
Analysis of Microbial Diversity and Community Structure of Rhizosphere Soil of Three Astragalus Species Grown in Special High-Cold Environment of Northwestern Yunnan, China

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Determination of Soil Physicochemical Properties
2.3. Genomic DNA Extraction and PCR Amplification
2.4. Illumina MiSeq Sequencing and Processing
2.5. Bioinformatics and Statistical Analysis
3. Results
3.1. Soil Physicochemical Properties
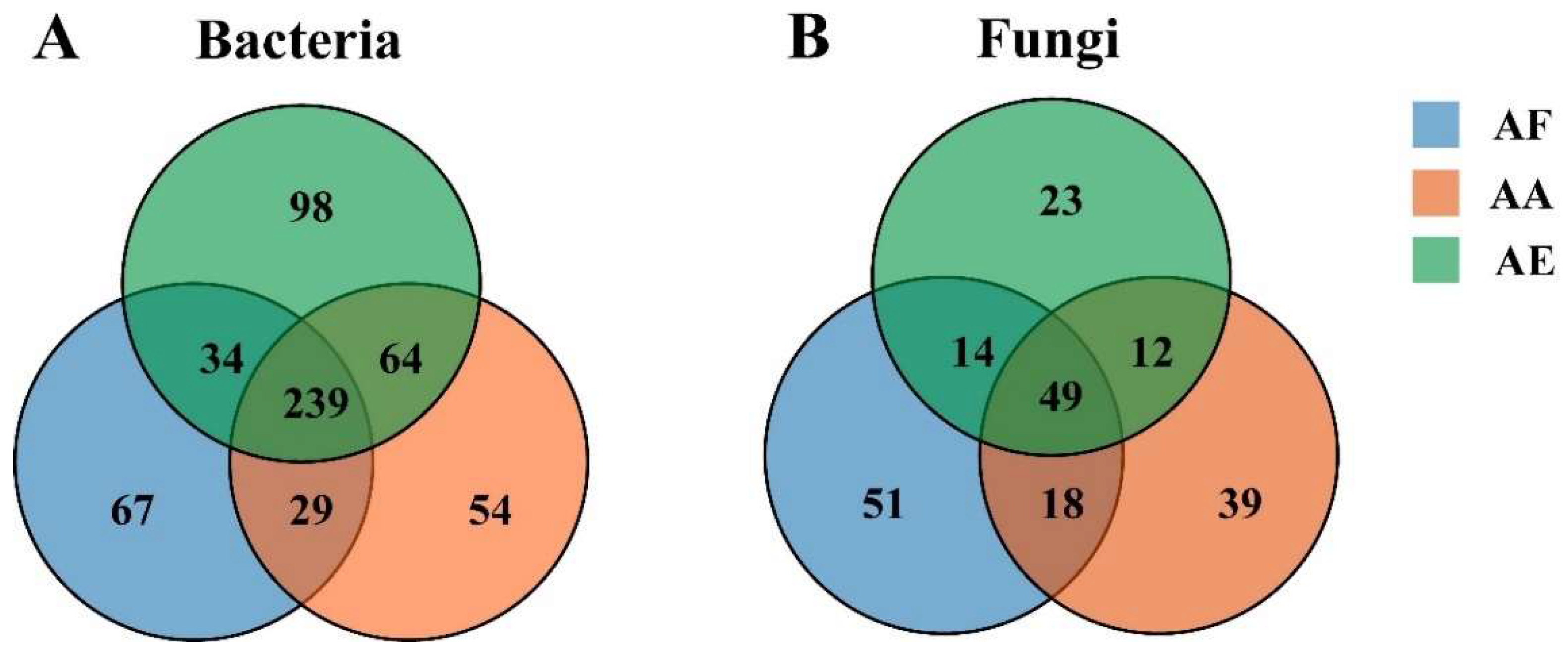
3.2. Differential Microbial Diversity in Astragalus’s Rhizosphere
3.3. Bacterial Community Analysis
3.4. Fungal Community Analysis
3.5. Identification of Microbial Biomarkers
3.6. Relationship between Microbial Communities and Soil Physicochemical Properties
3.7. KEGG Pathway Enrichment Analysis
3.8. Co-Occurrence Network Analysis of Microbial Community
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, M.; Chai, L.; Huang, M.; Jia, W.; Guo, J.; Huang, Y. Deciphering the archaeal communities in tree rhizosphere of the Qinghai-Tibetan plateau. BMC Microbiol. 2020, 20, 235. [Google Scholar] [CrossRef]
- Zuo, J.; Zu, M.; Liu, L.; Song, X.; Yuan, Y. Composition and diversity of bacterial communities in the rhizosphere of the Chinese medicinal herb Dendrobium. BMC Plant Biol. 2021, 21, 127. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Liu, A.; Li, Y.; Wang, Q.; Zhang, X.; Xiong, J.; Li, Y.; Lei, Y.; Sun, Y. Analysis of microbial diversity and community structure of rhizosphere soil of Cistanche Salsa from different host plants. Front. Microbiol. 2022, 13, 971228. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, W.; Shen, Z.; Wang, J.; Han, M. Comparison and interpretation of characteristics of rhizosphere microbiomes of three blueberry varieties. BMC Microbiol. 2021, 21, 30. [Google Scholar] [CrossRef]
- Jiménez, D.J.; Andreote, F.D.; Chaves, D.; Montaña, J.S.; Osorio-Forero, C.; Junca, H.; Zambrano, M.M.; Baena, S. Structural and functional insights from the metagenome of an acidic hot spring microbial planktonic community in the Colombian Andes. PLoS ONE 2012, 7, e52069. [Google Scholar] [CrossRef]
- Zhang, J.; Xing, P.; Niu, M.; Wei, G.; Shi, P. Taxonomic compositions and co-occurrence relationships of protists in bulk soil and rhizosphere of soybean fields in different regions of China. Front. Microbiol. 2021, 12, 738129. [Google Scholar] [CrossRef]
- Chen, C.; Gong, G.-F.; Liang, J.-Y.; Dou, Z.-R.; Ding, J.-J.; Jiang, B.; Wang, K.-L. Rhizosphere microbial diversity of Astragalus yunnanensis and A. tatsienensis var. incanus in Baima Snow Mountain and screening of microorganisms with anti-biofilm activity. Microbiology 2022, 49, 3813–3836. [Google Scholar]
- Li, J.-Q.; Xiao, C.-J.; Li, Y.-M.; Tian, X.-Y.; Dong, X.; Jiang, B. Astrernestin, a novel aurone-phenylpropanoid adduct from the roots of Astragalus ernestii. Nat. Prod. Res. 2020, 34, 19–21. [Google Scholar] [CrossRef]
- Abd Elkader, H.T.; Essawy, A.E.; Al-Shami, A.S. Astragalus species: Phytochemistry, biological actions and molecular mechanisms underlying their potential neuroprotective effects on neurological diseases. Phytochemistry 2022, 202, 113293. [Google Scholar] [CrossRef]
- Lv, C.; Wang, C.; Cai, A.; Zhou, Z. Global magnitude of rhizosphere effects on soil microbial communities and carbon cycling in natural terrestrial ecosystems. Sci. Total Environ. 2023, 856, 158961. [Google Scholar] [CrossRef]
- Chen, J.-S.; Li, J.; Zhang, Y.; Zong, H.; Lei, N.-F. Clonal integration ameliorates the carbon accumulation capacity of a stoloniferous herb, Glechoma longituba, growing in heterogenous light conditions by facilitating nitrogen assimilation in the rhizosphere. Ann. Bot. 2015, 115, 127–136. [Google Scholar] [CrossRef]
- Liu, H.; Cheng, J.; Jin, H.; Xu, Z.; Yang, X.; Min, D.; Xu, X.; Shao, X.; Lu, D.; Qin, B. Characterization of rhizosphere and endophytic microbial communities associated with Stipa purpurea and their correlation with soil environmental factors. Plants 2022, 11, 363. [Google Scholar] [CrossRef]
- He, H.; Ge, R.; Ren, X.; Zhang, L.; Chang, Q.; Xu, Q.; Zhou, G.; Xie, Z.; Wang, S.; Wang, H.; et al. Reference carbon cycle dataset for typical chinese forests via colocated observations and data assimilation. Sci. Data 2021, 8, 42. [Google Scholar] [CrossRef]
- Chen, J.; Wang, P.; Wang, C.; Wang, X.; Miao, L.; Liu, S.; Yang, Q.S. Bacterial communities in riparian sediments: A large-scale longitudinal distribution pattern and response to dam construction. Front. Microbiol. 2018, 9, 999. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Suo, M.; Qiu, Z.; Wu, H.; Zhao, M.; Yang, H. Regulating root fungal community using mortierella alpina for Fusarium oxysporum resistance in Panax ginseng. Front. Microbiol. 2022, 13, 850917. [Google Scholar] [CrossRef]
- Lutz, K.A.; Wang, W.; Zdepski, A.; Michael, T.P. Isolation and analysis of high quality nuclear DNA with reduced organellar DNA for plant genome sequencing and resequencing. BMC Biotechnol. 2011, 11, 1–9. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4516–4522. [Google Scholar] [CrossRef]
- Chen, J.; Huang, X.; Tong, B.; Wang, D.; Liu, J.; Liao, X.; Sun, Q. Effects of rhizosphere fungi on the chemical composition of fruits of the medicinal plant Cinnamomum Migao endemic to Southwestern China. BMC Microbiol. 2021, 21, 206. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Astorga-Eló, M.; Zhang, Q.; Larama, G.; Stoll, A.; Sadowsky, M.J.; Jorquera, M.A. Composition, predicted functions and co-occurrence networks of rhizobacterial communities impacting flowering desert events in the atacama desert, Chile. Front. Microbiol. 2020, 11, 571. [Google Scholar] [CrossRef]
- Deng, Q.; Zhang, T.; Xie, D.; Yang, Y. Rhizosphere microbial communities are significantly affected by optimized phosphorus management in a slope farming system. Front. Microbiol. 2021, 12, 2589. [Google Scholar] [CrossRef]
- Maestre, F.; Baquerizo, M.D.; Jeffries, T.; Singh, B. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc. Natl. Acad. Sci. USA 2015, 112, 15684–15689. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, X.; Tai, X.; Sun, L.; Dyson, P. Variation in actinobacterial community composition and potential function in different soil ecosystems belonging to the arid Heihe river basin of Northwest China. Front. Microbiol. 2019, 10, 2209. [Google Scholar] [CrossRef]
- Eisenlord, S.D.; Zak, D.R. Simulated atmospheric nitrogen deposition alters actinobacterial community composition in forest soils. Soil Sci. Soc. Am. J. 2010, 74, 1157–1166. [Google Scholar] [CrossRef]
- Wu, X.; Yang, J.; Ruan, H.; Wang, S.; Yang, Y.; Naeem, I.; Wang, L.; Liu, L.; Wang, D. The diversity and co-occurrence network of soil bacterial and fungal communities and their implications for a new indicator of grassland degradation. Ecol. Indic. 2021, 129, 107989. [Google Scholar] [CrossRef]
- Bratkov, V.M.; Shkondrov, A.M.; Zdraveva, P.K.; Krasteva, I.N. Flavonoids from the genus Astragalus: Phytochemistry and biological activity. Pharmacogn. Rev. 2016, 10, 11–32. [Google Scholar]
- Abdelmohsen, U.R.; Grkovic, T.; Balasubramanian, S.; Kamel, M.S.; Quinn, R.J.; Hentschel, U. Elicitation of secondary metabolism in actinomycetes. Biotechnol. Adv. 2015, 33, 798–811. [Google Scholar] [CrossRef]
- Wei, W.; Zhou, Y.; Chen, F.; Yan, X.; Lai, Y.; Wei, C.; Chen, X.; Xu, J.; Wang, X. Isolation, diversity, and antimicrobial and immunomodulatory activities of endophytic actinobacteria from tea cultivars Zijuan and Yunkang-10 (Camellia Sinensis Var. Assamica). Front. Microbiol. 2018, 9, 1304. [Google Scholar] [CrossRef]
- Corretto, E.; Antonielli, L.; Sessitsch, A.; Höfer, C.; Puschenreiter, M.; Widhalm, S.; Swarnalakshmi, K.; Brader, G. Comparative genomics of microbacterium species to reveal diversity, potential for secondary metabolites and heavy metal resistance. Front. Microbiol. 2020, 11, 1869. [Google Scholar] [CrossRef]
- Hill, P.; Krištůfek, V.; Dijkhuizen, L.; Boddy, C.; Kroetsch, D.; van Elsas, J.D. Land use intensity controls actinobacterial community structure. Microb. Ecol. 2011, 61, 286–302. [Google Scholar] [CrossRef]
- Chevrette, M.G.; Aicheler, F.; Kohlbacher, O.; Currie, C.R.; Medema, M.H. Sandpuma: Ensemble predictions of nonribosomal peptide chemistry reveal biosynthetic diversity across actinobacteria. Bioinformatics 2017, 33, 3202–3210. [Google Scholar] [CrossRef]
- Shi, S.; Cui, L.; Zhang, K.; Zeng, Q.; Li, Q.; Ma, L.; Long, L.; Tian, X. Streptomyces marincola sp. nov., a novel marine actinomycete, and its biosynthetic potential of bioactive natural products. Front. Microbiol. 2022, 13, 860308. [Google Scholar] [CrossRef]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.Z.; El-Enshasy, H.A.; Dailin, D.J.; Suriani, N.L. Recent understanding of soil acidobacteria and their ecological significance: A critical review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef]
- Tao, J.; Liu, X.; Liang, Y.; Niu, J.; Xiao, Y.; Gu, Y.; Ma, L.; Meng, D.; Zhang, Y.; Huang, W.; et al. Maize growth responses to soil microbes and soil properties after fertilization with different green manures. Appl. Microbiol. Biotechnol. 2017, 101, 1289–1299. [Google Scholar] [CrossRef]
- Nam, S.; Alday, J.G.; Kim, M.; Kim, H.; Kim, Y.; Park, T.; Lim, H.S.; Lee, B.Y.; Lee, Y.K.; Jung, J.Y. The relationships of present vegetation, bacteria, and soil properties with soil organic matter characteristics in moist acidic tundra in alaska. Sci. Total Environ. 2021, 772, 145386. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Villarreal Ruiz, L.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Fungal biogeography. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef]
- Kumar, J.; Atri, N.S. Studies on ectomycorrhiza: An appraisal. Bot. Rev. 2017, 84, 108–155. [Google Scholar] [CrossRef]
- Hannula, S.E.; de Boer, W.; van Veen, J.A. In situ dynamics of soil fungal communities under different genotypes of potato, including a genetically modified cultivar. Soil. Biol. Biochem. 2010, 42, 2211–2223. [Google Scholar] [CrossRef]
- Zhu, H.; Gong, L.; Luo, Y.; Tang, J.; Ding, Z.; Li, X. Effects of litter and root manipulations on soil bacterial and fungal community structure and function in a Schrenk’s Spruce (Picea Schrenkiana) forest. Front. Plant Sci. 2022, 13, 849483. [Google Scholar] [CrossRef]
- Guo, Y.; Ren, C.; Yi, J.; Doughty, R.; Zhao, F. Contrasting responses of rhizosphere bacteria, fungi and arbuscular mycorrhizal fungi along an elevational gradient in a temperate montane forest of China. Front. Microbiol. 2020, 11, 2042. [Google Scholar] [CrossRef]
- Dou, Z.-R.; Zhang, F.; Zhou, G.-J.; Gong, G.-F.; Ding, J.-J.; Fan, X.-H. Varicosporellopsis shangrilaensis sp. nov. (Nectriaceae, Hypocreales), a new terricolous species isolated from the rhizosphere soil of Astragalus polycladus in Northwestern Yunnan, China. Phytotaxa 2023, 600, 230–238. [Google Scholar] [CrossRef]
- Nuccio, E.E.; Anderson-Furgeson, J.; Estera, K.Y.; Pett-Ridge, J.; De Valpine, P.; Brodie, E.L.; Firestone, M.K. Climate and edaphic controllers influence rhizosphere community assembly for a wild annual grass. Ecology 2016, 97, 1307–1318. [Google Scholar] [CrossRef]
- Khan, A.; Jiang, H.; Bu, J.; Adnan, M.; Gillani, S.W.; Zhang, M. An insight to rhizosphere bacterial community composition and structure of consecutive winter-initiated sugarcane ratoon crop in Southern China. BMC Plant Biol. 2022, 22, 1–13. [Google Scholar] [CrossRef]
- Wang, P.P.; Yang, L.F.; Sun, J.L.; Yang, Y.; Qu, Y.; Wang, C.X.; Liu, D.Q.; Huang, L.Q.; Cui, X.M.; Liu, Y. Structure and function of rhizosphere soil and root endophytic microbial communities associated with root rot of Panax notoginseng. Front. Plant Sci. 2021, 12, 752683. [Google Scholar] [CrossRef]
- Zuo, Y.-W.; Yu, F.-Q.; Zhang, J.-H.; Xia, C.-Y.; Zhang, H.; Deng, H.-P. Contrasting responses of rhizosphere fungi of Scutellaria tsinyunensis, an endangered plant in Southwestern China. Microbiol. Spectr. 2022, 10, e0022522. [Google Scholar] [CrossRef]
- Milic, N.; Christinaki, A.C.; Benaki, D.; Stavrou, A.A.; Tsafantakis, N.; Fokialakis, N.; Kouvelis, V.N.; Gonou-Zagou, Z. Polyphasic systematics of the fungicolous genus Cladobotryum based on morphological, molecular and metabolomics data. JoF 2022, 8, 877. [Google Scholar] [CrossRef]
- Salvatore, M.M.; Andolfi, A.; Nicoletti, R. The genus Cladosporium: A rich source of diverse and bioactive natural compounds. Molecules 2021, 26, 3959. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant-microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Jiang, L.-L.; Tang, J.-X.; Bo, Y.-H.; Li, Y.-Z.; Feng, T.; Zhu, H.-W.; Yu, X.; Zhang, X.-X.; Zhang, J.-L.; Wang, W. Cytotoxic secondary metabolites isolated from the marine alga-associated fungus Penicillium chrysogenum LD-201810. Mar. Drugs 2020, 18, 276. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable Soil. ISME 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Havlin, J.L. Soil: Fertility and Nutrient Management. In Landscape and Land Capacity; CRC Press: Boca Raton, FL, USA, 2020; pp. 251–265. [Google Scholar]
- Naz, M.; Dai, Z.; Hussain, S.; Tariq, M.; Danish, S.; Khan, I.U.; Qi, S.; Du, D. The soil pH and heavy metals revealed their impact on soil microbial community. J. Environ. Manag. 2022, 321, 115770. [Google Scholar] [CrossRef]
- Ni, S.; Wen, H.; Wilson, G.V.; Cai, C.; Wang, J. A simulated study of surface morphological evolution on coarse-textured soils under intermittent rainfall events. CATENA 2022, 208, 105767. [Google Scholar] [CrossRef]
- Yu, W.; Huang, W.; Weintraub-Leff, S.R.; Hall, S.J. Where and why do particulate organic matter (POM) and mineral-associated organic matter (MAOM) differ among diverse soils? Soil. Biol. Biochem. 2022, 172, 108756. [Google Scholar] [CrossRef]
- Liang, J.; Zou, R.; Huang, Y.; Qin, H.; Tang, J.; Wei, X.; Liang, Y.; Chai, S. Structure and diversity of mycorrhizal fungi communities of different part of Bulbophyllum tianguii in three terrestrial environments. Front. Plant Sci. 2022, 13, 992184. [Google Scholar] [CrossRef]
- Du, Y.; Wang, T.; Jiang, J.; Wang, Y.; Lv, C.; Sun, K.; Sun, J.; Yan, B.; Kang, C.; Guo, L.; et al. Biological control and plant growth promotion properties of Streptomyces albidoflavus St-220 isolated from Salvia miltiorrhiza rhizosphere. Front. Plant Sci. 2022, 13, 976813. [Google Scholar] [CrossRef]
- Li, J.; Ai, J.M.; Liu, X.D.; Jiang, Y.Y.; Zheng, C.C.; Zhao, R.H.; Deng, Z.S. Environmental filtering drives the establishment of the distinctive rhizosphere, bulk, and root nodule bacterial communities of Sophora davidii in hilly and gully regions of the loess plateau of China. Front. Microbiol. 2022, 13, 945127. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | pH | TN (g/kg) | TP (g/kg) | TK (g/kg) | SOM (g/kg) |
|---|---|---|---|---|---|
| AF | 5.29 ± 0.01 a | 3.37 ± 0.02 a | 1.66 ± 0.01 | 13.04 ± 0.09 a | 55.12 ± 0.79 a |
| AA | 5.60 ± 0.01 a | 2.65 ± 0.02 a | 1.00 ± 0.02 | 18.33 ± 0.10 a | 46.32 ± 0.03 a |
| AE | 5.88 ± 0.02 a | 2.18 ± 0.02 a | 0.93 ± 0.04 | 16.98 ± 0.06 a | 40.12 ± 0.13 a |
| Sample ID | Chao1 | Observed Features | Shannon | Simpson |
|---|---|---|---|---|
| AF | 1044.76 ± 98.81 a | 962.67 ± 82.25 a | 4.52 ± 0.11 a | 0.86 ± 0.01 a |
| AA | 908.19 ± 8.32 a | 855.67 ± 11.59 a | 4.82 ± 0.04 a | 0.88 ± 0.01 a |
| AE | 1020.6 ± 8.98 a | 946.67 ± 14.74 a | 4.66 ± 0.09 a | 0.82 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, J.-J.; Zhou, G.-J.; Chen, X.-J.; Xu, W.; Gao, X.-M.; Zhang, Y.-Z.; Jiang, B.; Li, H.-F.; Wang, K.-L. Analysis of Microbial Diversity and Community Structure of Rhizosphere Soil of Three Astragalus Species Grown in Special High-Cold Environment of Northwestern Yunnan, China. Microorganisms 2024, 12, 539. https://doi.org/10.3390/microorganisms12030539
Ding J-J, Zhou G-J, Chen X-J, Xu W, Gao X-M, Zhang Y-Z, Jiang B, Li H-F, Wang K-L. Analysis of Microbial Diversity and Community Structure of Rhizosphere Soil of Three Astragalus Species Grown in Special High-Cold Environment of Northwestern Yunnan, China. Microorganisms. 2024; 12(3):539. https://doi.org/10.3390/microorganisms12030539
Chicago/Turabian StyleDing, Jia-Jie, Guo-Jun Zhou, Xiao-Jie Chen, Wei Xu, Xing-Mei Gao, Yong-Zeng Zhang, Bei Jiang, Hai-Feng Li, and Kai-Ling Wang. 2024. "Analysis of Microbial Diversity and Community Structure of Rhizosphere Soil of Three Astragalus Species Grown in Special High-Cold Environment of Northwestern Yunnan, China" Microorganisms 12, no. 3: 539. https://doi.org/10.3390/microorganisms12030539
APA StyleDing, J.-J., Zhou, G.-J., Chen, X.-J., Xu, W., Gao, X.-M., Zhang, Y.-Z., Jiang, B., Li, H.-F., & Wang, K.-L. (2024). Analysis of Microbial Diversity and Community Structure of Rhizosphere Soil of Three Astragalus Species Grown in Special High-Cold Environment of Northwestern Yunnan, China. Microorganisms, 12(3), 539. https://doi.org/10.3390/microorganisms12030539