
Lankesterella and Isospora Coccidians: Differences in Host Specificity of Blood Parasites in Passerines


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methodology
2.1. Field Work and Sampling
2.2. Parasite Detection Methods
2.3. Haplotype Network Analysis
2.4. Phylogenetic Analysis
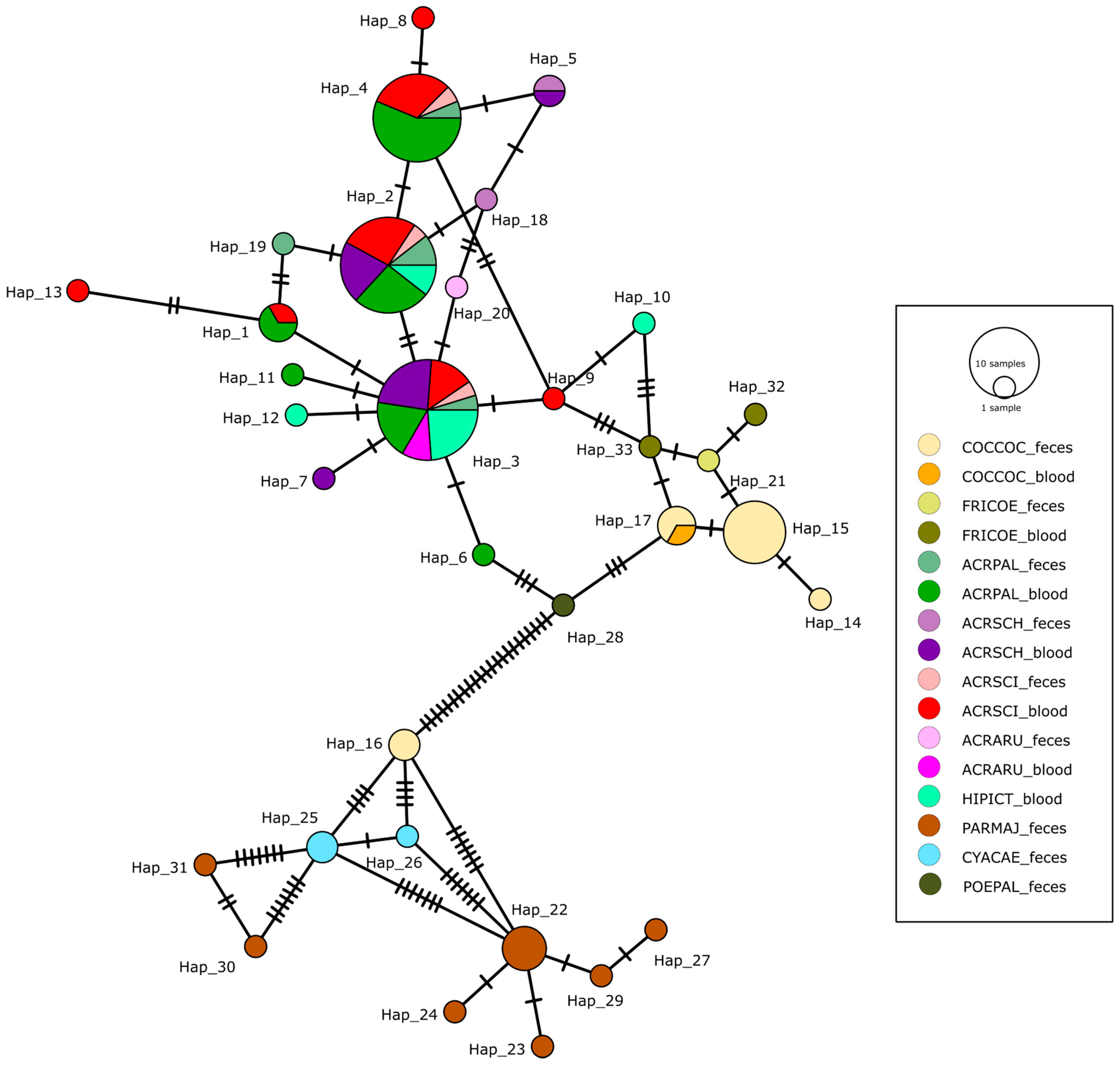
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Box, E.D. Influence of Isospora infections on patency of avian Lankesterella (Atoxoplasma, Garnham 1950). J. Parasitol. 1967, 53, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Keckeisen, C.; Šujanová, A.; Himmel, T.; Matt, J.; Nedorost, N.; Chagas, C.R.; Weissenböck, H.; Harl, J. Isospora and Lankesterella Parasites (Eimeriidae, Apicomplexa) of Passeriform Birds in Europe: Infection Rates, Phylogeny, and Pathogenicity. Pathogens 2024, 13, 337. [Google Scholar] [CrossRef] [PubMed]
- Chagas, C.R.F.; Harl, J.; Preikša, V.; Bukauskaitė, D.; Ilgūnas, M.; Weissenböck, H.; Valkiūnas, G. Lankesterella (Apicomplexa, Lankesterellidae) blood parasites of passeriform birds: Prevalence, molecular and morphological characterization, with notes on sporozoite persistence in vivo and development in vitro. Animals 2021, 11, 1451. [Google Scholar] [CrossRef]
- Merino, S.; Martínez, J.; Martínez-de la Puente, J.; Criado-Fornelio, Á.; Tomás, G.; Morales, J.; Lobato, E.; García-Fraile, S. Molecular characterization of the 18S rDNA gene of an avian Hepatozoon reveals that it is closely related to Lankesterella. J. Parasitol. 2006, 92, 1330–1335. [Google Scholar] [CrossRef]
- Biedrzycka, A.; Kloch, A.; Migalska, M.; Bielański, W. Molecular characterization of putative Hepatozoon sp. from the sedge warbler (Acrocephalus schoenobaenus). Parasitology 2013, 140, 695–698. [Google Scholar] [CrossRef]
- Martínez González, F.J.; Merino, S.; Pérez Badás, E.; Almazán, L.; Moksnes, A.; Barbosa, A. Hemoparasites and immunological parameters in Snow Bunting (Plectrophenax nivalis) nestlings. Polar Biol. 2018, 41, 1855–1866. [Google Scholar] [CrossRef]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eukaryot. Microbiol. 2018, 66, 4–119. [Google Scholar] [CrossRef]
- Labbé, A. Sporozoa. Das Tierreich. Deutschen Zoologischen Gesellschaft. 5. Lieferung; R. Friedländer und Sohn Verlag: Berlin, Germany, 1899; 180p. [Google Scholar]
- Alvarez Calvo, J.A. New species of haemococcidia in Spanish Lacertidae. Cuad. Cienc. Biol. 1975, 4, 207–222. [Google Scholar]
- Venkatachalam, A.K.S.B.; Kadlecová, A.; Kapustová, A.; Kulich Fialová, M.; Brzoňová, J.; Šálek, M.; Svobodová, M. Factors influencing a common but neglected blood parasite prevalence in breeding populations of passerines. Parasitology 2025, 1–24. [Google Scholar] [CrossRef]
- Venkatachalam, A.K.S.B.; Čepička, I.; Hrazdilová, K.; Svobodová, M. Host specificity of passerine Lankesterella (Apicomplexa: Coccidia). Eur. J. Protistol. 2023, 90, 126007. [Google Scholar] [CrossRef]
- Bennett, G.F.; Peirce, M.A. Hepatozoon parus n. sp. from the Paridae and redescription of H. atticorae (de Beaurepaire Aragão 1911) Hoare 1924 from the Hirundinidae. Can. J. Zool. 1989, 67, 2859–2863. [Google Scholar]
- Megía-Palma, R.; Martínez, J.; Nasri, I.; Cuervo, J.J.; Martín, J.; Acevedo, I.; Belliure, J.; Ortega, J.; García-Roa, R.; Selmi, S.; et al. Phylogenetic relationships of Isospora, Lankesterella, and Caryospora species (Apicomplexa: Eimeriidae) infecting lizards. Org. Divers. Evol. 2016, 16, 275–288. [Google Scholar] [CrossRef]
- Knight, A.; Ewen, J.G.; Brekke, P.; Santure, A.W. The evolutionary biology, ecology and epidemiology of coccidia of passerine birds. Adv. Parasitol. 2018, 99, 35–60. [Google Scholar] [CrossRef]
- Kubiski, S.V.; Witte, C.; Burchell, J.A.; Conradson, D.; Zmuda, A.; Barbon, A.R.; Vilches-Moure, J.G.; Felt, S.A.; Rideout, B. A Mitochondrial gene diversity and host specificity of Isospora in passerine birds. Front. Vet. Sci. 2022, 9, 847030. [Google Scholar] [CrossRef]
- Box, E.D. Isospora as an Extraintestinal Parasite of Passerine Birds 1. J. Protozool. 1981, 28, 244–246. [Google Scholar]
- Barta, J.R.; Schrenzel, M.D.; Carreno, R.; Rideout, B.A. The genus Atoxoplasma (Garnham 1950) as a junior objective synonym of the genus Isospora (Schneider 1881) species infecting birds and resurrection of Cystoisospora (Frenkel 1977) as the correct genus for Isospora species infecting mammals. J. Parasitol. 2005, 91, 726–727. [Google Scholar] [CrossRef]
- Schrenzel, M.D.; Maalouf, G.A.; Gaffney, P.M.; Tokarz, D.; Keener, L.L.; McClure, D.; Griffey, D.S.; Bruce, M.A.; Rideout, B. A Molecular characterization of isosporoid coccidia (Isospora and Atoxoplasma spp.) in passerine birds. J. Parasitol. 2005, 91, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Box, E.D. Life cycles of two Isospora species in the canary, Serinus canarius Linnaeus. J. Protozool. 1977, 24, 57–67. [Google Scholar] [CrossRef]
- Trefancová, A.; Kvičerová, J. Isospora svecica sp. n. (Apicomplexa: Eimeriidae), a new species of coccidium from the white-spotted bluethroat Luscinia svecica cyanecula (Aves: Passeriformes: Muscicapidae). Parasitol. Res. 2019, 118, 3043–3051. [Google Scholar] [CrossRef]
- Dolnik, O.V. Isospora sylvianthina (Protozoa: Coccidiida), parasite of Blackcap, does not infect Reed Warbler. Zoosyst. Ross. 2002, 10, 240. [Google Scholar]
- Levine, N.D. Isospora passeris n. sp. from the house sparrow Passer domesticus, I. lacazei, and related apicomplexan Protozoa. Trans. Am. Microsc. Soc. 1982, 101, 66–74. [Google Scholar] [CrossRef]
- Storchová, L.; Hořák, D. Life-history characteristics of European birds. Glob. Ecol. Biogeogr. 2018, 27, 400–406. [Google Scholar] [CrossRef]
- Fialová, M.; Santolíková, A.; Brotánková, A.; Brzoňová, J.; Svobodová, M. Complete life cycle of Trypanosoma thomasbancrofti, an avian trypanosome transmitted by culicine mosquitoes. Microorganisms 2021, 9, 2101. [Google Scholar] [CrossRef] [PubMed]
- Kvičerová, J.; Pakandl, M.; Hypša, V. Phylogenetic relationships among Eimeria spp. (Apicomplexa, Eimeriidae) infecting rabbits: Evolutionary significance of biological and morphological features. Parasitology 2008, 135, 443–452. [Google Scholar] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D.; Nakagawa, S. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Schumm, Y.R.; Wecker, C.; Marek, C.; Wassmuth, M.; Bentele, A.; Willems, H.; Reiner, G.; Quillfeldt, P. Blood parasites in Passeriformes in central Germany: Prevalence and lineage diversity of Haemosporida (Haemoproteus, Plasmodium and Leucocytozoon) in six common songbirds. PeerJ 2019, 6, e6259. [Google Scholar] [CrossRef]
- Wong, M.L.; Ahmed, M.A.; Sulaiman WY, W.; Manin, B.O.; Leong, C.S.; Quan, F.S.; Vythilingam, I. Genetic diversity of zoonotic malaria parasites from mosquito vector and vertebrate hosts. Infect. Genet. Evol. 2019, 73, 26–32. [Google Scholar] [CrossRef]
- Doussang, D.; Sallaberry-Pincheira, N.; Cabanne, G.S.; Lijtmaer, D.A.; González-Acuña, D.; Vianna, J.A. Specialist versus generalist parasites: The interactions between host diversity, environment and geographic barriers in avian malaria. Int. J. Parasitol. 2021, 51, 899–911. [Google Scholar] [CrossRef]
- Dolnik, O.V.; Palinauskas, V.; Bensch, S. Individual oocysts of Isospora (Apicomplexa: Coccidia) parasites from avian feces: From photo to sequence. J. Parasitol. 2009, 95, 169–174. [Google Scholar] [CrossRef]
- Svobodová, M. Isospora, Caryospora and Eimeria (Apicomplexa: Eimeriidae) in Passeriform Birds from Czech Republic. Acta Protozool. 1994, 33, 101–108. [Google Scholar]
- Lopez, G.; Figuerola, J.; Soriguer, R. Time of day, age and feeding habits influence coccidian oocyst shedding in wild passerines. Int. J. Parasitol. 2007, 37, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Dolnik, O.V.; Dolnik, V.R.; Bairlein, F. The effect of host foraging ecology on the prevalence and intensity of coccidian infection in wild passerine birds. Ardea 2010, 98, 97–103. [Google Scholar] [CrossRef]
- Svobodová, M.; Cibulkova, M. Isospora sp. (Apicomplexa: Eimeriidae) in icterine warbler (Hippolais icterina, Passeriformes: Sylviidae): The possibility of parents to nestlings transmission. Acta Protozool. 1995, 34, 233. [Google Scholar]
- Kadlecová, A. Blood Parasites of Warblers (Acrocephalus). Master’s Thesis, Charles University, Prague, Czech Republic, 18 September 2024. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saravana Bhavan Venkatachalam, A.K.; Brzoňová, J.; Svobodová, M. Lankesterella and Isospora Coccidians: Differences in Host Specificity of Blood Parasites in Passerines. Microorganisms 2025, 13, 743. https://doi.org/10.3390/microorganisms13040743
Saravana Bhavan Venkatachalam AK, Brzoňová J, Svobodová M. Lankesterella and Isospora Coccidians: Differences in Host Specificity of Blood Parasites in Passerines. Microorganisms. 2025; 13(4):743. https://doi.org/10.3390/microorganisms13040743
Chicago/Turabian StyleSaravana Bhavan Venkatachalam, Ashwin Kumar, Jana Brzoňová, and Milena Svobodová. 2025. "Lankesterella and Isospora Coccidians: Differences in Host Specificity of Blood Parasites in Passerines" Microorganisms 13, no. 4: 743. https://doi.org/10.3390/microorganisms13040743
APA StyleSaravana Bhavan Venkatachalam, A. K., Brzoňová, J., & Svobodová, M. (2025). Lankesterella and Isospora Coccidians: Differences in Host Specificity of Blood Parasites in Passerines. Microorganisms, 13(4), 743. https://doi.org/10.3390/microorganisms13040743