
Irrigation of Suaeda salsa with Saline Wastewater and Microalgae: Improving Saline–Alkali Soil and Revealing the Composition and Function of Rhizosphere Bacteria

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Sample and Irrigation Water Preparation
2.2. Experimental Design
2.3. Measurement of the Biomass of S. salsa and Rhizosphere Soil Properties
2.4. Analysis of Rhizosphere Soil Bacterial Communities
2.5. Data Processing and Statistical Analysis
3. Results
3.1. Suaeda Salsa Growth
3.2. Soil Physicochemical Properties
3.3. Soil Bacterial Community Diversity
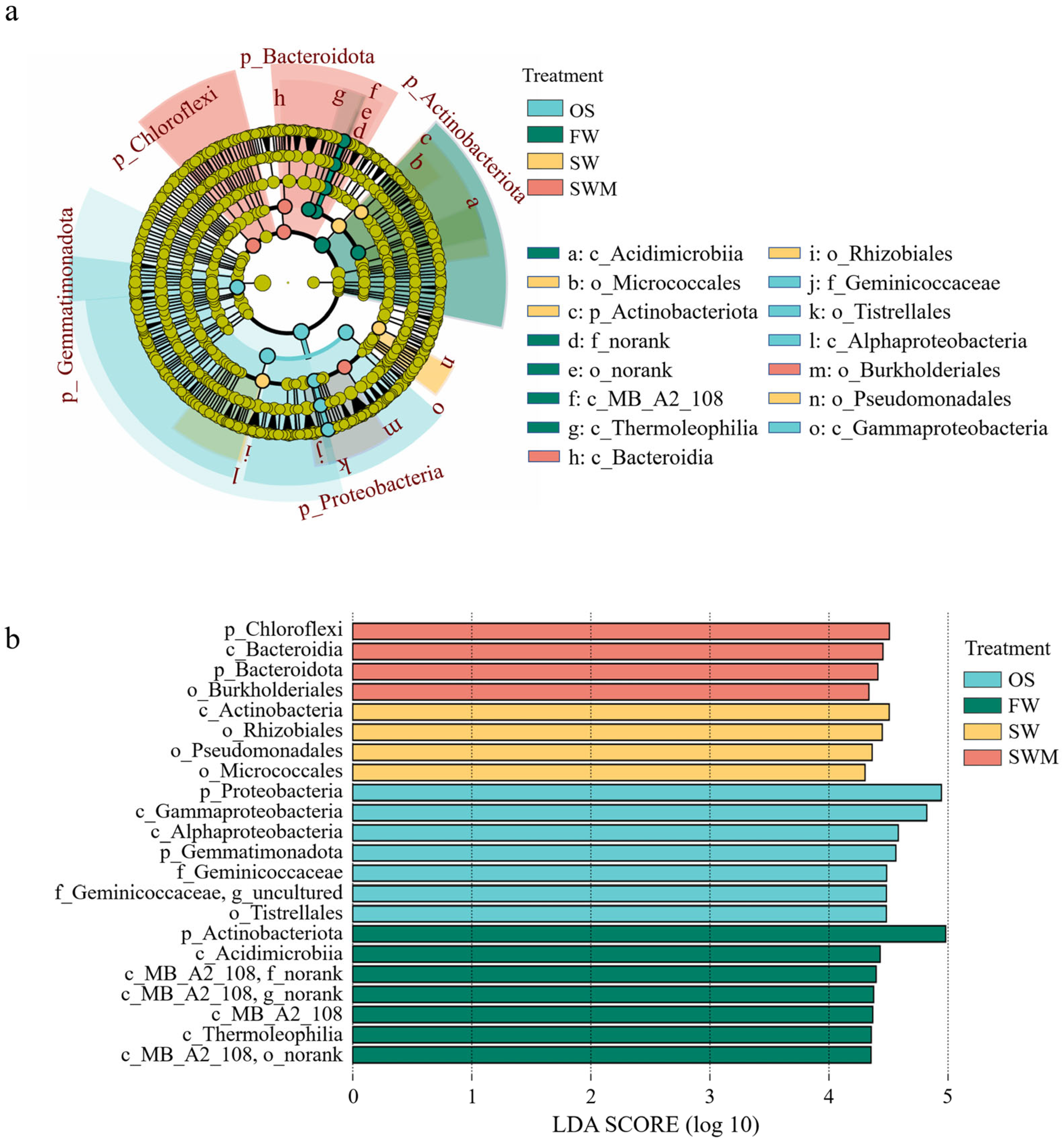
3.4. Soil Bacterial Community Composition
3.5. The Correlation Between Soil Bacterial Communities and Soil Physicochemical Properties
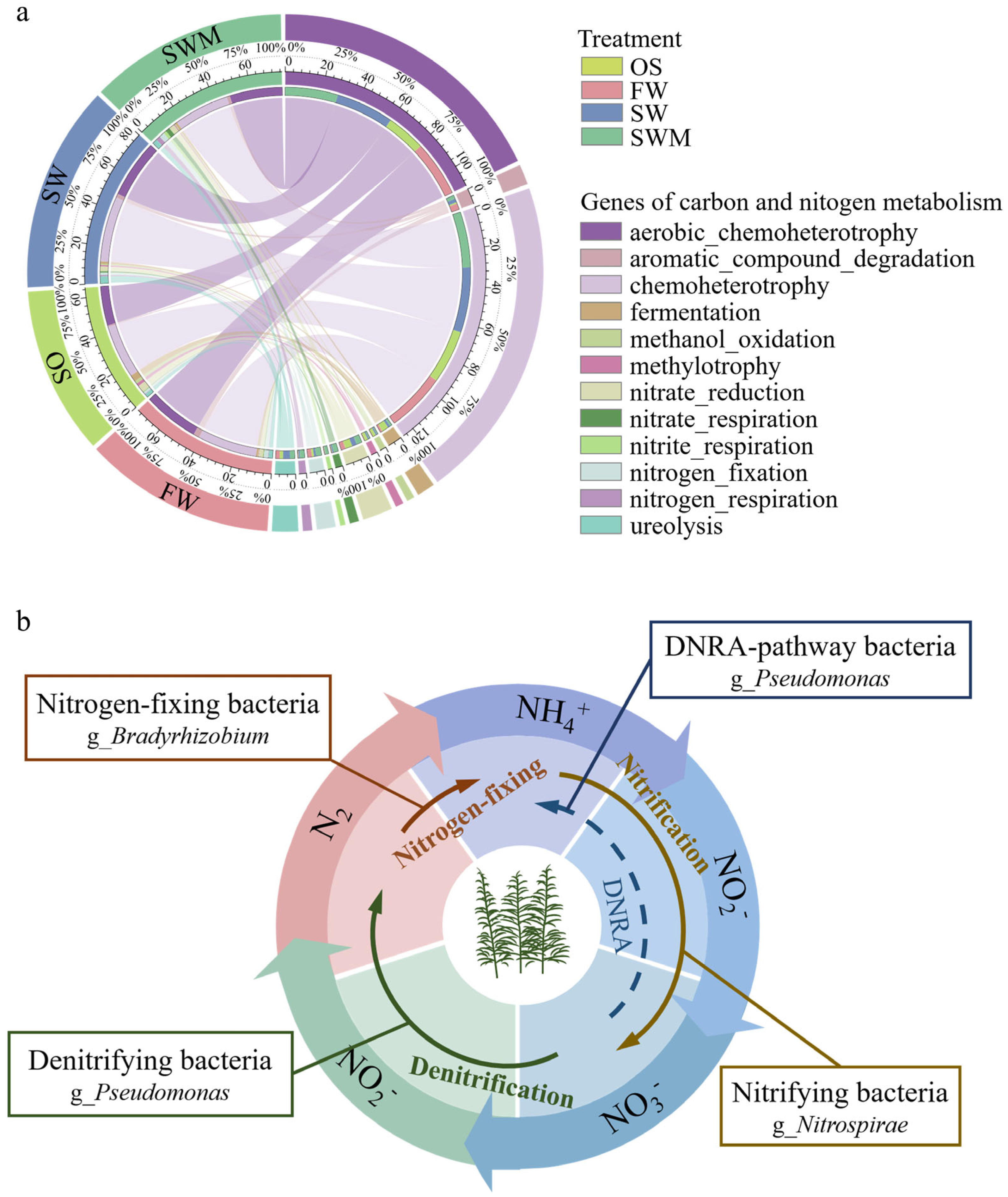
3.6. Soil Bacterial Community Function
3.6.1. Metabolic Pathways Based on PICRUSt2
3.6.2. Bacterial Community Functional Predictive Based on FAPROTAX
4. Discussion
4.1. Physicochemical Properties of the Rhizosphere Soil
4.2. Diversity and Composition of Bacterial Community
4.3. Influence of Soil Physicochemical Properties on Bacterial Community Composition
4.4. Functional Prediction of Rhizosphere Soil Bacteria
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ding, B.; Bai, Y.; Guo, S.; He, Z.; Wang, B.; Liu, H.; Zhai, J.; Cao, H. Effect of irrigation water salinity on soil characteristics and microbial communities in cotton fields in Southern Xinjiang, China. Agronomy 2023, 13, 1679. [Google Scholar] [CrossRef]
- Zhao, X.; Meng, T.; Jin, S.; Ren, K.; Cai, Z.; Cai, B.; Li, S. The salinity survival strategy of Chenopodium quinoa: Investigating microbial community shifts and nitrogen cycling in saline soils. Microorganisms 2023, 11, 2829. [Google Scholar] [CrossRef]
- Qadir, M.; Oster, J.D.; Schubert, S.; Noble, A.D.; Sahrawat, K.L. Phytoremediation of sodic and saline-sodic soils. Adv. Agron. 2007, 96, 197–247. [Google Scholar] [CrossRef]
- Gao, L.; Huang, Y.; Liu, Y.; Mohamed, O.A.A.; Fan, X.; Wang, L.; Li, L.; Ma, J. Bacterial community structure and potential microbial coexistence mechanism associated with three halophytes adapting to the extremely hypersaline environment. Microorganisms 2022, 10, 1124. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yu, S.; Yu, Z.; Ma, M.; Liu, M.; Pei, H. Phycoremediation potential of salt-tolerant microalgal species: Motion, metabolic characteristics, and their application for saline–alkali soil improvement in eco-farms. Microorganisms 2024, 12, 676. [Google Scholar] [CrossRef]
- Tang, L.; Zhan, L.; Han, Y.; Wang, Z.; Dong, L.; Zhang, Z. Microbial community assembly and functional profiles along the soil-root continuum of salt-tolerant Suaeda glauca and Suaeda salsa. Front. Plant Sci. 2023, 14, 1301117. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Xu, X.; Liu, J.; Han, J.; Lu, H. Planting Suaeda salsa improved the soil properties and bacterial community diversity in a coastal mudflat. Land Degrad. Dev. 2023, 34, 3262–3271. [Google Scholar] [CrossRef]
- Ma, C.; Lei, C.; Zhu, X.; Ren, C.; Liu, N.; Liu, Z.; Du, H.; Tang, T.; Li, R.; Cui, H. Saline-alkali land amendment and value development: Microalgal biofertilizer for efficient production of a halophytic crop-Chenopodium quinoa. Land Degrad. Dev. 2023, 34, 956–968. [Google Scholar] [CrossRef]
- Shayesteh, H.; Jenkins, S.N.; Moheimani, N.R.; Bolan, N.; Bühlmann, C.H.; Gurung, S.K.; Vadiveloo, A.; Bahri, P.A.; Mickan, B.S. Nitrogen dynamics and biological processes in soil amended with microalgae grown in abattoir digestate to recover nutrients. J. Environ. Manag. 2023, 344, 118467. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, T.; Shao, P.; Sun, J.; Xu, W.; Zhao, Y. Effects of short-term nitrogen addition on rhizosphere and bulk soil bacterial community structure of three halophytes in the Yellow River Delta. Land Degrad. Dev. 2023, 34, 3281–3294. [Google Scholar] [CrossRef]
- Zhou, T.; Zhang, L.; Yang, X.; Wu, Z.; Yang, Z.; Wang, J.; Chen, N.; Ren, X.; Hu, S. Prioritizing microbial functions over soil quality for enhanced multifunctionality in saline-sodic soil remediation. J. Environ. Manag. 2025, 379, 124731. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yasen, M.; Gong, M.; Zhou, Q.; Li, M. Structural variability in the rhizosphere bacterial communities of three halophytes under different levels of salinity-alkalinity. Plant Soil 2024, 502, 709–723. [Google Scholar] [CrossRef]
- Lin, H.; Liu, C.; Li, B.; Dong, Y. Trifolium repens L. regulated phytoremediation of heavy metal contaminated soil by promoting soil enzyme activities and beneficial rhizosphere associated microorganisms. J. Hazard. Mater. 2021, 402, 123829. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Jiang, Z.; Xiang, Z.; Zhou, A.; Wang, C.; Wang, Z.; Zhou, F. Genomic features of a plant growth-promoting endophytic Enterobacter cancerogenus JY65 dominant in microbiota of halophyte Suaeda salsa. Plant Soil. 2024, 496, 269–287. [Google Scholar] [CrossRef]
- Li, J.; Hussain, T.; Feng, X.; Guo, K.; Chen, H.; Yang, C.; Liu, X. Comparative study on the resistance of Suaeda glauca and Suaeda salsa to drought, salt, and alkali stresses. Ecol. Eng. 2019, 140, 105593. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, L.; Li, T.; Fu, Z.; Sun, J.; Hu, R.; Zhang, Y. Effects of short-term nitrogen and phosphorus addition on soil bacterial community of different halophytes. mSphere 2024, 9, e00226-24. [Google Scholar] [CrossRef]
- Ran, J.; Liu, X.; Hui, X.; Ma, Q.; Liu, J. Differentiating bacterial community responses to long-term phosphorus fertilization in wheat bulk and rhizosphere soils on the Loess Plateau. Appl. Soil Ecol. 2021, 166, 104090. [Google Scholar] [CrossRef]
- Liu, F.; Mo, X.; Kong, W.; Song, Y. Soil bacterial diversity, structure, and function of Suaeda salsa in rhizosphere and non-rhizosphere soils in various habitats in the Yellow River Delta, China. Sci. Total Environ. 2020, 740, 140144. [Google Scholar] [CrossRef]
- Zhang, S.; Pei, L.; Zhao, Y.; Shan, J.; Zheng, X.; Xu, G.; Sun, Y.; Wang, F. Effects of microplastics and nitrogen deposition on soil multifunctionality, particularly C and N cycling. J. Hazard. Mater. 2023, 451, 131152. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, J.; Li, T.; Shao, P.; Ma, J.; Dong, K. Effects of nitrogen and phosphorus imbalance input on rhizosphere and bulk soil bacterial community of Suaeda salsa in the Yellow River Delta. Front. Mar. Sci. 2023, 10, 1131713. [Google Scholar] [CrossRef]
- Li, Y.; Lin, Q.; Wang, S.; Li, X.; Liu, W.; Luo, C.; Zhang, Z.; Zhu, X.; Jiang, L.; Li, X. Soil bacterial community responses to warming and grazing in a Tibetan alpine meadow. FEMS Microbiol. Ecol. 2015, 92, fiv152. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Chen, Y.; Dou, X.; Liao, D.; Li, K.; An, C.; Li, G.; Dong, Z. Microbial fertilizers improve soil quality and crop yield in coastal saline soils by regulating soil bacterial and fungal community structure. Sci. Total Environ. 2024, 949, 175127. [Google Scholar] [CrossRef]
- Siddikee, M.A.; Zereen, M.I.; Li, C.; Dai, C. Endophytic fungus Phomopsis liquidambari and different doses of N-fertilizer alter microbial community structure and function in rhizosphere of rice. Sci. Rep. 2016, 6, 32270. [Google Scholar] [CrossRef]
- Li, X.; Sardans, J.; Hou, L.; Gao, D.; Liu, M.; Peñuelas, J. Dissimilatory nitrate/nitrite reduction processes in river sediments across climatic gradient: Influences of biogeochemical controls and climatic temperature regime. J. Geophys. Res. Biogeosci. 2019, 124, 2305–2320. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, C.; Wu, J.; Liu, X.; Xu, J. Impact of organic matter addition on pH change of paddy soils. J. Soils Sediments 2013, 13, 12–23. [Google Scholar] [CrossRef]
- Gr, S.; Yadav, R.K.; Chatrath, A.; Gerard, M.; Tripathi, K.; Govindsamy, V.; Abraham, G. Perspectives on the potential application of cyanobacteria in the alleviation of drought and salinity stress in crop plants. J. Appl. Phycol. 2021, 33, 3761–3778. [Google Scholar] [CrossRef]
- Dineshkumar, R.; Kumaravel, R.; Gopalsamy, J.; Sikder, M.N.A.; Sampathkumar, P. Microalgae as bio-fertilizers for rice growth and seed yield productivity. Waste Biomass Valorization 2018, 9, 793–800. [Google Scholar] [CrossRef]
- Mo, X.; Song, Y.; Chen, F.; You, C.; Li, D.; Liu, F. Replacement of plant communities altered soil bacterial diversity and structure rather than the function in similar habitats of the Yellow River Delta, China. Ecol. Indic. 2023, 146, 109793. [Google Scholar] [CrossRef]
- Singh, K. Microbial and enzyme activities of saline and sodic soils. Land Degrad. Dev. 2016, 27, 706–718. [Google Scholar] [CrossRef]
- Alvarez, A.L.; Weyers, S.L.; Goemann, H.M.; Peyton, B.M.; Gardner, R.D. Microalgae, soil and plants: A critical review of microalgae as renewable resources for agriculture. Algal Res. 2021, 54, 102200. [Google Scholar] [CrossRef]
- Zhang, L.; Li, J.; Bi, H.; Shi, Q.; Gong, B. Effects of fulvic acid on tomato yield and rhizosphere soil microecology under different soil pH and phosphorus levels. China Veg. 2021, 2021, 45–52. [Google Scholar] [CrossRef]
- Li, Y.; Kang, E.; Song, B.; Wang, J.; Zhang, X.; Wang, J.; Li, M.; Yan, L.; Yan, Z.; Zhang, K.; et al. Soil salinity and nutrients availability drive patterns in bacterial community and diversity along succession gradient in the Yellow River Delta. Estuarine, Coast. Shelf Sci. 2021, 262, 107621. [Google Scholar] [CrossRef]
- Chi, Z.; Zhu, Y.; Li, H.; Wu, H.; Yan, B. Unraveling bacterial community structure and function and their links with natural salinity gradient in the Yellow River Delta. Sci. Total Environ. 2021, 773, 145673. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Andersen, M.N.; Qi, X.; Li, P.; Li, Z.; Fan, X.; Zhou, Y. Effects of reclaimed water irrigation and nitrogen fertilization on the chemical properties and microbial community of soil. J. Integr. Agric. 2017, 16, 679–690. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, F.; Abalos, D.; Luo, Y.; Hui, D.; Hungate, B.A.; García-Palacios, P.; Kuzyakov, Y.; Olesen, J.E.; Jørgensen, U.; et al. Stimulation of ammonia oxidizer and denitrifier abundances by nitrogen loading: Poor predictability for increased soil N2O emission. Glob. Change Biol. 2022, 28, 2158–2168. [Google Scholar] [CrossRef]
- Cai, J.; Fan, J.; Liu, X.; Sun, K.; Wang, W.; Zhang, M.; Li, H.; Xu, H.; Kong, W.; Yu, F. Biochar-amended coastal wetland soil enhances growth of Suaeda salsa and alters rhizosphere soil nutrients and microbial communities. Sci. Total Environ. 2021, 788, 147707. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yu, Q.; Bai, S.; Wang, M.; Sun, W.; Xu, S.; Shi, X.; Lu, J.; Xie, X.; Qiu, W. Soil organic carbon impact on soil physical properties through quantity and quality modifications. Soil Adv. 2024, 2, 100014. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, K.; Yu, J.; Zhou, D.; Li, Y.; Guan, B.; Yu, Y.; Wang, X.; Ren, Z.; Wang, W.; et al. Soil bacterial community structure in different micro-habitats on the tidal creek section in the Yellow River Estuary. Front. Ecol. Evol. 2022, 10, 950605. [Google Scholar] [CrossRef]
- Hao, H.; Yue, Y.; Chen, Q.; Yang, Y.; Kuai, B.; Wang, Q.; Xiao, T.; Chen, H.; Zhang, J. Effects of an efficient straw decomposition system mediated by Stropharia rugosoannulata on soil properties and microbial communities in forestland. Sci. Total Environ. 2024, 916, 170266. [Google Scholar] [CrossRef]
- Qu, Z.; Li, Y.; Xu, W.; Chen, W.; Hu, Y.; Wang, Z. Different genotypes regulate the microbial community structure in the soybean rhizosphere. J. Integr. Agric. 2023, 22, 585–597. [Google Scholar] [CrossRef]
- Khalid, M.; Du, B.; Tan, H.; Liu, X.; Su, L.; Saeed ur, R.; Ali, M.; Liu, C.; Sun, N.; Hui, N. Phosphorus elevation erodes ectomycorrhizal community diversity and induces divergence of saprophytic community composition between vegetation types. Sci. Total Environ. 2021, 793, 148502. [Google Scholar] [CrossRef]
- Xiao, C.; Guo, S.; Wang, Q.; Chi, R. Enhanced reduction of lead bioavailability in phosphate mining wasteland soil by a phosphate-solubilizing strain of Pseudomonas sp., LA, coupled with ryegrass (Lolium perenne L.) and sonchus (Sonchus oleraceus L.). Environ. Pollut. 2021, 274, 116572. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and interactions in plant growth promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Kakumanu, M.L.; Ma, L.; Williams, M.A. Drought-induced soil microbial amino acid and polysaccharide change and their implications for C-N cycles in a climate change world. Sci. Rep. 2019, 9, 10968. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Chen, Y.; Lu, P.; Liu, M.; Sun, P.; Zhang, Z. Roles of endophytic bacteria in Suaeda salsa grown in coastal wetlands: Plant growth characteristics and salt tolerance mechanisms. Environ. Pollut. 2021, 287, 117641. [Google Scholar] [CrossRef] [PubMed]
- Becker, E.A.; Seitzer, P.M.; Tritt, A.; Larsen, D.; Krusor, M.; Yao, A.I.; Wu, D.; Madern, D.; Eisen, J.A.; Darling, A.E.; et al. Phylogenetically driven sequencing of extremely halophilic archaea reveals strategies for static and dynamic osmo-response. PLoS Genet. 2014, 10, e1004784. [Google Scholar] [CrossRef]
- Tao, J.; Wang, S.; Liao, T.; Luo, H. Evolutionary origin and ecological implication of a unique nif island in free-living Bradyrhizobium lineages. ISME J. 2021, 15, 3195–3206. [Google Scholar] [CrossRef]
- An, X.; Wang, Z.; Jiao, K.; Teng, X.; Zhou, R.; Xu, M.; Lian, B. Bacterial community characteristics in the rhizosphere of Suaeda glauca versus bulk soil in coastal silt soil modified by sea-sand and their implications. Front. Mar. Sci. 2023, 9, 1001449. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Treatment | |||
|---|---|---|---|---|
| OS | FW | SW | SWM | |
| pH | 7.63 ± 0.01 a | 7.34 ± 0.03 d | 7.48 ± 0.02 c | 7.56 ± 0.02 b |
| salt (g/kg) | 8.29 ± 0.07 b | 8.32 ± 0.03 b | 8.95 ± 0.24 a | 7.74 ± 0.08 c |
| EC (mS/cm) | 2.15 ± 0.04 b | 2.22 ± 0.03 b | 2.59 ± 0.06 a | 2.03 ± 0.03 c |
| TN (g/kg) | 0.27 ± 0.00 d | 0.37 ± 0.00 b | 0.39 ± 0.00 c | 0.40 ± 0.00 a |
| AN (mg/kg) | 11.53 ± 0.30 a | 8.38 ± 0.13 c | 11.13 ±0.29 b | 11.67 ± 0.17 ab |
| AP (mg/kg) | 10.69 ± 0.20 bc | 9.85 ± 0.10 c | 11.21 ± 0.26 b | 16.20 ± 0.54 a |
| AK (mg/kg) | 122.88 ±1.15 c | 195.00 ± 2.84 a | 190.09 ± 2.83 ab | 187.06 ± 0.57 b |
| SOC (g/kg) | 1.21 ± 0.00 d | 2.36 ± 0.02 c | 2.37 ±0.03 b | 2.94 ± 0.01 a |
| Alpha Diversity | Treatment | |||
|---|---|---|---|---|
| OS | FW | SW | SWM | |
| No. of sequences | 34,671 | 39,938 | 34,600 | 34,587 |
| ASVs | 978 | 1266 | 862 | 118 |
| Observed_species | 976.00 | 1244.00 | 955.00 | 1160.00 |
| Chao1 | 976.00 | 1244.08 | 955.00 | 1160.00 |
| ACE | 976.00 | 1244.08 | 955.19 | 1160.00 |
| Shannon | 6.16 | 6.50 | 6.12 | 6.29 |
| Simpson | 0.9960 | 0.9969 | 0.9958 | 0.9958 |
| Pielou_J | 0.8952 | 0.9115 | 0.8918 | 0.8919 |
| Pd_faith | 61.74 | 70.29 | 59.83 | 74.93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Q.; Zhang, Y.; Xu, Z.; Qu, W.; Li, J.; Li, W.; Zhao, C.; Ling, H. Irrigation of Suaeda salsa with Saline Wastewater and Microalgae: Improving Saline–Alkali Soil and Revealing the Composition and Function of Rhizosphere Bacteria. Microorganisms 2025, 13, 1653. https://doi.org/10.3390/microorganisms13071653
Yan Q, Zhang Y, Xu Z, Qu W, Li J, Li W, Zhao C, Ling H. Irrigation of Suaeda salsa with Saline Wastewater and Microalgae: Improving Saline–Alkali Soil and Revealing the Composition and Function of Rhizosphere Bacteria. Microorganisms. 2025; 13(7):1653. https://doi.org/10.3390/microorganisms13071653
Chicago/Turabian StyleYan, Qiaoyun, Yitong Zhang, Zhenting Xu, Wenying Qu, Junfeng Li, Wenhao Li, Chun Zhao, and Hongbo Ling. 2025. "Irrigation of Suaeda salsa with Saline Wastewater and Microalgae: Improving Saline–Alkali Soil and Revealing the Composition and Function of Rhizosphere Bacteria" Microorganisms 13, no. 7: 1653. https://doi.org/10.3390/microorganisms13071653
APA StyleYan, Q., Zhang, Y., Xu, Z., Qu, W., Li, J., Li, W., Zhao, C., & Ling, H. (2025). Irrigation of Suaeda salsa with Saline Wastewater and Microalgae: Improving Saline–Alkali Soil and Revealing the Composition and Function of Rhizosphere Bacteria. Microorganisms, 13(7), 1653. https://doi.org/10.3390/microorganisms13071653