Larvae of Ostrea edulis. Fish Investig. 1966, 25, 1–53. [Google Scholar]
- Pindyck, R.S.; Rubinfeld, D.L. Econometric Models and Economic Forecasts; McGraw-Hill: New York, NY, USA, 1981; 630p. [Google Scholar]
- Stein, J.R. Handbook of Phycological Methods: Culture Methods and Growth Measurements; Cambridge University Press: London, UK, 1973; pp. 1–448. [Google Scholar]
- Matsubara, T.; Nagasoe, S.; Yamasaki, Y.; Shikata, T.; Shimasaki, Y.; Oshima, Y.; Honjo, T. Effects of temperature, salinity, and irradiance on the growth of the dinoflagellate Akashiwo sanguinea. J. Exp. Mar. Bio. Ecol. 2007, 342, 226–230. [Google Scholar] [CrossRef]
- Grzebyk, D.; Béchemin, C.; Ward, C.J.; Vérité, C.; Codd, G.A.; Maestrini, S.Y. Effects of salinity and two coastal waters on the growth and toxin content of the dinoflagellate Alexandrium minutum. J. Plankton Res. 2003, 25, 1185–1199. [Google Scholar] [CrossRef]
- Lim, P.-T.; Ogata, T. Salinity effect on growth and toxin production of four tropical Alexandrium species (Dinophyceae). Toxicon 2005, 45, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Zafar, A.M.; Javed, M.A.; Aly Hassan, A.; Mehmood, K.; Sahle-Demessie, E. Recent updates on ions and nutrients uptake by halotolerant freshwater and marine microalgae in conditions of high salinity. J. Water Process Eng. 2021, 44, 102382. [Google Scholar] [CrossRef]
- Bui, Q.T.N.; Kim, H.-S.; Ki, J.-S. Low salinity causes oxidative stress and modulates specific antioxidant gene expression in the toxic dinoflagellate Alexandrium pacificum. J. Appl. Phycol. 2022, 34, 2437–2447. [Google Scholar] [CrossRef]
- Klueter, A.; Trapani, J.; Archer, F.I.; McIlroy, S.E.; Coffroth, M.A. Comparative growth rates of cultured marine dinoflagellates in the genus Symbiodinium and the effects of temperature and light. PLoS ONE 2017, 12, e0187707. [Google Scholar] [CrossRef]
- Guimarães, S.C.P.; Rörig, L.R. Efeito da salinidade no crescimento dos dinoflagelados Prorocentrum micans Ehrenberg e Prorocentrum cf. obtusum ostenfeld isolados da costa catarinense—Brasil. Rev. Estud. Biologia 2004, 26, 29.36. [Google Scholar] [CrossRef]
- Oliveira, C.Y.B.; Brandão, B.C.S.; Jannuzzi, L.G.D.S.; Oliveira, D.W.S.; Yogui, G.T.; Müller, M.N.; Gálvez, A.O. New insights on the role of nitrogen in the resistance to environmental stress in an endosymbiotic dinoflagellate. Environ. Sci. Pollut. Res. 2023, 30, 82142–82151. [Google Scholar] [CrossRef]
- Martens, H.; Van de Waal, D.B.; Brandenburg, K.M.; Krock, B.; Tillmann, U. Salinity effects on growth and toxin production in an Alexandrium ostenfeldii (Dinophyceae) isolate from The Netherlands. J. Plankton Res. 2016, 38, 1302–1316. [Google Scholar] [CrossRef]
- Johnson, J.G.; Morey, J.S.; Neely, M.G.; Ryan, J.C.; Van Dolah, F.M. Transcriptome remodeling associated with chronological aging in the dinoflagellate, Karenia brevis. Mar. Genom. 2012, 5, 15–25. [Google Scholar] [CrossRef]
- Gao, Y.; Erdner, D.L. Insights into Stress-Induced Death Processes during Aging in the Marine Bloom-Forming Dinoflagellate Karenia brevis. J. Mar. Sci. Eng. 2022, 10, 1993. [Google Scholar] [CrossRef]
- González-Pech, R.A.; Stephens, T.G.; Chen, Y.; Mohamed, A.R.; Cheng, Y.; Shah, S.; Dougan, K.E.; Fortuin, M.D.A.; Lagorce, R.; Burt, D.W.; et al. Comparison of 15 dinoflagellate genomes reveals extensive sequence and structural divergence in family Symbiodiniaceae and genus Symbiodinium. BMC Biol. 2021, 19, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Novosel, N.; Mišić Radić, T.; Levak Zorinc, M.; Zemla, J.; Lekka, M.; Vrana, I.; Gašparović, B.; Horvat, L.; Kasum, D.; Legović, T.; et al. Salinity-induced chemical, mechanical, and behavioral changes in marine microalgae. J. Appl. Phycol. 2022, 34, 1293–1309. [Google Scholar] [CrossRef]
- Jeong, H.J.; Lee, S.Y.; Kang, N.S.; Du Yoo, Y.; Lim, A.S.; Lee, M.J.; Kim, H.S.; Yih, W.; Yamashita, H.; LaJeunesse, T.C. Genetics and Morphology Characterize the Dinoflagellate Symbiodinium voratum, n. sp., (Dinophyceae) as the Sole Representative of Symbiodinium Clade E. J. Eukaryot. Microbiol. 2013, 61, 75–94. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Salinity (g L−1) | |||
|---|---|---|---|---|
| 10 | 20 | 30 | 40 | |
| K (divisions day−1) | 0.27 ± 0.05 a | 0.23 ± 0.02 a | 0.23 ± 0.05 a | 0.42 ± 0.18 a |
| DT (days division−1) | 3.89 ± 0.81 ab | 4.32 ± 0.32 ab | 4.42 ± 0.88 a | 2.70 ± 0.96 b |
| µ (day−1) | 0.19 ± 0.06 c | 0.35 ± 0.03 b | 0.22 ± 0.03 bc | 0.62 ± 0.11 a |
| MCD (×104 cells mL−1) | 27.25 ± 9.28 a | 35.38 ± 12.79 a | 34.38 ± 13.42 a | 30.44 ± 6.34 a |
| dMCD (day) | 18 | 6 | 6 | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Cassia S. Brandão, B.; de Abreu, J.L.; Oliveira, D.W.S.; da Silva Campos, C.V.F.; de Aguiar, I.M.T.; de Sena, P.R.; Gálvez, A.O.; Oliveira, C.Y.B. New Findings on the Survival of Durusdinium glynnii Under Different Acclimation Methods to Low Salinities. Microorganisms 2025, 13, 946. https://doi.org/10.3390/microorganisms13040946
de Cassia S. Brandão B, de Abreu JL, Oliveira DWS, da Silva Campos CVF, de Aguiar IMT, de Sena PR, Gálvez AO, Oliveira CYB. New Findings on the Survival of Durusdinium glynnii Under Different Acclimation Methods to Low Salinities. Microorganisms. 2025; 13(4):946. https://doi.org/10.3390/microorganisms13040946
Chicago/Turabian Stylede Cassia S. Brandão, Barbara, Jéssika L. de Abreu, Deyvid Willame S. Oliveira, Clarissa Vilela F. da Silva Campos, Ingrid M. Thó de Aguiar, Pedro R. de Sena, Alfredo O. Gálvez, and Carlos Yure B. Oliveira. 2025. "New Findings on the Survival of Durusdinium glynnii Under Different Acclimation Methods to Low Salinities" Microorganisms 13, no. 4: 946. https://doi.org/10.3390/microorganisms13040946
APA Stylede Cassia S. Brandão, B., de Abreu, J. L., Oliveira, D. W. S., da Silva Campos, C. V. F., de Aguiar, I. M. T., de Sena, P. R., Gálvez, A. O., & Oliveira, C. Y. B. (2025). New Findings on the Survival of Durusdinium glynnii Under Different Acclimation Methods to Low Salinities. Microorganisms, 13(4), 946. https://doi.org/10.3390/microorganisms13040946