
Metagenomic Analysis Revealed the Changes in Antibiotic Resistance Genes and Heavy Metal Resistance Genes in Phosphate Tailings Compost

 ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Composting and Sampling
2.2. Determination of Physicochemical Indexes
2.3. Metagenome Analysis
2.4. Statistical Analysis
3. Results and Discussion
3.1. Changes in Physicochemical Properties of Compost
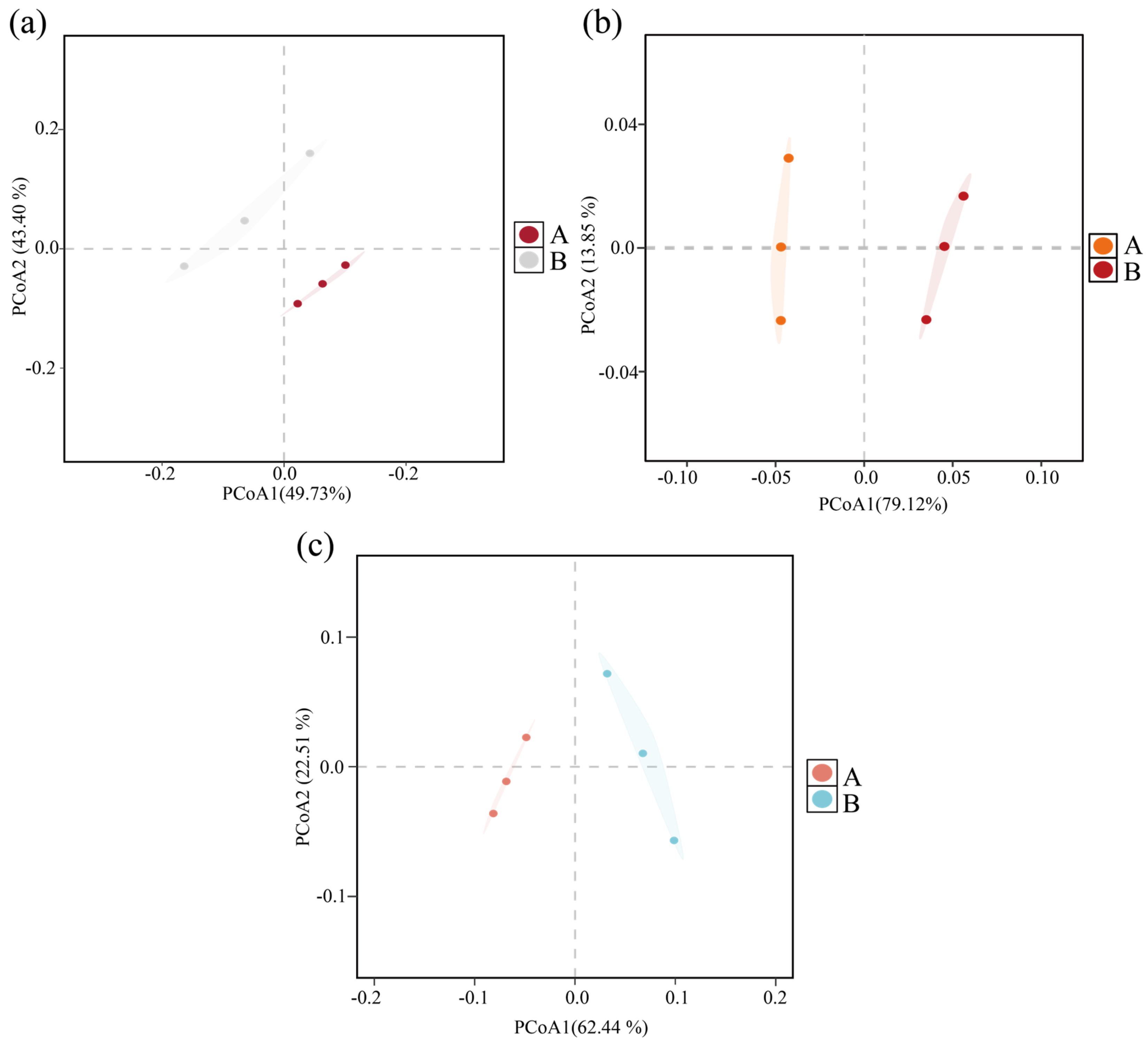
3.2. Microbial Diversity and Community Succession
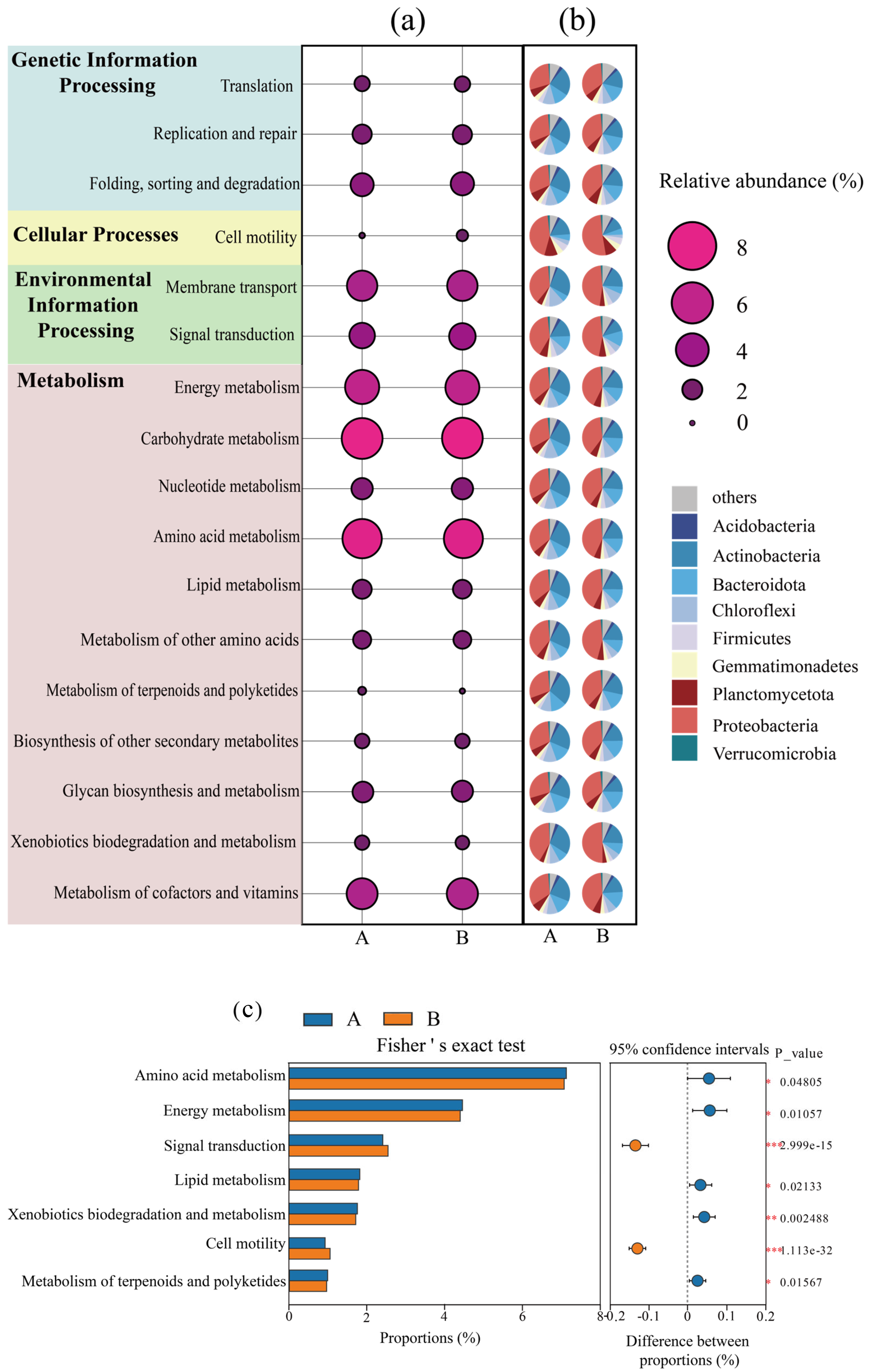
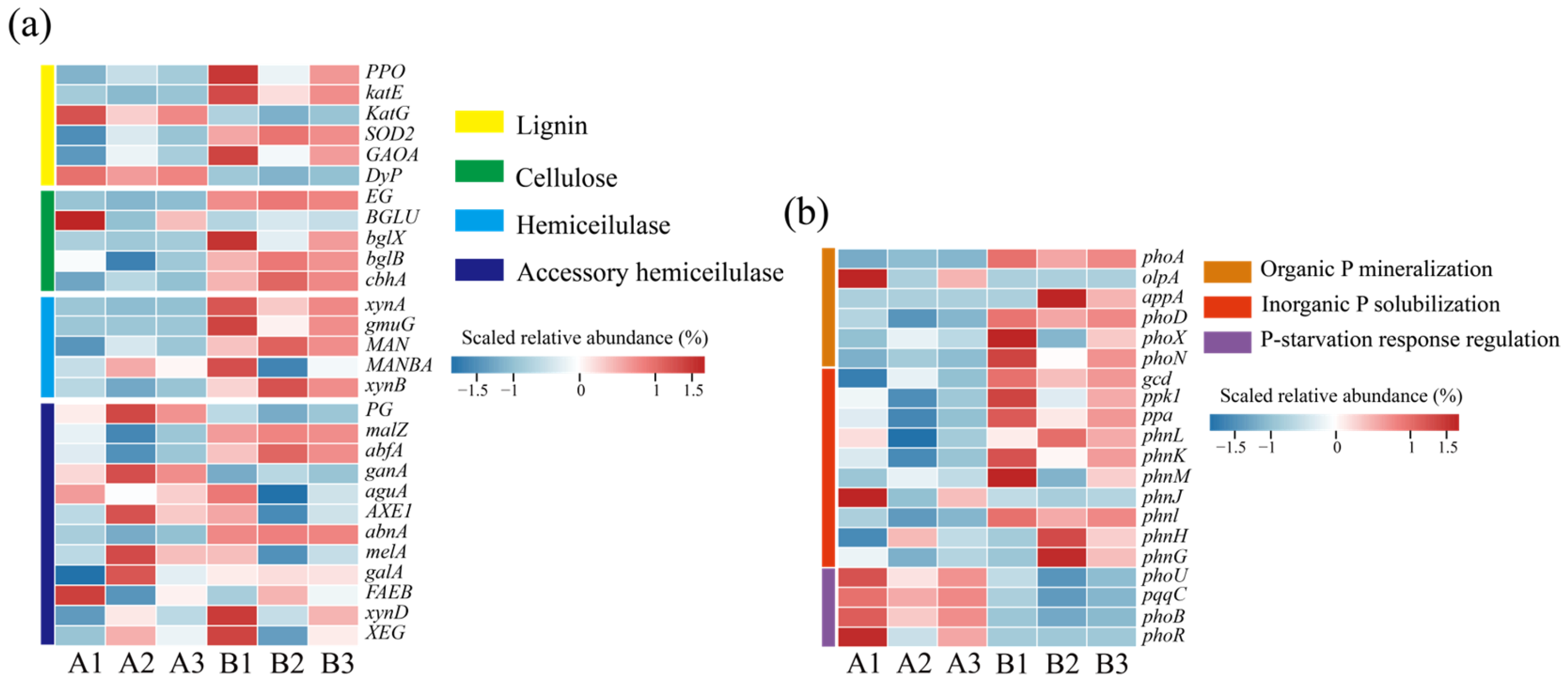
3.3. Metabolic Function Analysis
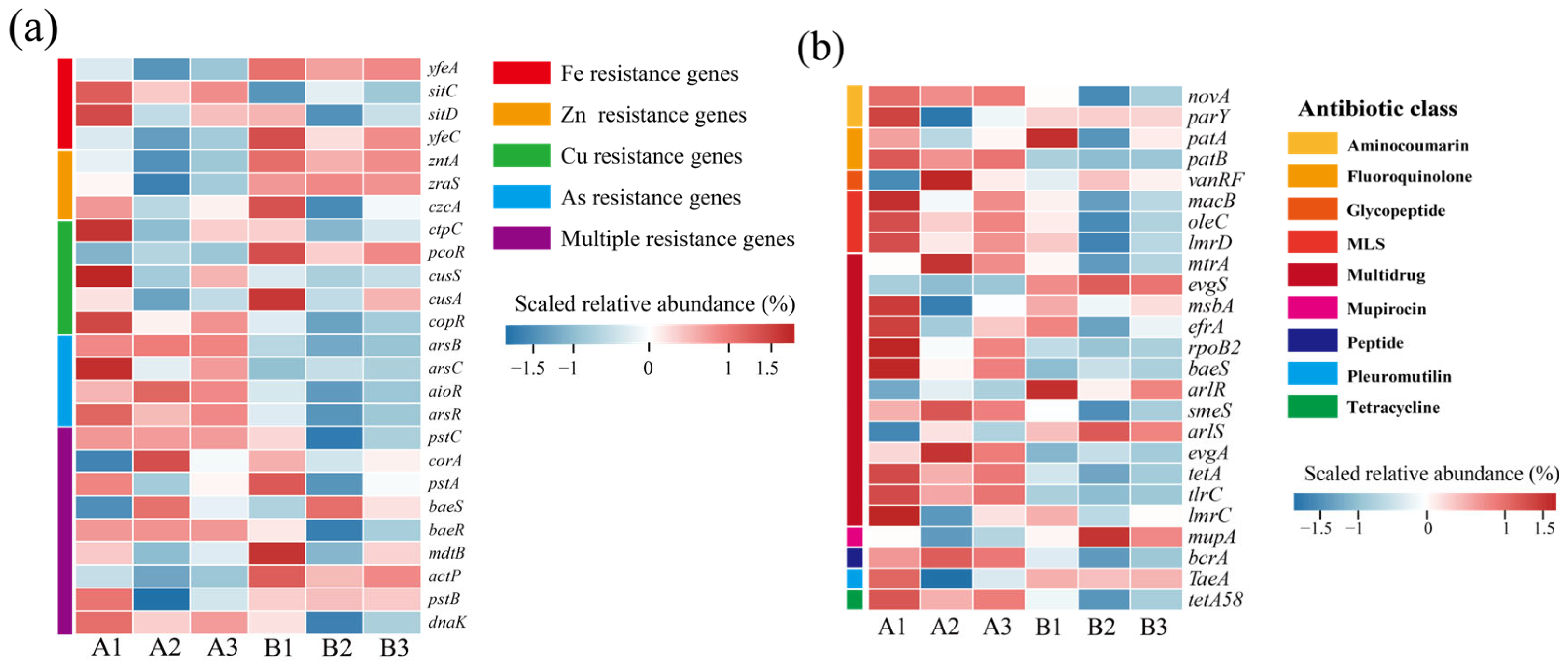
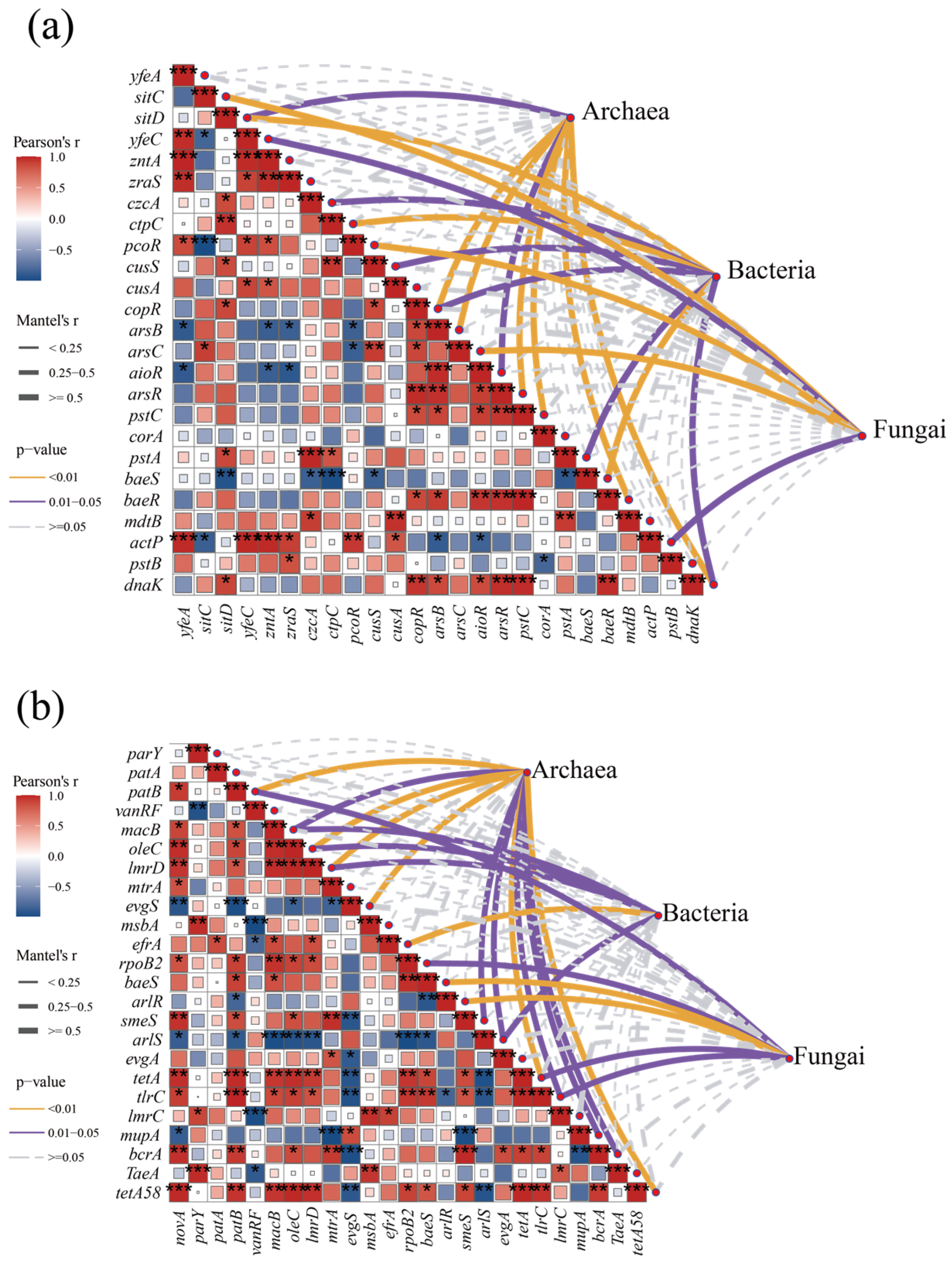
3.4. Changes in Resistance Genes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- García-Berumen, J.A.; Flores de la Torre, J.A.; de los Santos-Villalobos, S.; Espinoza-Canales, A.; Echavarría-Cháirez, F.G.; Gutiérrez-Bañuelos, H. Phosphorus dynamics and sustainable agriculture: The role of microbial solubilization and innovations in nutrient management. Curr. Res. Microb. Sci. 2025, 8, 100326. [Google Scholar] [CrossRef] [PubMed]
- Rana, M.S.; Hu, C.X.; Shaaban, M.; Imran, M.; Afzal, J.; Moussa, M.G.; Elyamine, A.M.; Bhantana, P.; Saleem, M.H.; Syaifudin, M.; et al. Soil phosphorus transformation characteristics in response to molybdenum supply in leguminous crops. J. Environ. Manag. 2020, 268, 110610. [Google Scholar]
- Li, Y.; Guo, S.; Zheng, Y.; Yu, J.; Chi, R.; Xiao, C. Bioimmobilization of lead in phosphate mining wasteland by isolated strain Citrobacter farmeri CFI-01. Environ. Pollut. 2022, 307, 119485. [Google Scholar] [PubMed]
- U.S. Geological Survey. Mineral Commodity Summaries 2022; U.S. Geological Survey: Reston, VA, USA, 2022; p. 202.
- Lustosa Filho, J.F.; Viana, R.d.S.R.; Melo, L.C.A.; de Figueiredo, C.C. Changes in phosphorus due to pyrolysis and in the soil-plant system amended with sewage sludge biochar compared to conventional P fertilizers: A global meta-analysis. Chemosphere 2025, 371, 144055. [Google Scholar]
- Lamghari, K.; Taha, Y.; Ait-Khouia, Y.; Elghali, A.; Hakkou, R.; Benzaazoua, M. Sustainable phosphate mining: Enhancing efficiency in mining and pre-beneficiation processes. J. Environ. Manag. 2024, 358, 120833. [Google Scholar]
- Dai, L.; Li, J.; Zhang, J.; Li, X.; Liu, T.; Yu, Q.; Tao, S.; Zhou, M.; Hou, H. Development and mechanistic study of phosphate tailings based soil heavy metal prophylactic agents with encapsulated structure for lead stabilization and phosphorus speciation in soils. J. Environ. Manag. 2025, 373, 123578. [Google Scholar]
- Yu, Y.-H.; Du, C.-M.; Zhang, Y.-T.; Yuan, R.-Y. Phosphorus recovery from phosphate tailings through a two-stage leaching-precipitation process: Toward the harmless and reduction treatment of P-bearing wastes. Environ. Res. 2024, 248, 118328. [Google Scholar] [CrossRef]
- Pan, C.; Zhang, G.; Yang, Y.; Chen, X.; Wu, J.; Wang, L.; Wei, Z.; Kang, K. Identifying the role of fired clay minerals on reducing of nitrogen loss and immobilization of organic nitrogen during chicken manure composting. Bioresour. Technol. 2022, 349, 126839. [Google Scholar]
- Zhang, S.; Song, C.; Wang, L.; Wang, M.; Zhang, D.; Tang, G. Exploring the promoting effect of nitrilotriacetic acid on hydroxyl radical and humification during magnetite-amended composting of sewage sludge. Bioresour. Technol. 2024, 403, 130863. [Google Scholar]
- Yin, Y.; Yang, C.; Li, M.; Zheng, Y.; Ge, C.; Gu, J.; Li, H.; Duan, M.; Wang, X.; Chen, R. Research progress and prospects for using biochar to mitigate greenhouse gas emissions during composting: A review. Sci. Total Environ. 2021, 798, 149294. [Google Scholar]
- Aycan DÜMencİ, N.; Cagcag Yolcu, O.; Aydin Temel, F.; Turan, N.G. Identifying the maturity of co-compost of olive mill waste and natural mineral materials: Modelling via ANN and multi-objective optimization. Bioresour. Technol. 2021, 338, 125516. [Google Scholar]
- Qiu, X.; Feng, M.; Zhou, G.; Wang, H. Effects of mineral additives on antibiotic resistance genes and related mechanisms during chicken manure composting. Bioresour. Technol. 2022, 346, 126631. [Google Scholar]
- Ravindran, B.; Karmegam, N.; Awasthi, M.K.; Chang, S.W.; Selvi, P.K.; Balachandar, R.; Chinnappan, S.; Azelee, N.I.W.; Munuswamy-Ramanujam, G. Valorization of food waste and poultry manure through co-composting amending saw dust, biochar and mineral salts for value-added compost production. Bioresour. Technol. 2022, 346, 126442. [Google Scholar]
- Zhang, J.; Dai, L.; Li, J.; Zeng, Q.; Zhou, M.; Hou, H. Phosphate tailings-based slow-release heavy metal passivation materials: Mechanism, environmental risk, and microbial community evolution. J. Clean. Prod. 2024, 434, 126442. [Google Scholar]
- Zhou, Z.; Keiblinger, K.M.; Huang, Y.; Bhople, P.; Shi, X.; Yang, S.; Yu, F.; Liu, D. Virome and metagenomic sequencing reveal the impact of microbial inoculants on suppressions of antibiotic resistome and viruses during co-composting. J. Hazard. Mater. 2024, 477, 135355. [Google Scholar]
- Xiao, G.; Li, T.; Zhang, X.; Yu, H.; Huang, H.; Gupta, D.K. Uptake and accumulation of phosphorus by dominant plant species growing in a phosphorus mining area. J. Hazard. Mater. 2009, 171, 542–550. [Google Scholar]
- Sharifi, M.; Zebarth, B.J.; Burton, D.L.; Grant, C.A.; Hajabbasi, M.A.; Abbassi-Kalo, G. Sodium Hydroxide Direct Distillation: A Method for Estimating Total Nitrogen in Soil. Commun. Soil Sci. Plant Anal. 2009, 40, 2505–2520. [Google Scholar] [CrossRef]
- Meng, L.; Xu, C.; Wu, F.; Huhe. Microbial co-occurrence networks driven by low-abundance microbial taxa during composting dominate lignocellulose degradation. Sci. Total Environ. 2022, 845, 157197. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar]
- Zhang, D.; Li, X.; Li, H.; Xu, Y. Microbial inoculants enhance the persistence of antibiotic resistance genes in aerobic compost of food waste mainly by altering interspecific relationships. Bioresour. Technol. 2023, 385, 129443. [Google Scholar]
- Hong, L.; Hu, Z.; Sun, S.; Tang, X.; Wang, J.; Tan, Q.; Zheng, L.; Wang, S.; Xu, S.; King, I.; et al. Fast, sensitive detection of protein homologs using deep dense retrieval. Nat. Biotechnol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Hoang, H.G.; Thuy, B.T.P.; Lin, C.; Vo, D.-V.N.; Tran, H.T.; Bahari, M.B.; Le, V.G.; Vu, C.T. The nitrogen cycle and mitigation strategies for nitrogen loss during organic waste composting: A review. Chemosphere 2022, 300, 134514. [Google Scholar] [PubMed]
- Mo, J.; Xin, L.; Zhao, C.; Qin, Y.; Nan, Q.; Mei, Q.; Wu, W. Reducing nitrogen loss during kitchen waste composting using a bioaugmented mechanical process with low pH and enhanced ammonia assimilation. Bioresour. Technol. 2023, 372, 128664. [Google Scholar]
- Liu, J.; Bao, Z.; Wang, C.; Wei, J.; Wei, Y.; Chen, M. Understanding of mercury and methylmercury transformation in sludge composting by metagenomic analysis. Water Res. 2022, 226, 119204. [Google Scholar]
- Shen, C.; Shangguan, H.; Fu, T.; Mi, H.; Lin, H.; Huang, L.; Tang, J. Electric field-assisted aerobic co-composting of chicken manure and kitchen waste: Ammonia mitigation and maturation enhancement. Bioresour. Technol. 2024, 391, 129931. [Google Scholar] [CrossRef]
- Kitamura, R.; Kozaki, T.; Ishii, K.; Iigo, M.; Kurokura, T.; Yamane, K.; Maeda, I.; Iwabuchi, K.; Saito, T. Utilizing Cattle Manure Compost Increases Ammonia Monooxygenase A Gene Expression and Ammonia-oxidizing Activity of Both Bacteria and Archaea in Biofiltration Media for Ammonia Deodorization. Microbes Environ. 2021, 36, ME20148. [Google Scholar]
- Yang, H.; Huang, Y.; Li, K.; Zhu, P.; Wang, Y.; Li, X.; Meng, Q.; Niu, Q.; Wang, S.; Li, Q. Lignocellulosic depolymerization induced by ionic liquids regulating composting habitats based on metagenomics analysis. Environ. Sci. Pollut. Res. 2022, 29, 76298–76309. [Google Scholar] [CrossRef]
- Jiang, Z.; Li, X.; Li, M.; Zhu, Q.; Li, G.; Ma, C.; Li, Q.; Meng, J.; Liu, Y.; Li, Q. Impacts of red mud on lignin depolymerization and humic substance formation mediated by laccase-producing bacterial community during composting. J. Hazard. Mater. 2021, 410, 124557. [Google Scholar]
- Neher, D.A.; Limoges, M.A.; Weicht, T.R.; Sharma, M.; Millner, P.D.; Donnelly, C. Bacterial Community Dynamics Distinguish Poultry Compost from Dairy Compost and Non-Amended Soils Planted with Spinach. Microorganisms 2020, 8, 1601. [Google Scholar] [CrossRef]
- Zhong, B.; An, X.; An, W.; Xiao, X.; Li, H.; Xia, X.; Zhang, Q. Effect of bioaugmentation on lignocellulose degradation and antibiotic resistance genes removal during biogas residues composting. Bioresour. Technol. 2021, 340, 125742. [Google Scholar] [CrossRef]
- Zheng, Y.; Feng, Z.; Wang, P.; Xu, S.; Gao, X.; Ren, L.; Yang, T.; Zhao, X.; Wei, Y.; Li, J. Suppressive performance of food waste composting with polylactic acid: Emphasis on microbial core metabolism pathways and mechanism. Bioresour. Technol. 2023, 384, 129339. [Google Scholar]
- Duan, M.; Zhang, Y.; Zhou, B.; Qin, Z.; Wu, J.; Wang, Q.; Yin, Y. Effects of Bacillus subtilis on carbon components and microbial functional metabolism during cow manure–straw composting. Bioresour. Technol. 2020, 303, 122868. [Google Scholar]
- Zheng, Y.; Cao, X.; Zhou, Y.; Ma, S.; Wang, Y.; Li, Z.; Zhao, D.; Yang, Y.; Zhang, H.; Meng, C.; et al. Purines enrich root-associated Pseudomonas and improve wild soybean growth under salt stress. Nat. Commun. 2024, 15, 1–14. [Google Scholar]
- Qu, F.; Cheng, H.; Han, Z.; Wei, Z.; Song, C. Identification of driving factors of lignocellulose degrading enzyme genes in different microbial communities during rice straw composting. Bioresour. Technol. 2023, 381, 129109. [Google Scholar]
- Tian, X.; Qin, X.; Jia, X.; Lyu, Q.; Li, S.; Jiang, L.; Chen, L.; Yan, Z.; Huang, J. Lignocellulose degradation and temperature adaptation mechanisms during composting of mushroom residue and wood chips at low temperature with inoculation of psychrotolerant microbial agent. Environ. Pollut. 2024, 363, 125156. [Google Scholar] [CrossRef]
- Siles, J.A.; Starke, R.; Martinovic, T.; Parente Fernandes, M.L.; Orgiazzi, A.; Bastida, F. Distribution of phosphorus cycling genes across land uses and microbial taxonomic groups based on metagenome and genome mining. Soil Biol. Biochem. 2022, 174, 108826. [Google Scholar]
- Dai, Z.; Liu, G.; Chen, H.; Chen, C.; Wang, J.; Ai, S.; Wei, D.; Li, D.; Ma, B.; Tang, C.; et al. Long-term nutrient inputs shift soil microbial functional profiles of phosphorus cycling in diverse agroecosystems. ISME J. 2020, 14, 757–770. [Google Scholar] [CrossRef]
- Liu, X.; Han, R.; Cao, Y.; Turner, B.L.; Ma, L.Q. Enhancing Phytate Availability in Soils and Phytate-P Acquisition by Plants: A Review. Environ. Sci. Technol. 2022, 56, 9196–9219. [Google Scholar] [CrossRef]
- Guo, H.; Gu, J.; Wang, X.; Yu, J.; Nasir, M.; Peng, H.; Zhang, R.; Hu, T.; Wang, Q.; Ma, J. Responses of antibiotic and heavy metal resistance genes to bamboo charcoal and bamboo vinegar during aerobic composting. Environ. Pollut. 2019, 252, 1097–1105. [Google Scholar]
- Sun, Q.; Zhang, Q.; Li, H.; Ming, C.; Gao, J.; Li, Y.; Zhang, Y. Regulatory effects of different anionic surfactants on the transformation of heavy metal fractions and reduction of heavy metal resistance genes in chicken manure compost. Environ. Pollut. 2023, 335, 122297. [Google Scholar]
- Jiang, X.; Liu, W.; Xu, H.; Cui, X.; Li, J.; Chen, J.; Zheng, B. Characterizations of heavy metal contamination, microbial community, and resistance genes in a tailing of the largest copper mine in China. Environ. Pollut. 2021, 280, 116947. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhu, L.; Chen, S.; Wu, H.; Li, R.; Li, J.; Yuan, J.; Wen, T.; Xue, C.; Shen, Q. Risk assessment and dissemination mechanism of antibiotic resistance genes in compost. Environ. Int. 2023, 178, 108126. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Xie, S.; Zeng, M.; Xu, X.; Li, Y.; Liu, X.; Wang, X. Impacts of pile temperature on antibiotic resistance, metal resistance and microbial community during swine manure composting. Sci. Total Environ. 2020, 744, 140920. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Cao, X.; Chen, S.; Ao, X.; Li, J.; Hu, K.; Liu, S.; Penttinen, P.; Yang, Y.; Yu, X.; et al. Antibiotic and heavy metal resistance genes in sewage sludge survive during aerobic composting. Sci. Total Environ. 2023, 866, 161386. [Google Scholar] [CrossRef]
- Liang, Y.; Yan, Y.; Shi, L.; Wang, M.; Yuan, X.; Wang, S.; Ye, L.; Yan, Z. Molecular Basis of Thioredoxin-Dependent Arsenic Transformation in Methanogenic Archaea. Environ. Sci. Technol. 2024, 59, 443–453. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | A | B |
|---|---|---|
| TN (g/kg) | 1.75 ± 0.12 a | 0.98 ± 0.23 b |
| Ws-P (mg/kg) | 3.24 ± 0.14 b | 7.21 ± 0.09 a |
| AK (mg/kg) | 0.56 ± 0.21 b | 1.90 ± 0.11 a |
| pH | 7.32 ± 0.15 b | 8.68 ± 0.21 a |
| Bacteria | Fungi | Archaea | ||||
|---|---|---|---|---|---|---|
| A | B | A | B | A | B | |
| Shannon | 5.43 ± 0.05 | 5.48 ± 0.01 | 5.59 ± 0.01 | 5.45 ± 0.09 | 3.49 ± 0.09 | 3.42 ± 0.29 |
| ACE | 3906.66 ± 23.31 | 3827 ± 34.36 | 985.21 ± 14.61 | 960.43 ± 33.99 | 163.48 ± 2.59 | 164.63 ± 5.10 |
| Good’s coverage | 0.998 ± 0.001 | 0.997 ± 0.001 | 0.944 ± 0.004 | 0.945 ± 0.001 | 0.989 ± 0.001 | 0.984 ± 0.003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, C.; Wan, K.; Chen, Y.; Jin, Y.; Zhou, F.; Yu, J.; Chi, R. Metagenomic Analysis Revealed the Changes in Antibiotic Resistance Genes and Heavy Metal Resistance Genes in Phosphate Tailings Compost. Microorganisms 2025, 13, 768. https://doi.org/10.3390/microorganisms13040768
Xiao C, Wan K, Chen Y, Jin Y, Zhou F, Yu J, Chi R. Metagenomic Analysis Revealed the Changes in Antibiotic Resistance Genes and Heavy Metal Resistance Genes in Phosphate Tailings Compost. Microorganisms. 2025; 13(4):768. https://doi.org/10.3390/microorganisms13040768
Chicago/Turabian StyleXiao, Chunqiao, Kai Wan, Yan Chen, Yongtong Jin, Fang Zhou, Junxia Yu, and Ruan Chi. 2025. "Metagenomic Analysis Revealed the Changes in Antibiotic Resistance Genes and Heavy Metal Resistance Genes in Phosphate Tailings Compost" Microorganisms 13, no. 4: 768. https://doi.org/10.3390/microorganisms13040768
APA StyleXiao, C., Wan, K., Chen, Y., Jin, Y., Zhou, F., Yu, J., & Chi, R. (2025). Metagenomic Analysis Revealed the Changes in Antibiotic Resistance Genes and Heavy Metal Resistance Genes in Phosphate Tailings Compost. Microorganisms, 13(4), 768. https://doi.org/10.3390/microorganisms13040768