Abstract
Listeria monocytogenes (L. monocytogenes) is a pathogen that is transmitted through contaminated food and causes the illness known as listeriosis. The virulence factor InlA plays a crucial role in the invasion of L. monocytogenes into the human intestinal epithelium. In addition, InlA enhances the pathogenicity of host strains, and different strains of L. monocytogenes contain varying variations of InlA. Our study analyzed a total of 4393 published L. monocytogenes genomes from 511 sequence types (STs) of diverse origins. We identified 300 unique InlA protein sequence types (PSTs) and revealed 45 highly mutated amino acid sites. The leucine-rich repeat (LRR) region was found to be the most conserved among the InlA, while the protein A (PA) region experienced the highest mutation rate. Two new types of mutations were identified in the B-repeat region of InlA. Correspondence analysis (CA) was used to analyze correlations between the lineages or 10 most common sequence types (STs) and amino acid (aa) sites. ST8 was strongly correlated with site 192_F, 454_T. ST7 exhibited a strong correlation with site 51_A, 573_E, 648_S, and 664_A, and it was also associated with ST6 and site 544_N, 671_A, 738_B, 739_B, 740_B, and 774_Y. Additionally, a strong correlation between ST1 and site 142_S, 738_N, ST2 and site 2_K, 142_S, 738_N, as well as ST87 and site2_K, 738_N was demonstrated. Our findings contribute significantly to the understanding of the distribution, composition, and conservation of InlA in L. monocytogenes. These findings also suggest a potential role of InlA in supporting molecular epidemiological tracing efforts.
1. Introduction
Listeria monocytogenes (L. monocytogenes) is a foodborne pathogen that can cause gastroenteritis in healthy individuals, meningitis in immunocompromised individuals, and may lead to abortions in pregnant women, with a fatality rate of up to 20–30% [1]. L. monocytogenes is a Gram-positive saprophytic species that can survive in harsh environments, such as low temperatures, partial sanitizers, a broad range of pH, and high salt levels, among other conditions [2]. As a facultative intracellular parasite, L. monocytogenes can invade and survive within a wide range of non-phagocytic cells.
InlA is the first identified virulence factor linked with L. monocytogenes invasion. It can be located on the host cell membrane by binding to the E-cadherin receptor and enhances the crossing of both the intestinal and placental barrier [3]. Point mutations in the InlA may result in premature stop codons (PMSC) and truncated InlA, which makes it a secretory protein and cannot be anchored to the bacterial cell wall. This weakens the ability of L. monocytogenes to invade intestinal epithelial cells. Previous research has shown this on Caco-2 epithelial cells [4,5,6,7].
L. monocytogenes is categorized into four lineages (I, II, III, and IV), with I and II being the most prevalent, showing noticeable differences in sources [8]. Most of the strains in lineage I are clinical isolates, whereas the lineage II strains are predominantly isolated from contaminated foods [9]. Lineages III and IV strains are less common and obtained mainly from animals. The multi-locus sequence typing (MLST) of L. monocytogenes is determined by seven housekeeping genes [10]. The clonal complex (CC) refers to sequence types that vary only in one allele. Studies suggest that CC1, CC2, CC4, and CC6 are the primary clone complexes linked to listeriosis, whereas CC9 and CC121 are common clone complexes of L. monocytogenes in food [11,12,13].
The intestinal barrier represents the initial defense against L. monocytogenes infection, and InlA is pivotal for enabling the bacteria to invade [14]. A lot of studies have reported that PMSCs in InlA, resulting in a truncated form of InlA, suggesting attenuated pathogenicity [15,16]. For example, in the United States, the detection of L. monocytogenes strains harboring stop codons has been demonstrated at 35–45% in food isolates, while human isolates only account for 5.1% [17]. At present, many types of PMSC mutations in the InlA have been documented [18,19,20]. However, Wang et al. showed that the presence of PMSC type 19 at position 976 may not impact the ability of L. monocytogenes to adhere to and invade epithelial cells [21]. Moreover, Dellafiora drew attention to missense mutations, which are often overlooked, and suggested that they should be investigated for their potential effects on cell invasion processes [22]. Indeed, there is a lack of comprehensive analysis of InlA amino acid sequence characteristics. Early research on the InlA in L. monocytogenes solely focused on the presence or absence of PMSC, using a small number of total genomes or a few STs of interest [23]. Nowadays, thousands of L. monocytogenes genomes are available in public databases. We conducted an InlA examination on L. monocytogenes utilizing a significantly larger data set than prior studies [9,24,25]. The aim of this study was to investigate the distribution and length of InlA in L. monocytogenes, identify the characteristics of InlA protein sequences among strains of different lineages, sources and STs, and obtain more understanding about the role of InlA in causing listeriosis.
2. Materials and Methods
2.1. Genomes of Listeria monocytogenes
A total of 4393 genomes of L. monocytogenes were collected from various sources and diverse geographic locations, including 318 newly sequenced genomes from our laboratory (Chinese Center for Disease Control and Prevention), and 4075 publicly available assembled genomes (available to download until March 2022) from 43 other countries on six continents (Supplementary Table S1). There were 695, 1580, 167, and 1090 strains from humans, food, animals, and the environment respectively, with 861 strains from an unknown source.
2.2. MLST Analysis
Using seven housekeeping genes (abcZ, bglA, cat, dapE, dat, ldh, and lhkA), the local BLAST method was employed to determine the ST type of the strains. If the MLST of the strain is unknown, the genome sequence is submitted to the Genome Pasteur Database website (https://bigsdb.pasteur.fr/cgi-bin/bigsdb/bigsdb.pl?db=pubmlst_listeria_seqdef&page=sequenceQuery (accessed on 7 September 2023)) for MLST determination (Supplementary Table S1).
2.3. Alignment and Analysis of InlA Sequence
MEGA software (version 11) was used to align and obtain amino acid sequences of the InlA. Protein types of InlA were identified by comparing them with the reference strain (EGD-e, NC_003210.1) by using local BLAST methods [20]. An amino acid difference is a new protein sequence type (PST). The presence of an asterisk (∗) in the matched query sequences indicated a PMSC happening in the genome assembly sequences. The mutation sites were identified by BLASTN with the reference sequence, while the specific types of PMSC were referenced from previous studies [18,19,20]. The graphical representation of the amino acid probabilities at each site was generated by WebLogo [26].
2.4. Statistical Analysis
The relationship among the L. monocytogenes lineages or STs (columns) and the aa of the InlA site (row) (Table S2) was explored by corresponding analysis (CA) in R language [19]. The angle between the row and column arrows represents the correlation between them, and the smaller the angle, the stronger the correlation.
3. Results
3.1. Distribution Characteristics and MLST of L. monocytogenes Genomes
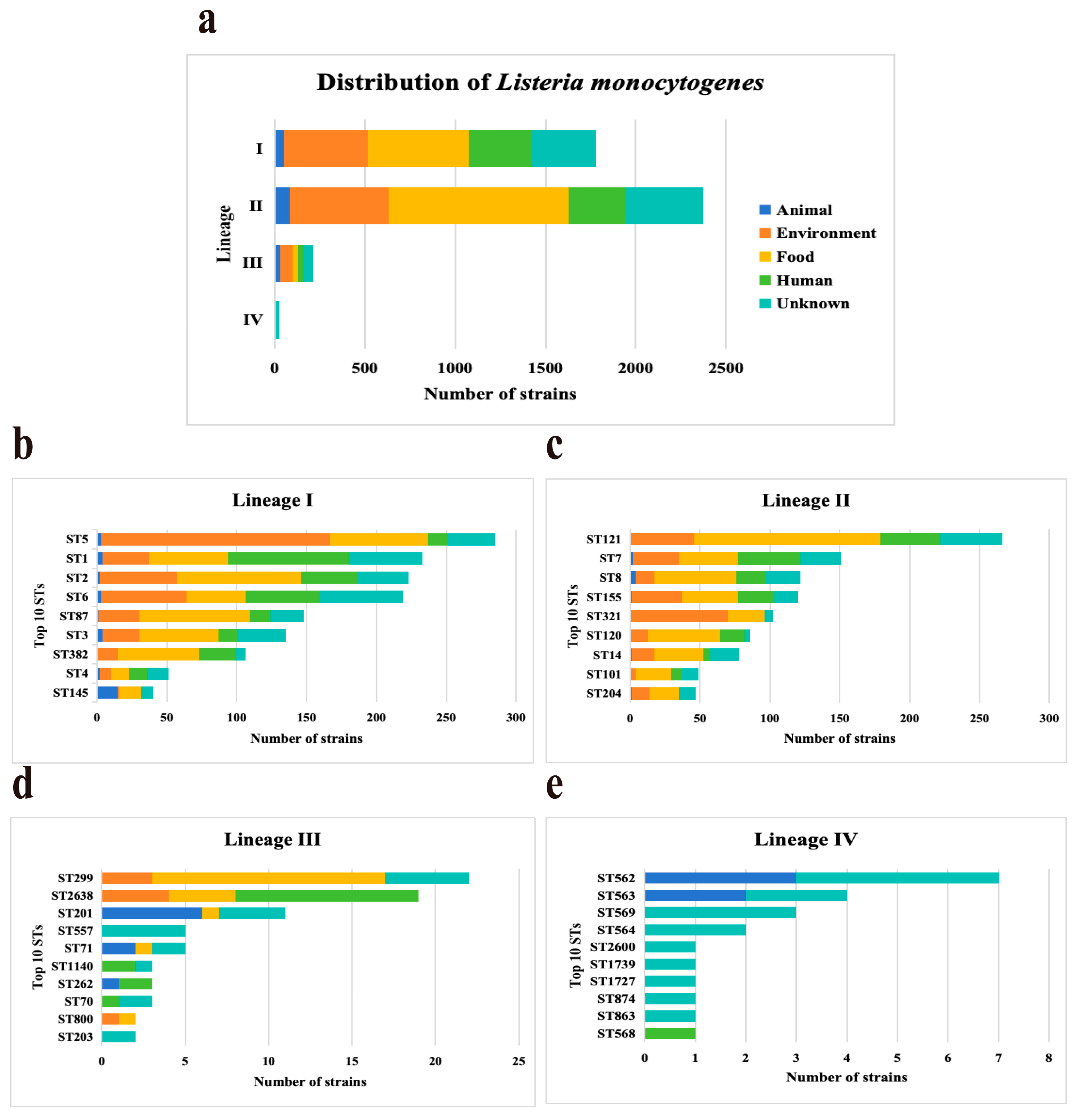
Four lineages were identified among 4393 L. monocytogenes strains, including lineage I (n = 1781, 40.54%), II (n = 2375, 54.06%), III (n = 213, 4.85%), and IV (n = 24, 0.55%). Lineages distributions of L. monocytogenes strains were from four sources (Figure 1a). Lineages I, II, and III accounted for a similar proportion of strains from sources. Lineages IV strains were predominant in the unknown with a frequency of 85.71% (18/21). Based on MLST, all the studied isolates were classified into 511 STs and grouped into 290 CCs. ST distributions of L. monocytogenes strains were different in four lineages (Figure 1). ST5 (n = 285, 16%), ST121 (n = 266, 11.2%), ST299 (n = 22, 10.33%), and ST562 (n = 7, 29.17%) strains were predominant in Lineages I, Lineages II, Lineages III, and Lineages IV, respectively. All the L. monocytogenes strains selected in this study carried the InlA.
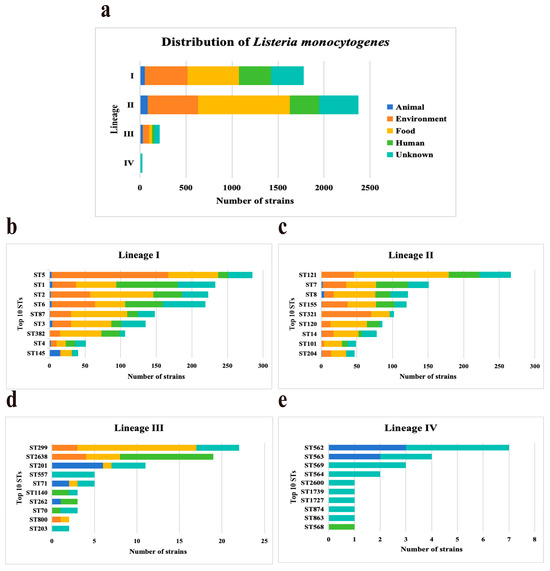
Figure 1.
Distribution characteristics and domain sequence types of Listeria monocytogenes. (a) Distribution of four Lineages (I, II, III, and IV) in five sources. (b–e) Distribution of top 10 sequence types (STs) in four Lineages among the 4393 L. monocytogenes strains.
3.2. The Classification and Analysis of InlA Sequence Types
In this study, a total of 300 PSTs of InlA were identified from the strains and classified into two groups based on the presence of PMSC: the non-PMSC group (group A) and the PMSC group (group B). Groups A and B were then further subdivided into six (by InlA length) or seven subgroups (by truncated in the domain of InlA). Group A consisted of 262 PSTs, accounting for 77.44% of the total strains (3402/4393), while group B had 38 PSTs, accounting for 22.56% (991/4393). In group A, there were 2995 strains from A-1 (800aa), 351 from A-2 (797aa), 33 from A-3 (730aa), 21 from A-4 (799aa), 1 from A-5 (733aa), and 1 from A-6 (801aa). Among these, the PSTs that encode 800 amino acids exhibited maximum diversity, encompassing 162 types.
The InlA sequences of group B were truncated in the signal sequence (SS), α-helix region, Leucine rich repeats (LRR) region, inter-repeat (IR) region, B-repeat region, Protein A (PA) region, and Leu-Pro-X-Thr-Gly C terminal cell wall anchor motif (LPXTG), X for any amino acid region with 3, 3, 10, 2, 12, 2, and 1 truncation types, respectively (Figure 2f). B-5 (B-repeat) exhibited the highest number of strains and the most diverse PSTs in group B. While the number of strains in B-4 (IR) exceeded those in B-3 (LRR), the PSTs of B-3 were noticeably more diverse. Three types of truncations were identified in group B-1 (SS), alongside a greater number of strains (n = 155). Similarly, in group B-2 (α-helix), three truncation types were found, but only six strains. B-5 and B-6 each had four and one strains, respectively, and these strains belonged to two or one PSTs, respectively.
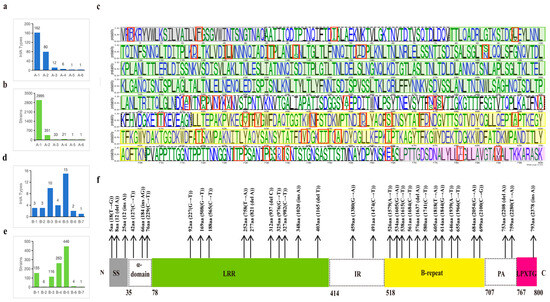
Figure 2.
Distribution characteristics of InlA sequences in this study. Distribution of InlA types (a) or strains (b) in group A. Map of the 261 mutation sites in the protein sequences of group A, where the red boxes mark the sites where more than 10 types of protein sequences are mutated (c). Distribution of InlA types (d) or strains (e) in group B. The length of truncated of InlA and nucleotide position of mutation in group B (f).
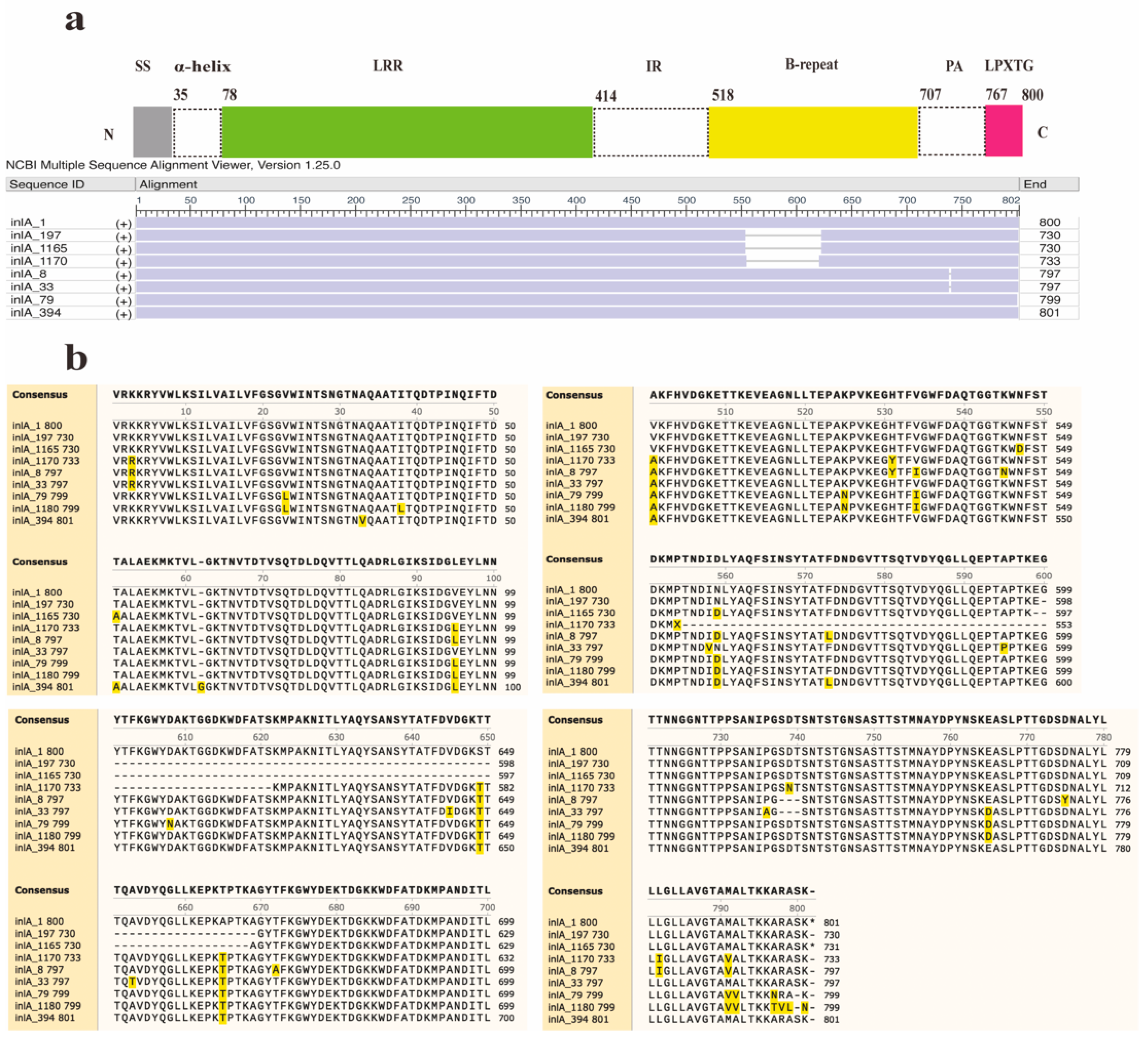
Furthermore, two new InlA mutations encoding proteins with 730 and 733 amino acids were discovered in this study (Figure 3). Out of the 12 distinct types of PST encoding 730 amino acids, 11 sequences were deficient in 70 amino acids at positions 599–668, whereas another sequence (InlA_1165) had a deficiency of 70 amino acids at positions 598–667. The InlA sequences of group A-3 encode 730 amino acids (Figure 3b). It was found predominantly in lineage II (30/33) strains and was distributed in lineages I and III. The sequence of group A-5 encodes 733 amino acids exhibited a deletion of 67 amino acids at positions 554–621 compared to InlA of EGD-e. All PSTs in Group A-2 encode 797 amino acids and showed a deletion of three amino acids (SDT) at positions 737–739. There is a total of 80 InlA PSTs in Group A-2, predominantly from lineage III strains, but also including lineage I and II strains. The most common protein types in this group were PST 6 (InlA_8) and PST 25 (InlA_33), which also existed in China. Furthermore, Group A-4 results from a deletion of one amino acid (S) at position 799, leading to 799 amino acids. Six PSTs encoding 799 amino acids in group A-4 were solely identified in lineage IV strains. Finally, group A-6 arises from an insertion of a ‘G’ amino acid at position 62, producing 801 amino acids.
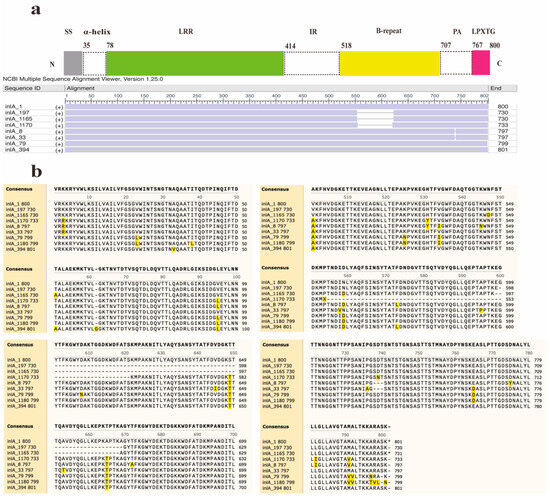
Figure 3.
Alignment and schematic representation of the InlA mutant and EGDe (InlA_1). (a) Represents the overall schematic diagram of InlA alignment, and (b) shows the specific location of the mutation. SS: Signal Sequence.
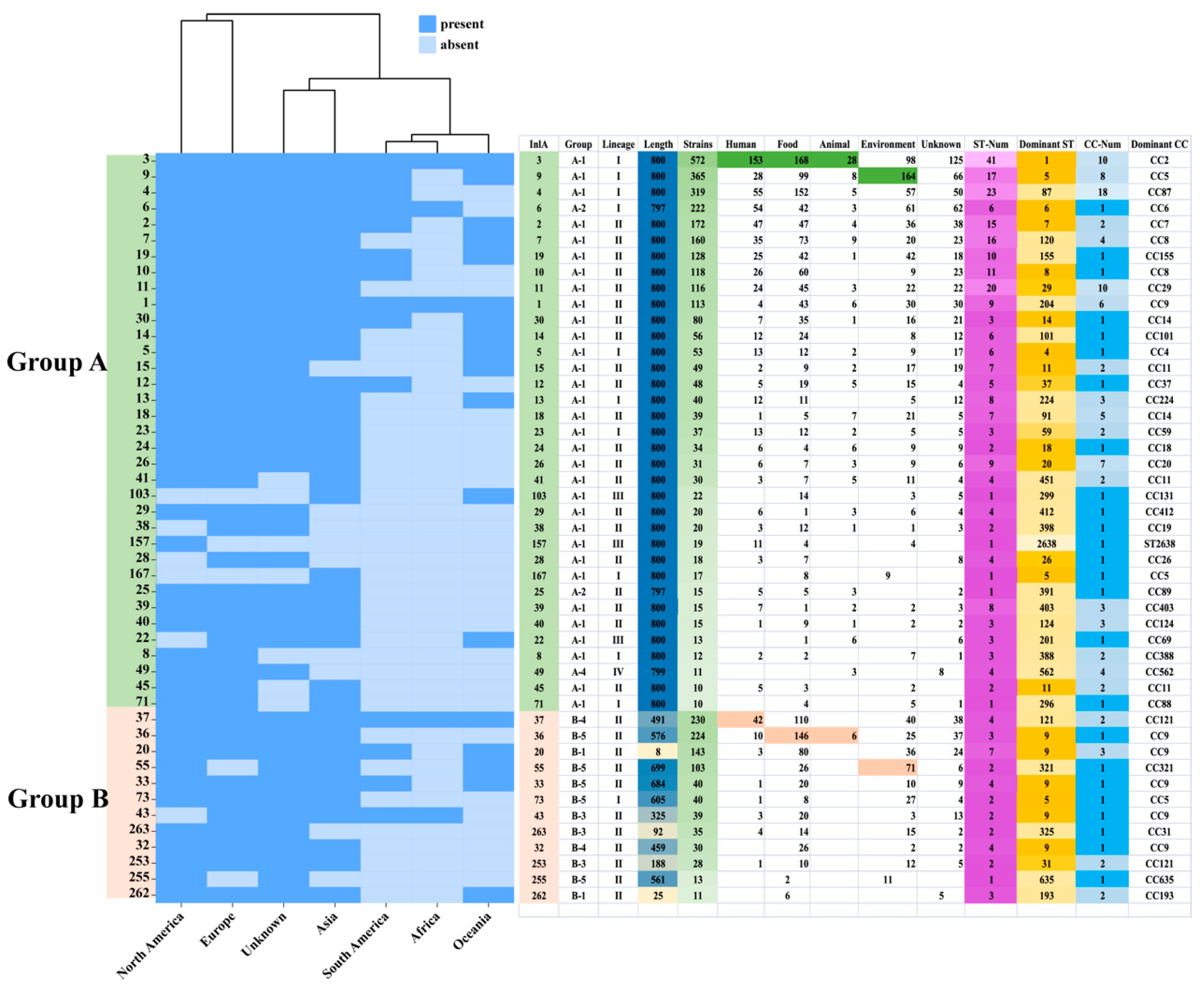
To investigate the relationship between the InlA and the strains, 47 InlA PSTs (n ≥ 10, 3964 strains) were selected for further analysis. We found out that most protein types are distributed in multiple continents (Figure 4). However, PST 167 was only found in Asian strains; PST 157, on the other hand, was only found in North American strains. Out of these 47 PSTs, 35 PSTs were grouped into group A, and the most common PSTs detected in humans, food, animals, and environmental sources were PST 3, PST 3, PST 3, and PST 9, respectively. Figure 4 shows that out of 32 PSTs that encode 800 aa, 15 corresponded to a single CC, while the remainder corresponded to multiple CCs. Within group A, PST 3 was the most common corresponding to 10 CCs, with CC2 as the dominant type. PST 9 followed, corresponding to 8 CCs, and CC5 was the dominant type. PST 4 corresponded to 18 CCs, in which CC87 was the most dominant CC type. Group B strains were predominantly concentrated in lineage II, followed by lineage I. Only a single strain belonging to group B-7 was identified in lineage III, which encoded 793 amino acids. Lineage I strains were distributed mainly in groups B-1, B-3, B-5, and B-6. Among them, B-5 (B-repeat) represented the highest proportion, accounting for 78.79% (52/66) of the total. Lineage II strains had a broader distribution ranging from groups B-1 to B-6. Additionally, group B-5 (B-repeat) accounted for the highest proportion again, at 42.64% (394/924). Among groups B-1, B-5, and B-6, CC9 was the most dominant CC type. Figure 4 shows that out of the 12 PSTs (n ≥ 10), in group B, 8 PSTs correspond to one CC type while the other 4 PSTs correspond to multiple CC types. PST 37, PST 20, PST 253, and PST 262 correspond to 2, 3, 2, and 2 CC types, respectively, with the dominant CC types of CC121, CC9, CC121, and CC193.
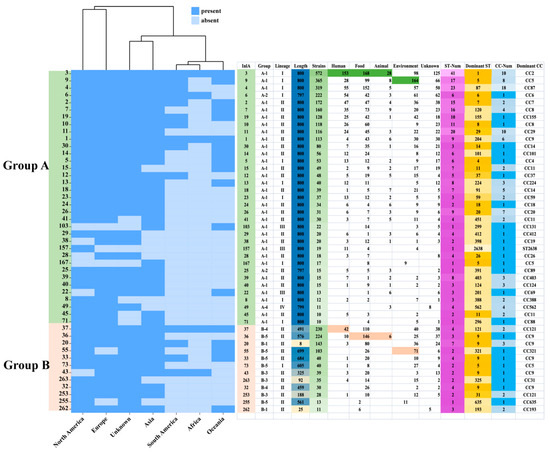
Figure 4.
Distribution of 47 InlA types (n ≧ 10). The InlA number, the lineage, source, location, ST, and CC are shown on the right rows. The color legend is shown above.
Group B includes 67 human strains, representing 6 CCs with CC121 predominating (43/67) (Table 1). Among the human strains, 64/67 are from lineage II with the remaining 3 strains belonging to lineage I. Truncated InlA regions were identified in the following distribution: 62.69% (42/67) of the strains in the IR region (B-4), 20.9% (14/67) in the B repeat region (B-5), 11.94% (8/67) in the LRR region (B-3), and 4.48% (3/67) in the SS region (B-1). Additionally, these strains included 9 PMSC types, with the PMSC type 6 (491 aa) being the most prevalent, accounting for 62.69% (42/67).

Table 1.
Characteristics of truncated internalin A in clinical Listeria monocytogenes strains circulating in this study.
3.3. The Classification and Analysis of InlA Sequence Types
The 261 PSTs of InlA in Group A (except for the sequence of one strain, 801aa) were compared with the InlA of EGD-e, and further analysis revealed that 526 were fully conserved and 274 were variable sites (Table S1). There were 45 amino acid mutation sites that exhibited a substitution rate higher than 1.47% (n > 50, 50/3401). The mutation rates in various structural domains of amino acids, named SS, α-helix, LRR, IR, B-repeat, PA, and LPXTG regions were calculated to be 8.82% (3/34), 9.3% (4/43), 1.79% (6/336), 6.73% (7/104), 8.47% (16/189), 10% (6/60), and 8.82% (3/34), respectively. The highly conserved IR region was best described as an immunoglobulin (Ig) and is structurally the most flexible part of the internalin domain [27]. In our study, the LRR region of InlA exhibited the highest conservation, while the PA region displayed the most frequent mutations, followed by the α-helix region, these two high mutation regions may have no effect on binding receptor function [28].
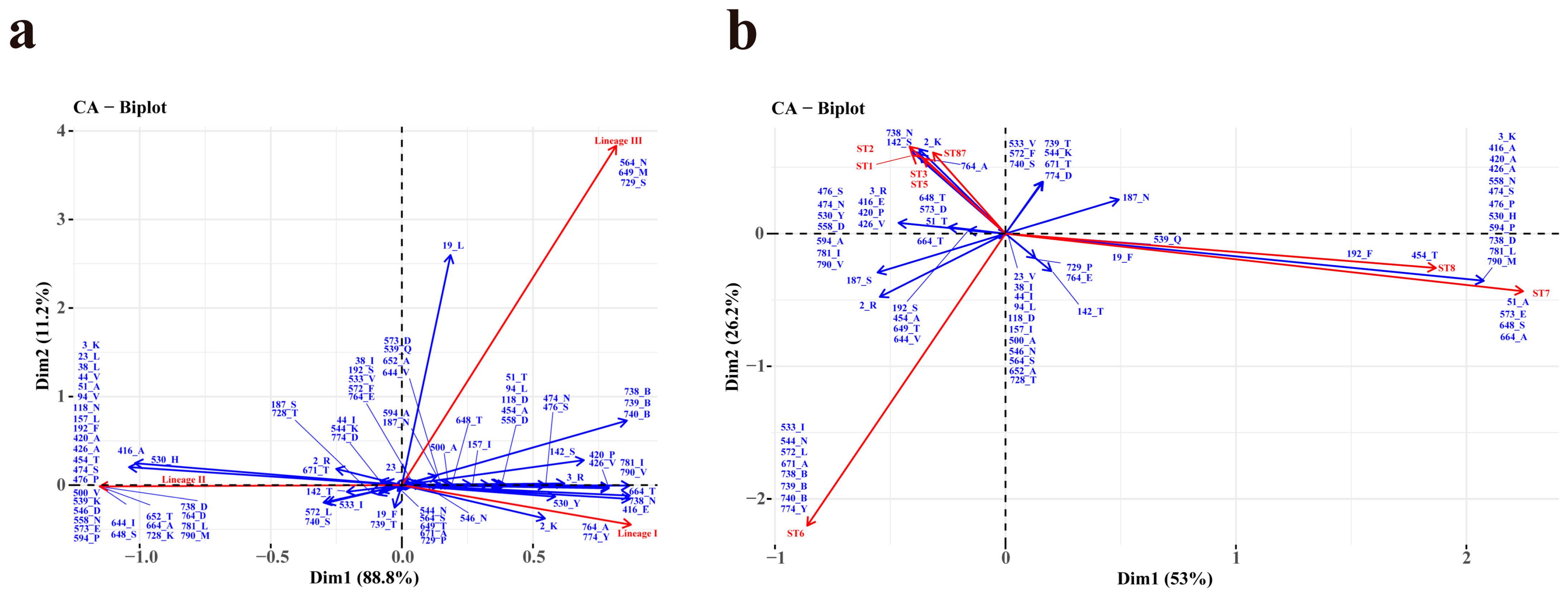
Certain amino acid sites exhibit specific amino acids, including acidic, basic, polar, or nonpolar residues, and a subset of these sites (n > 50) demonstrate a pronounced correlation with lineages (Figure 5a). Specifically, Site 764_A, and 774_Y are significantly associated with lineage I. A minor association between site 2_K and lineage I was also observed. Site 3_K, 23_L, 38_L, 44_V, 51-A, 94_V, 118_N, 157_L, 192_F, 420_A, 426_A, 454_T, 474_S, 476_P, 500_V, 539_K, 546_D, 573_E, 594_P, 644_I, 648_S, 652_T, 664_A, 558-N, 728_K, 738_D, 764_D, 781_L, and 790_M were demonstrated a strong correlation with lineage II. A minor association among site 416_A, 530_H and lineage II was also observed. Site 564_N, 649_M, and 729_S showed a high correlation with lineage III. A minor association between site19_L and lineage III was also observed. On the contrary, lineage IV did not exhibit a particular association with any specific site.
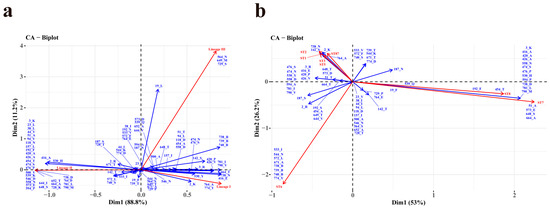
Figure 5.
Correspondence analysis (CA) between the aa site (n > 50, represented in blue) and lineages (a) or ten most prevalent STs (b) (represented in red) of Listeria monocytogenes strains. “B” represents the deletion of aa.
In Figure 5b, the associations among the ten most prevalent STs and aa sites (n > 50) were shown using CA in R. The graphical representation shows a strong correlation between ST1 and site 142_S, 738_N, ST2 and site 2_K, 142_S, 738_N, as well as ST87 and site 2_K, 738_N. Correspondingly, ST7 is related to site 51_A, 573_E, 648_S, and 664_A. Moreover, ST8 is strong related to site 192_F and 454_T, and as well as among ST6 and site 544_N, 671_A, 738_B, 739_B, 740_B, and 774_Y. Additionally, the site 3_K, 416_A, 420_A, 426_A, 474_S, 530_H, 558_N, 594_P, 738_D, 781_L, and 790_M demonstrated a minor correlation with ST7 and ST8. In addition, site 738_N and 764_A exhibited a strong correlation with ST3 and ST5.
A correlation was observed between amino acids at 21 out of 45 sites (sites 2, 3, 19, 23, 38, 44, 94, 142, 157, 187, 474, 500, 533, 564, 572, 644, 648, 739, 740, 781, and 790) and a specific lineage, without any alteration in their polarity or charge (Table S2). The remaining 24 sites exhibited distinct attributes, with 10 sites (site 118, 530, 539, 544, 546, 558, 573, 728, 738, and 774) altering only the charge without affecting the polarity, and 14 sites (site 51, 192, 416, 420, 426, 454, 476, 594, 649, 652, 664, 671, 729, and 764) changing the polarity of the amino acid side chain. Sites 51_A (α-helix), 192_F (LRR), 420_A, 426_A, 454-T, 476_P (IR), 594-P, 649_B, 652_B, 652_T, and 664_A (B-repeat) with changed polarity exhibited a strong correlation with lineage II. Meanwhile, sites 416_E and 764_A had a strong correlation with lineage I, and sites 649_M, 664_M, and 729_S significantly correlated with lineage III. It is worth mentioning that site 420 (n = 1) and site 594 (n = 1) were both mutated to threonine, resulting in a change from nonpolar amino acids to polar, uncharged amino acids.
4. Discussion
A thorough comprehension of the polymorphism, truncation types, and distribution characteristics of Internalin A is crucial for studying and evaluating the virulence of L. monocytogenes, as it constitutes a significant virulence factor [29]. Furthermore, advances in whole-genome sequencing technology enable us to analyze the epidemiological features of L. monocytogenes strains via the InlA and to investigate their potential virulence by InlA sequences.
In this study, a total of 4393 strains of L. monocytogenes were divided into 4 lineages, which were then clustered into 290 CCs. The dominant CCs varied from different sources, with CC1 (99/695), CC9 (310/1580), CC2 (17/167), and CC5 (166/1090) were the dominant CCs for strains obtained from humans, food, animals, and environmental sources, respectively. Studies have shown that CC1 and CC2 are the most prevalent clones in clinical strains, whereas CC9 is often found in food and environmental samples and is a clone with low-virulence [19]. ST5 and ST121 are known to exhibit higher tolerance to adverse conditions and their capacity to adapt to such environments has been found to be crucial in the formation of biofilms, thereby affecting the persistence of L. monocytogenes in the environment [30,31,32].
The PST 3 (lineage I, 800 aa) of InlA exhibited the highest carriage rates among strains found in humans (153/659), animals (28/167), and food (168/1580). This suggested that the strains of PST 3 existed in the dominant three sources may be due to the most common PST3 strains. PST 3 corresponded to 10 CCs, with CC1 (269/572) as the dominant type. The PST 9 (lineage I, 800 aa), with the dominant CC type being CC5 (221/365), has the highest carriage rate among environmental strains (164/1090). The correlation between InlA and CC is typically one-to-one with a few exceptions. For example, PST 4 (lineage I, 800 aa) corresponded to 18 CCs, with CC87 as the dominant type. CC87 is made up of ST87 and three other STs, which is the most common subpopulation connected with food, recreational beach sands, and human clinical infections in China [33,34,35]. Among groups B-1 (SS), B-5 (B-repeat), and B-6 (PA), CC9 was the most dominant CC. This observation could be attributed to the mutations that CC9 strains have undergone to adapt to their respective environments, primarily food and the surroundings. Therefore, InlA can serve as a biomarker of virulence for strains.
Based on the alignment of 300 InlA sequences with that of EGD-e, the LRR region was identified as the most conserved, while PA was the most active mutation region. The LRR region is known to play a critical role in InlA binding, while the LPXTG region serves as a membrane anchoring region [36,37]. These two key functional regions are highly conserved, indicating that they were necessary for the InlA. Among the truncated InlA variants, the B repeat region was the most frequently truncated, followed by the IR region, then the SS region, LRR region, α-helix region, PA region, and LPXTG region [35]. There is no necessary link between the frequency of amino acid mutations and the truncation of InlA, which can possibly be attributed to the environmental pressures of coping with complex external factors. This study identified two new InlA mutations, which encode 730 and 733 amino acids, respectively. These mutants do not fall under the PMSC classification. A previous study indicated that knocking down the B repeat or PA regions has no impact on the invasion of L. monocytogenes [28], but these findings were limited to artificially constructed InlA mutants. Further confirmation is required for the invasion of naturally occurring truncated InlA with a B repeat region mutation. Our study found that 33 strains from 13 STs harbored 730aa-InlA, including 30 lineage II strains, one lineage I strain, and one lineage III strain, which shows that the 730aa mutation type is dominant in lineage II strains. All of them belong to the US Food and Drug Administration's surveillance project for the rapid detection of foodborne contamination events (PRJNA304956). Twenty-nine strains were collected from Italy and four from the United States. The predominant ST of the Italian strains was ST204 (6/29), while all four US strains (collected from Snow King Peach) belonged to ST1331. Drupes, including peaches, are generally considered to pose a low risk of foodborne illness. However, in 2014, an outbreak involving L. monocytogenes highlighted the potential of drupes as a new food vector for transmitting L. monocytogenes, based on cases of human listeriosis [38]. Italy faced a continuous outbreak caused by an invasive lineage II strain of L. monocytogenes [39,40]. These findings indicate the importance of taking this type of mutation seriously.
Previous research has shown that the InlA, which encodes 797 amino acids, experiences a deletion of 3 amino acids in the C-terminal sequence and still maintains a complete LPXTG region [35]. We discovered that InlA PST 6, which encodes 797 amino acids, is more commonly present among the strains and belongs to lineage I, serotype IVb, which is similar to earlier research [20]. Strains of serotype IVb have been responsible for several listeriosis outbreaks [11]. For instance, in 2017–2018, the outbreak in South Africa was caused by ready-to-eat meat products contaminated with these strains, leading to a widespread infection with a mortality rate of 28.6%. The strains associated with the outbreak were of serotype 4b and ST6 genotype [35,41]. Our research indicates that the InlAs encoding for 797 amino acids exhibited the most diversity in lineage III. The strain had a 3-amino acid deletion in the transmembrane region near the C-terminus of InlA and was accountable for the 2002 outbreak of contaminated ready-to-eat meat. However, this type still retained its virulence [42,43]. Kovacevic and colleagues reported a similar variation in which three amino acids were deleted from a strain isolated from food and food processing environments in British Columbia, Canada. This variation was able to invade Caco-2 cells [43]. Although we did not test the invasion of these InlA proteins, we inferred from their sequence alone that they have invasive capabilities.
Most L. monocytogenes strains belong to Lineages I and II, while Lineages III and IV are infrequent and are primarily found in ruminants [44,45]. Our study discovered that 22.56% (991/4393) strains carried truncated InlA belong to group B, with 93.24% (924/991) of those strains being lineage II. Additionally, InlA of lineage II strains revealed a higher level of diversity. A percentage of 38.91% of strains of lineage II were categorized as group B, whereas only 3.71% of lineage I strains were categorized as group B, which is consistent with previous research results [19]. Lineage II strains exhibit the largest number of truncated InlA variants that may cause attenuated pathogenicity [46]. The association between this diversity and the complex food environment and stress exposures of this strain's source requires further investigation and confirmation.
Researchers have demonstrated that strains containing InlA surface protein with PMSCs display diminished invasive capacity within cells [1,9,21]. Nevertheless, other researchers have observed that these strains still have the capability of causing disease [19,24,47]. Therefore, varying types of PMSC in InlA may impact the virulence of L. monocytogenes differently. We analyzed 67 isolates of L. monocytogenes obtained from humans and carrying truncated InlA, with the highest frequency being PMSC type 6 (231/991). Type 6 encodes 491 amino acids and is classified under group B-4. Most of these strains belonged to CC121 (43/67) and CC9 (16/67). Previous reports indicated that ST9 and ST121 strains often possess truncated InlA and have strong adaptability to survive in food and plants, whereas they play a minor role in causing clinical cases and exhibit low invasive activity [19,33,48]. Most truncated InlA from human strains have the intact LRR region, which is the key domain contact with the receptor on the host cell. Three strains carried truncated InlA (CC3, 646aa and CC5, 605aa) belonging to lineage I. Usually, these strains belong to CC3 or CC5 presented intact InlA and were linked to listeriosis cases. Additionally, it should be emphasized that the PMSC mutation type of the three clinical isolates of ST9 was PMSC type 19. According to the study, the presence of PMSC 19 at position 976 may not impact the ability of L. monocytogenes to adhere to and invade epithelial cells [21]. This indicates that the existence of PMSCs may affect the ability of the strain to invade depending on their nucleotide position. It is, therefore, essential to verify not only the presence or absence of the InlA mutation but also the type of PMSC mutation. However, additional verification and studies on molecular mechanisms are required to fully understand the precise impact of different mutation types on their function.
Proteins are composed of amino acids and perform a wide range of physiological functions in the body [49]. Previous studies have identified that Phe367Ala and Tyr343Ala, both of which were shown to significantly reduce the InlA-Ecad interaction, and Tyr369Ser and the double mutant Ser192Asn-Tyr369Ser, which were shown to increase the InlA-Ecad complex formation [22]. This indicated that single amino acid changes could affect the function of InlA. Then we found that site 192, which has been identified as a single residue causing intermolecular contact, exhibited polarity change. Strains that exhibit a change from S to F at this location belong to CC8 and are situated within the LRR region. Therefore, it is hypothesized that this alteration could potentially impact the function of InlA. However, further investigation is required to determine the exact mechanism of action. The remaining 13 sites consist of one in the α-helix and the remaining in the IR, B repeat, and PA region. These sites have no direct association with the primary binding site [27,28]. Although no previous studies have reported that altering these sites could affect the function of InlA, we found that the polarity of these sites is altered, which may disrupt hydrogen bonding, ionic bonding, or hydrophobic interactions within the protein, thereby altering the folding mode and stability of the protein. This could consequently affect its function and interactions in space [50]. There are some sites that show a higher correlation with lineages and STs, which should also be focused on. Nevertheless, further studies need to be conducted to determine the exact effects.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms12030485/s1, Table S1: Information and 274 variable sites of 4393 Listeria monocytogenes genomes; Table S2: The relationship among the lineages or STs and the aa of InlA site was explored by CA.
Author Contributions
Conceptualization, C.Y. and L.L. (Lingling Li);methodology, Y.W., L.L. (Lingling Li) and J.P.; validation, L.L. (Lingling Li); formal analysis, Y.W. and L.L. (Lingling Li); investigation, Y.W., L.L. (Lingling Li), J.P., J.C., L.L. (Lingyun Liu), P.M., H.S., X.L. and C.Y.; writing—original draft preparation, L.L. (Lingling Li); writing—review and editing, Y.W., L.L. (Lingling Li) and C.Y.; visualization, L.L. (Lingling Li); supervision, H.S., X.L. and Y.W.; project administration, L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by National Institute for Communicable Disease Control and Prevention, China CDC (2021ZZKT003) (102393220020020000029).
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Acknowledgments
We thank the Shunshi Ji for his contributions to the data analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Su, X.; Cao, G.; Zhang, J.; Pan, H.; Zhang, D.; Kuang, D.; Yang, X.; Xu, X.; Shi, X.; Meng, J. Characterization of internalin genes in Listeria monocytogenes from food and humans, and their association with the invasion of Caco-2 cells. Gut Pathog. 2019, 11, 30. [Google Scholar] [CrossRef]
- Wagner, E.; Zaiser, A.; Leitner, R.; Quijada, N.; Pracser, N.; Pietzka, A.; Ruppitsch, W.; Schmitz-Esser, S.; Wagner, M.; Rychli, K. Virulence characterization and comparative genomics of Listeria monocytogenes sequence type 155 strains. BMC Genom. 2020, 21, 847. [Google Scholar] [CrossRef] [PubMed]
- Ortega, F.; Rengarajan, M.; Chavez, N.; Radhakrishnan, P.; Gloerich, M.; Bianchini, J.; Siemers, K.; Luckett, W.; Lauer, P.; Nelson, W.; et al. Listeria monocytogenes Adhesion to the host cell surface is sufficient to mediate entry into epithelial cells. Mol. Biol. Cell 2017, 28, 2945–2957. [Google Scholar] [CrossRef] [PubMed]
- Lyautey, E.; Hartmann, A.; Pagotto, F.; Tyler, K.; Lapen, D.R.; Wilkes, G.; Piveteau, P.; Rieu, A.; Robertson, W.J.; Medeiros, D.T. Characteristics and frequency of detection of fecal Listeria monocytogenes shed by livestock, wildlife, and humans. Can. J. Microbiol. 2007, 53, 1158–1167. [Google Scholar] [CrossRef]
- Chen, Y.; Ross, W.H.; Whiting, R.C.; Stelten, A.V.; Nightingale, K.K.; Wiedmann, M.; Scott, V.N. Variation in Listeria monocytogenes dose responses in relation to subtypes encoding a full-length or truncated internalin A. Appl. Environ. Microbiol. 2011, 77, 1171. [Google Scholar] [CrossRef] [PubMed]
- Van Stelten, A.; Nightingale, K.K. Development and Implementation of a Multiplex Single-Nucleotide Polymorphism Genotyping Assay for Detection of Virulence-Attenuating Mutations in the Listeria monocytogenes Virulence-Associated Gene inlA. Appl. Environ. Microbiol. 2008, 74, 7365–7375. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Molina, A.; Cobo-Díaz, J.F.; López, M.; Prieto, M.; de Toro, M.; Alvarez-Ordóez, A. Unraveling the emergence and population diversity of Listeria monocytogenes in a newly built meat facility through whole genome sequencing. Int. J. Food Microbiol. 2021, 340, 109043. [Google Scholar] [CrossRef] [PubMed]
- Orsi, R.H.; Bakker, H.C.D.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. IJMM 2011, 301, 79–96. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.F.; Ferreira, V.; Magalhaes, R.; Almeida, G.; Alves, A.; Teixeira, P. Detection of premature stop codons leading to truncated internalin A among food and clinical strains of Listeria monocytogenes. Food Microbiol. 2017, 63, 6–11. [Google Scholar] [CrossRef]
- Chen, M.; Cheng, J.; Zhang, J.; Chen, Y.; Wu, Q. Isolation, Potential Virulence, and Population Diversity of Listeria monocytogenes From Meat and Meat Products in China. Front. Microbiol. 2019, 10, 946. [Google Scholar] [CrossRef]
- Song, Z.; Ji, S.; Wang, Y.; Luo, L.; Wang, Y.; Mao, P.; Li, L.; Jiang, H.; Ye, C. The population structure and genetic diversity of Listeria monocytogenes ST9 strains based on genomic analysis. Front. Microbiol. 2022, 13, 982220. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Z.; Li, Z.; Xu, C.; Wang, H.; Yang, R.; Liu, L. Clinical and Genomic Characteristics of a Clinical Listeria monocytogenes ST120 Isolate Recovered from a Pregnant Woman. Infect. Drug Resist. 2024, 17, 229–237. [Google Scholar] [CrossRef]
- Muchaamba, F.; Eshwar, A.; Stevens, M.; Stephan, R.; Tasara, T. Different Shades of Listeria monocytogenes: Strain, Serotype, and Lineage-Based Variability in Virulence and Stress Tolerance Profiles. Front. Microbiol. 2021, 12, 792162. [Google Scholar] [CrossRef]
- Gyanwali, G.C.; Herath, T.U.B.; Gianfelice, A.; Ireton, K. Listeria monocytogenes Co-Opts the Host Exocyst Complex to Promote Internalin A-Mediated Entry. Infect. Immun. 2022, 90, e0032622. [Google Scholar] [CrossRef]
- Parra-Flores, J.; Holý, O.; Bustamante, F.; Lepuschitz, S.; Pietzka, A.; Contreras-Fernández, A.; Castillo, C.; Ovalle, C.; Alarcón-Lavín, M.; Cruz-Córdova, A.; et al. Virulence and Antibiotic Resistance Genes in Listeria monocytogenes Strains Isolated from Ready-to-Eat Foods in Chile. Front. Microbiol. 2021, 12, 796040. [Google Scholar] [CrossRef]
- Agostinho Davanzo, E.; Dos Santos, R.; Castro, V.; Palma, J.; Pribul, B.; Dallago, B.; Fuga, B.; Medeiros, M.; Titze de Almeida, S.; da Costa, H.; et al. Molecular characterization of Salmonella spp. and Listeria monocytogenes strains from biofilms in cattle and poultry slaughterhouses located in the federal District and State of Goiás, Brazil. PLoS ONE 2021, 16, e0259687. [Google Scholar] [CrossRef]
- Cruz, C.; Pitman, A.; Harrow, S.; Fletcher, G. Listeria monocytogenes associated with New Zealand seafood production and clinical cases: Unique sequence types, truncated InlA, and attenuated invasiveness. Appl. Environ. Microbiol. 2014, 80, 1489–1497. [Google Scholar] [CrossRef]
- Li, L.L.; Ji, S.S.; Wang, Y.; Mao, P.; Chen, J.N.; Liu, L.Y.; Ye, C.Y. Genetic polymorphism of inlA in Listeria monocytogenes. Chin. J. Zoonoses 2023, 39, 240–248. (In Chinese) [Google Scholar] [CrossRef]
- Magagna, G.; Gori, M.; Russini, V.; De Angelis, V.; Spinelli, E.; Filipello, V.; Tranquillo, V.; De Marchis, M.; Bossù, T.; Fappani, C.; et al. Evaluation of the Virulence Potential of Listeria monocytogenes through the Characterization of the Truncated Forms of Internalin A. Int. J. Mol. Sci. 2023, 24, 10141. [Google Scholar] [CrossRef]
- Ji, S.; Song, Z.; Luo, L.; Wang, Y.; Li, L.; Mao, P.; Ye, C.; Wang, Y. Whole-genome sequencing reveals genomic characterization of Listeria monocytogenes from food in China. Front. Microbiol. 2022, 13, 1049843. [Google Scholar] [CrossRef]
- Medeiros, M.; Castro, V.; Mota, A.; Pereira, M.G.; Santana, A.P. Assessment of Internalin A Gene Sequences and Cell Adhesion and Invasion Capacity of Listeria monocytogenes Strains Isolated from Foods of Animal and Related Origins. Foodborne Pathog. Dis. 2020, 18. [Google Scholar] [CrossRef]
- Dellafiora, L.; Filipello, V.; Dall’Asta, C.; Finazzi, G.; Galaverna, G.; Losio, M.N. A Structural Study on the Listeria monocytogenes Internalin A—Human E-cadherin Interaction: A Molecular Tool to Investigate the Effects of Missense Mutations. Toxins 2020, 12, 60. [Google Scholar] [CrossRef]
- Schmitz-Esser, S.; Anast, J.M.; Cortes, B.W. A Large-Scale Sequencing-Based Survey of Plasmids in Listeria monocytogenes Reveals Global Dissemination of Plasmids. Front. Microbiol. 2021, 12, 653155. [Google Scholar] [CrossRef]
- Gelbíčová, T.; Koláčková, I.; Pantůček, R.; Karpíšková, R. A novel mutation leading to a premature stop codon in inlA of Listeria monocytogenes isolated from neonatal listeriosis. New Microbiol. 2015, 38, 293–296. [Google Scholar]
- Kanki, M.; Naruse, H.; Taguchi, M.; Kumeda, Y. Characterization of specific alleles in InlA and PrfA of Listeria monocytogenes isolated from foods in Osaka, Japan and their ability to invade Caco-2 cells. Int. J. Food Microbiol. 2015, 211, 18–22. [Google Scholar] [CrossRef]
- Mao, P.; Wang, Y.; Gan, L.; Liu, L.; Chen, J.; Li, L.; Sun, H.; Luo, X.; Ye, C. Large-scale genetic analysis and biological traits of two SigB factors in Listeria monocytogenes: Lineage correlations and differential functions. Front. Microbiol. 2023, 14, 1268709. [Google Scholar] [CrossRef]
- Schubert, W.D.; Urbanke, C.; Ziehm, T.; Beier, V.; Heinz, D.W. Structure of internalin, a major invasion protein of Listeria monocytogenes, in complex with its human receptor E-cadherin. Cell 2002, 111, 825–836. [Google Scholar] [CrossRef]
- Lecuit, M.; Ohayon, H.; Braun, L.; Mengaud, J.; Cossart, P. Internalin of Listeria monocytogenes with an intact leucine-rich repeat region is sufficient to promote internalization. Infect. Immun. 1997, 65, 5309–5319. [Google Scholar] [CrossRef]
- Osek, J.; Lachtara, B.; Wieczorek, K. How This Pathogen Survives in Food-Production Environments? Front. Microbiol. 2022, 13, 866462. [Google Scholar] [CrossRef]
- Liu, X.; Chen, W.; Fang, Z.; Yu, Y.; Bi, J.; Wang, J.; Dong, Q.; Zhang, H. Persistence of Listeria monocytogenes ST5 in Ready-to-Eat Food Processing Environment. Foods 2022, 11, 2561. [Google Scholar] [CrossRef]
- Frédérique, P.; Federica, P.; Laurent, G.; Alex, L.; Alessandra, D.C.; Gerardo, M. Listeria monocytogenes sequence Types 121 and 14 Repeatedly Isolated Within One Year of Sampling in a Rabbit Meat Processing Plant: Persistence and Ecophysiology. Front. Microbiol. 2018, 9, 596. [Google Scholar]
- Koutsoumanis, K.; Allende, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; De Cesare, A.; Herman, L.; Hilbert, F.; Lindqvist, R.; Nauta, M.; et al. Persistence of microbiological hazards in food and feed production and processing environments. EFSA J. Eur. Food Saf. Auth. 2024, 22, e8521. [Google Scholar] [CrossRef]
- Mao, P.; Wang, Y.; Li, L.; Ji, S.; Li, P.; Liu, L.; Chen, J.; Sun, H.; Luo, X.; Ye, C. ListeriaThe Isolation, Genetic Analysis and Biofilm Characteristics of spp. from the Marine Environment in China. Microorganisms 2023, 11, 2166. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, L.; Ji, S.; Li, Q.; Wang, H.; Zhang, Z.; Mao, P.; Sun, H.; Li, L.; Wang, Y.; et al. Dissecting Listeria monocytogenes Persistent Contamination in a Retail Market Using Whole-Genome Sequencing. Microbiol. Spectr. 2022, 10, e0018522. [Google Scholar] [CrossRef]
- Lisa, G.; Parker, C.T.; Liang, A.S.; Samarpita, W.; Romanolo, K.F.; Raymond, S. The Majority of Genotypes of the Virulence Gene inlA Are Intact among Natural Watershed Isolates of Listeria monocytogenes from the Central California Coast. PLoS ONE 2016, 11, e0167566. [Google Scholar]
- Ireton, K.; Mortuza, R.; Gyanwali, G.; Gianfelice, A.; Hussain, M. Role of internalin proteins in the pathogenesis of Listeria monocytogenes. Mol. Microbiol. 2021, 116, 1407–1419. [Google Scholar] [CrossRef]
- Gómez Borrego, J.; Torrent Burgas, M. Analysis of Host-Bacteria Protein Interactions Reveals Conserved Domains and Motifs That Mediate Fundamental Infection Pathways. Int. J. Mol. Sci. 2022, 23, 11489. [Google Scholar] [CrossRef]
- Jackson, B.; Salter, M.; Tarr, C.; Conrad, A.; Harvey, E.; Steinbock, L.; Saupe, A.; Sorenson, A.; Katz, L.; Stroika, S.; et al. Notes from the field: Listeriosis associated with stone fruit—United States, 2014. MMWR. Morb. Mortal. Wkly. Rep. 2015, 64, 282–283. [Google Scholar]
- Maurella, C.; Gallina, S.; Ru, G.; Adriano, D.; Decastelli, L. Outbreak of febrile gastroenteritis caused by Listeria monocytogenes 1/2a in sliced cold beef ham, Italy, May 2016. Euro. Surveill. 2018, 23, 17-00155. [Google Scholar] [CrossRef] [PubMed]
- Acciari, V.; Iannetti, L.; Gattuso, A.; Sonnessa, M.; Scavia, G.; Montagna, C.; Addante, N.; Torresi, M.; Zocchi, L.; Scattolini, S.; et al. Tracing sources of Listeria contamination in traditional Italian cheese associated with a US outbreak: Investigations in Italy. Epidemiol. Infect. 2016, 144, 2719–2727. [Google Scholar] [CrossRef]
- Thomas, J.; Govender, N.; Mccarthy, K.M.; Erasmus, L.K.; Blumberg, L.H. Outbreak of Listeriosis in South Africa Associated With Processed Meat. New Engl. J. Med. 2020, 382, 632–643. [Google Scholar] [CrossRef]
- Chen, Y.; Strain, E.A.; Allard, M.; Brown, E.W. Genome sequences of Listeria monocytogenes strains J1816 and J1-220, associated with human outbreaks. J. Bacteriol. 2011, 193, 3424–3425. [Google Scholar] [CrossRef]
- Kovacevic, J.; Arguedas-Villa, C.; Wozniak, A.; Tasara, T.; Allen, K. Examination of food chain-derived Listeria monocytogenes strains of different serotypes reveals considerable diversity in inlA genotypes, mutability, and adaptation to cold temperatures. Appl. Environ. Microbiol. 2013, 79, 1915–1922. [Google Scholar] [CrossRef]
- Pirone-Davies, C.; Chen, Y.; Pightling, A.; Ryan, G.; Wang, Y.; Yao, K.; Hoffmann, M.; Allard, M. Genes significantly associated with lineage II food isolates of Listeria monocytogenes. BMC Genom. 2018, 19, 708. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Pouillot, R.; Dennis, S.; Brown, E.W. Genetic diversity and profiles of genes associated with virulence and stress resistance among isolates from the 2010–2013 interagency Listeria monocytogenes market basket survey. PLoS ONE 2020, 15, e0231393. [Google Scholar] [CrossRef]
- Magagna, G.; Finazzi, G.; Filipello, V. inlANewly Designed Primers for the Sequencing of the Gene of Lineage I and II Isolates. Int. J. Mol. Sci. 2022, 23, 14106. [Google Scholar] [CrossRef]
- Holch, A.; Ingmer, H.; Licht, T.R.; Gram, L. Listeria monocytogenes strains encoding premature stop codons in inlA invade mice and guinea pig fetuses in orally dosed dams. J. Med. Microbiol. 2013, 62, 1799–1806. [Google Scholar] [CrossRef]
- Sévellec, Y.; Torresi, M.; Félix, B.; Palma, F.; Roussel, S. First Report on the Finding of Listeria monocytogenes ST121 Strain in a Dolphin Brain. Pathogens 2020, 9, 802. [Google Scholar] [CrossRef]
- Thalacker-Mercer, A.R.; Riddle, E.; Barre, L. Protein and amino acids for skeletal muscle health in aging. Adv. Food Nutr. Res. 2020, 91, 29–64. [Google Scholar]
- Sechrest, E.; Ma, X.; Cahill, M.; Barbera, R.; Wang, Y.; Deng, W. Structural and functional rescue of cones carrying the most common cone opsin C203R missense mutation. JCI Insight 2023, 23, e172834. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).