


Endophytes and Plant Extracts as Potential Antimicrobial Agents against Candidatus Liberibacter Asiaticus, Causal Agent of Huanglongbing


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Endophyte Isolation
2.2. Bacterial Strains and Culture Conditions
2.3. Dual Culture Assay
2.4. Agar-Well Diffusion Assay
2.5. Leaf-Disk Assay
2.6. Psyllid Homogenate Assay
2.7. Identification of CLas Antagonists
3. Results
3.1. Dual Culture Assay
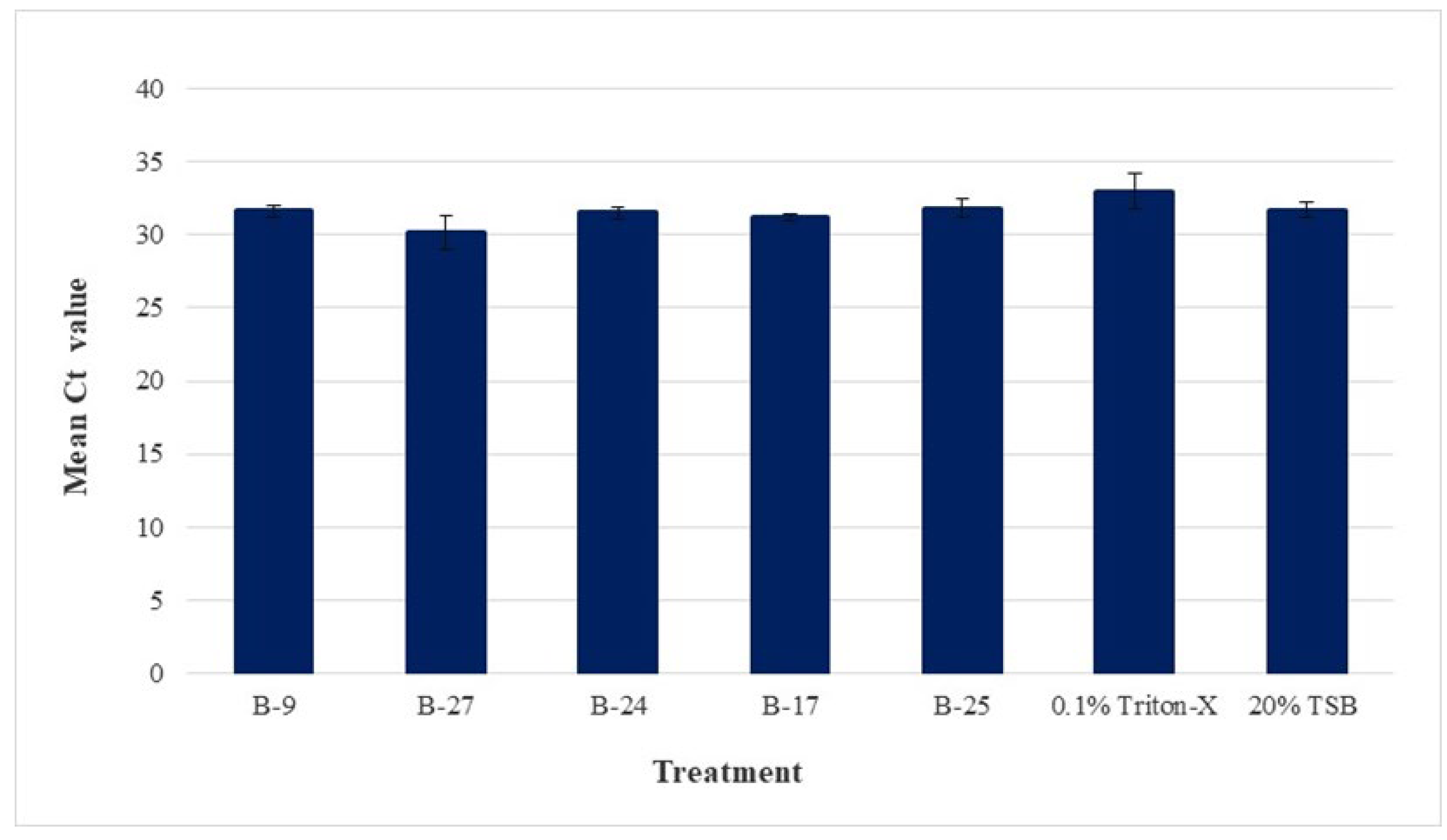
3.2. Agar-Well Diffusion Assay
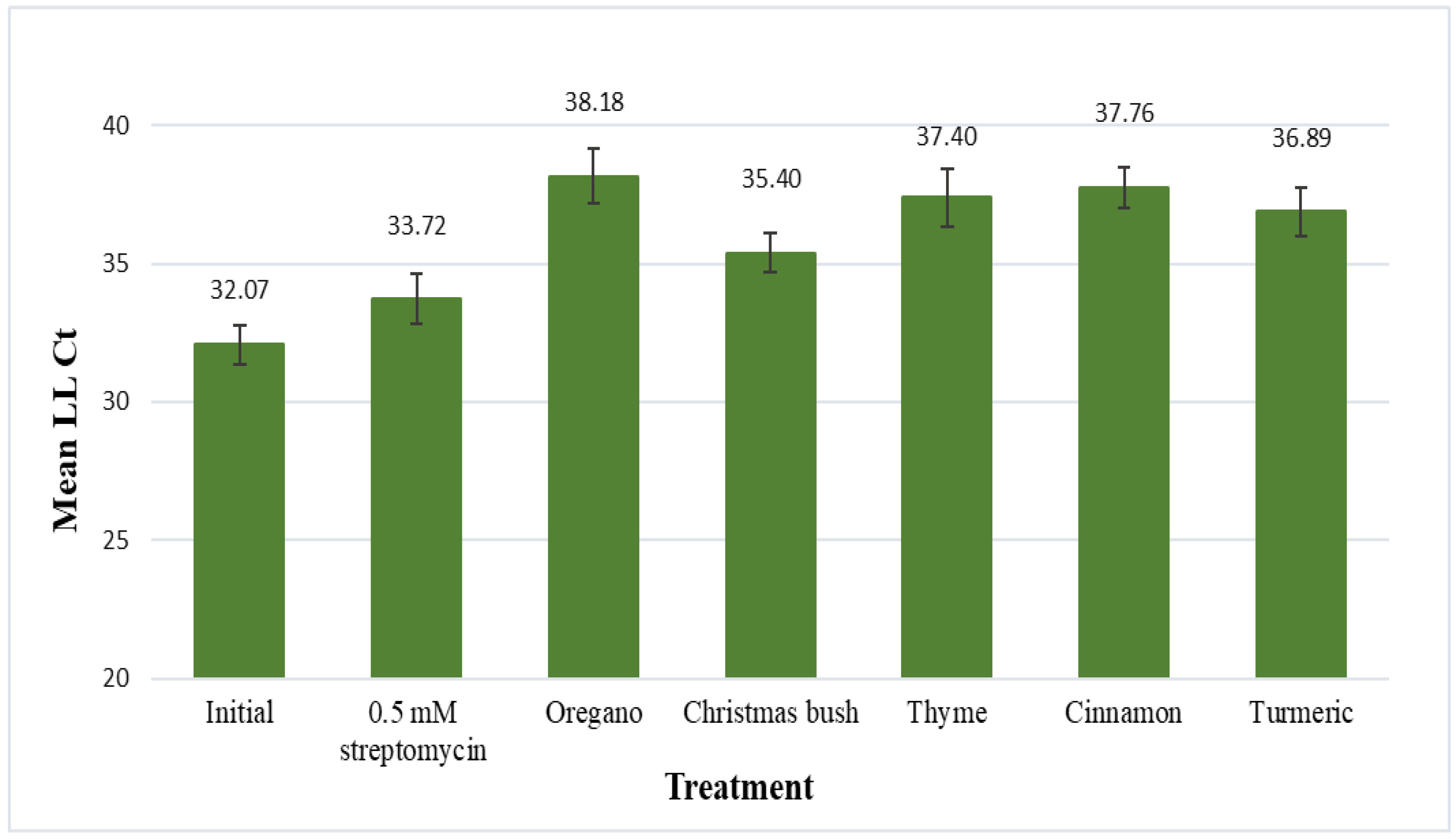
3.3. Leaf-Disk Assay
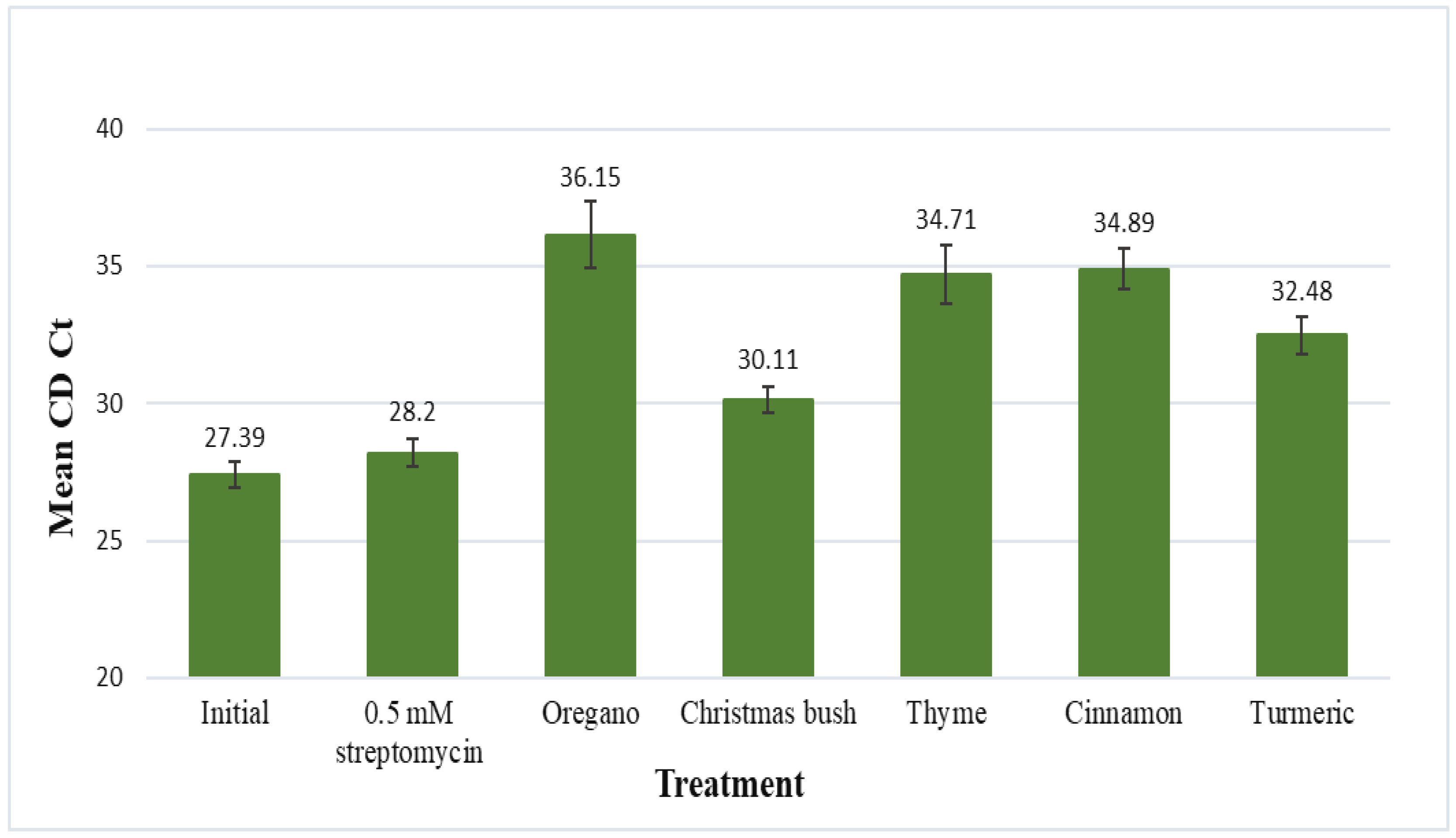
3.4. Psyllid Homogenate Assay
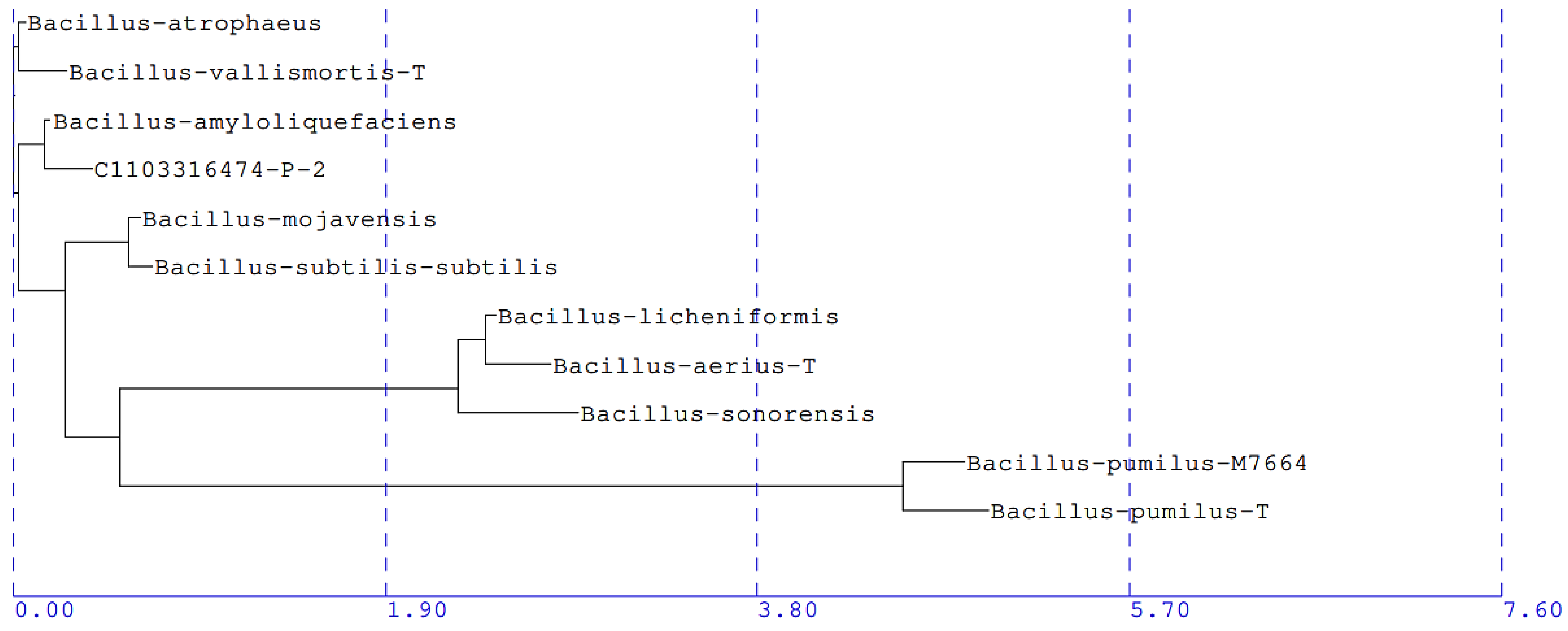
3.5. Identification of CLas Antagonists
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bové, J.M. Huanglongbing: A Destructive, Newly-Emerging, Century-Old Disease of Citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar] [CrossRef]
- Zhang, M.; Guo, Y.; Powell, C.A.; Duan, Y. Antimicrobial Compounds to Combat Citrus Huanglongbing. J. Citrus. Pathol. 2014, 1. [Google Scholar] [CrossRef]
- Miles, G.P.; Stover, E.; Ramadugu, C.; Keremane, M.L.; Lee, R.F. Apparent Tolerance to Huanglongbing in Citrus and Citrus-Related Germplasm. HortScience 2017, 52, 31–39. [Google Scholar] [CrossRef]
- Jagoueix, S.; Bove, J.-M.; Garnier, M. The Phloem-Limited Bacterium of Greening Disease of Citrus Is a Member of the Subdivision of the Proteobacteria. Int. J. Syst. Bacteriol. 1994, 44, 379–386. [Google Scholar] [CrossRef]
- Stover, E.; Gmitter, F.G.; Grosser, J.; Baldwin, E.; Wu, G.A.; Bai, J.; Wang, Y.; Chaires, P.; Motamayor, J.C. Rationale for Reconsidering Current Regulations Restricting Use of Hybrids in Orange Juice. Hortic. Res. 2020, 7, 38. [Google Scholar] [CrossRef]
- Zhang, M.; Powell, C.A.; Guo, Y.; Benyon, L.; Duan, Y. Characterization of the Microbial Community Structure in Candidatus Liberibacter Asiaticus-Infected Citrus Plants Treated with Antibiotics in the Field. BMC Microbiol. 2013, 13, 112. [Google Scholar] [CrossRef]
- Gottwald, T.R. Current Epidemiological Understanding of Citrus Huanglongbing. Annu. Rev. Phytopathol. 2010, 48, 119–139. [Google Scholar] [CrossRef]
- National Research Council. Strategic Planning for the Florida Citrus Industry: Addressing Citrus Greening Disease; National Academies Press: Washington, DC, USA, 2010. [Google Scholar]
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and Management of Asian Citrus Psyllid, Vector of the Huanglongbing Pathogens. Annu. Rev. Entomol. 2013, 58, 413–432. [Google Scholar] [CrossRef]
- Blaustein, R.A.; Lorca, G.L.; Teplitski, M. Challenges for Managing Candidatus Liberibacter Spp. (Huanglongbing Disease Pathogen): Current Control Measures and Future Directions. Phytopathology 2018, 108, 424–435. [Google Scholar] [CrossRef]
- Munir, S.; He, P.; Wu, Y.; He, P.; Khan, S.; Huang, M.; Cui, W.; He, P.; He, Y. Huanglongbing Control: Perhaps the End of the Beginning. Microb. Ecol. 2018, 76, 192–204. [Google Scholar] [CrossRef]
- Al-Rimawi, F.; Hijaz, F.; Nehela, Y.; Batuman, O.; Killiny, N. Uptake, Translocation, and Stability of Oxytetracycline and Streptomycin in Citrus Plants. Antibiotics 2019, 8, 196. [Google Scholar] [CrossRef] [PubMed]
- Brown, C. To Combat Citrus Greening, Farmers Are Spraying Antibiotics on Their Trees. Available online: https://thecounter.org/citrus-greening-florida-farmers-medically-important-antibiotics/ (accessed on 24 May 2022).
- Blacutt, A.; Ginnan, N.; Dang, T.; Bodaghi, S.; Vidalakis, G.; Ruegger, P.; Peacock, B.; Viravathana, P.; Vieira, F.C.; Drozd, C.; et al. An in Vitro Pipeline for Screening and Selection of Citrus-Associated Microbiota with Potential Anti-”Candidatus Liberibacter Asiaticus” Properties. Appl. Environ. Microbiol. 2020, 86, e02883-19. [Google Scholar] [CrossRef] [PubMed]
- Neff, E. Injectable Oxytetracycline Registered—Citrus Industry. Available online: https://citrusindustry.net/2022/11/02/injectable-oxytetracycline-registered/ (accessed on 24 May 2023).
- Fletcher, J.; Wayadanda, A. Fastidious Vascular-Colonizing Bacteria. Plant Health Instr. 2002. [Google Scholar] [CrossRef]
- Trivedi, P.; Trivedi, C.; Grinyer, J.; Anderson, I.C.; Singh, B.K. Harnessing Host-Vector Microbiome for Sustainable Plant Disease Management of Phloem-Limited Bacteria. Front. Plant Sci. 2016, 7, 1423. [Google Scholar] [CrossRef] [PubMed]
- Michaud, J.P. Biological Control of Asian Citrus Psyllid, Diaphorina Citri (Hemiptera: Psyllidae) in Florida: A Preliminary Report. Entomol. News Phila. 2002, 113, 216–222. [Google Scholar]
- Li, S.; Wu, F.; Duan, Y.; Singerman, A.; Guan, Z. Citrus Greening: Management Strategies and Their Economic Impact. HortScience 2020, 55, 604–612. [Google Scholar] [CrossRef]
- Djeddour, D.; Rwomushana, I. The Asian Citrus Greening Disease (Huanglongbing): Evidence Note on Invasiveness and Potential Economic Impacts for East Africa. View Project. CABI Work. Pap. 2021. [Google Scholar] [CrossRef]
- Wang, N.; Pierson, E.A.; Setubal, J.C.; Xu, J.; Levy, J.G.; Zhang, Y.; Li, J.; Rangel, L.T.; Martins, J. The Candidatus Liberibacter–Host Interface: Insights into Pathogenesis Mechanisms and Disease Control. Annu. Rev. Phytopathol. 2017, 55, 451–482. [Google Scholar] [CrossRef]
- Hu, J.; Wang, N. Evaluation of the Spatiotemporal Dynamics of Oxytetracycline and Its Control Effect Against Citrus Huanglongbing via Trunk Injection. Phytopathology 2016, 106, 1495–1503. [Google Scholar] [CrossRef]
- Ho, M.Y.; Chung, W.C.; Huang, H.C.; Chung, W.H.; Chung, W.H. Identification of Endophytic Fungi of Medicinal Herbs of Lauraceae and Rutaceae with Antimicrobial Property. Taiwania 2012, 57, 229–241. [Google Scholar]
- Petrini, O.; Sieber, T.N.; Toti, L.; Viret, O. Ecology, Metabolite Production, and Substrate Utilization in Endophytic Fungi. Nat. Toxins 1993, 1, 185–196. [Google Scholar] [CrossRef]
- Biba, V.S.; Amily, A.; Sangeetha, S.; Remani, P. Anticancer, Antioxidant and Antimicrobial Activity of Annonaceae Family. World J. Pharm. Pharm. Sci. 2014, 3, 1595–1604. [Google Scholar]
- Kamat, S.; Dixit, R.; Kumari, M. Endophytic Microbiome in Bioactive Compound Production and Plant Disease Management. Microb. Biocontrol: Food Secur. Post Harvest. Manag. 2022, 2, 79–128. [Google Scholar] [CrossRef]
- Munir, S.; Li, Y.; He, P.; Huang, M.; He, P.; Cui, W.; Wu, Y.; He, Y. Core Endophyte Communities of Different Citrus Varieties from Citrus Growing Regions in China. Sci. Rep. 2020, 10, 3648. [Google Scholar] [CrossRef] [PubMed]
- Araújo, W.L.; Maccheroni, W.; Barroso, P.; Araújo, W.L.; Aguilar-Vildoso, C.I.; Barroso, P.A.V.; Saridakis, H.O.; Lúcio De Azevedo, J. Variability and Interactions between Endophytic Bacteria and Fungi Isolated from Leaf Tissues of Citrus Rootstocks. Artic. Can. J. Microbiol. 2001, 47, 229–236. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, C.C. Endophytes: A Potential Resource for Biosynthesis, Biotransformation, and Biodegradation. Ann. Microbiol. 2011, 61, 207–215. [Google Scholar] [CrossRef]
- Anjum, N.; Chandra, R. Endophytic Bacteria: Optimizaton of Isolation Procedure from Various Medicinal Plants and Their Preliminary Characterization. Asian J. Pharm. Clin. Res. 2015, 8, 233–238. [Google Scholar]
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A.; Nautiyal, A. Antimicrobial Resistance and the Alternative Resources with Special Emphasis on Plant-Based Antimicrobials—A Review. Plants 2017, 6, 16. [Google Scholar] [CrossRef]
- Raut, D.; Aruna, K. Antimicrobial Activity of Stevia Rebaudiana against Antibiotic Resistant ESBL Producing Uropathogens and Evaluation of Its Antioxidant Activity. Int. J. Adv. Res. Biol. Sci. 2017, 4, 110–118. [Google Scholar] [CrossRef]
- Gupta, P.D.; Birdi, T.J. Development of Botanicals to Combat Antibiotic Resistance. J. Ayurveda Integr. Med. 2017, 4, 110–118. [Google Scholar] [CrossRef]
- Menpara, D.; Chanda, S. Endophytic Bacteria-Unexplored Reservoir of Antimicrobials for Combating Microbial Pathogens 2013. Available online: https://pdfs.semanticscholar.org/95c0/d95a3a4f84a70e3dc1a5ee0bdeb2fe3c3b4b.pdf (accessed on 16 April 2019).
- Eljounaidi, K.; Lee, S.K.; Bae, H. Bacterial Endophytes as Potential Biocontrol Agents of Vascular Wilt Diseases—Review and Future Prospects. Biological. Control. 2016, 103, 62–68. [Google Scholar] [CrossRef]
- Cazorla, F.M.; Mercado-Blanco, J. Biological Control of Tree and Woody Plant Diseases: An Impossible Task? BioControl 2016, 61, 233–242. [Google Scholar] [CrossRef]
- Kandel, S.L.; Joubert, P.M.; Doty, S.L. Bacterial Endophyte Colonization and Distribution within Plants. Microorganisms 2017, 5, 77. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Gmitter, F. What Can We Learn from HLB Survivor Trees? Citrus Ind. 2014, 16–17. Available online: https://swfrec.ifas.ufl.edu/hlb/database/pdf/2_Wang_14.pdf (accessed on 26 April 2023).
- England, G.K. Further Observations of Parson Brown and Other “Surviving” Citrus. Proc. Fla. State Hort. Soc. 2016, 129, 48–50. Available online: https://www.cabdirect.org/cabdirect/abstract/20193451437 (accessed on 24 May 2022).
- Ginnan, N.A.; Dang, T.; Bodaghi, S.; Ruegger, P.M.; McCollum, G.; England, G.; Vidalakis, G.; Borneman, J.; Rolshausen, P.E.; Caroline Roper, M. Disease-Induced Microbial Shifts in Citrus Indicate Microbiome-Derived Responses to Huanglongbing across the Disease Severity Spectrum. Phytobiomes. J. 2020, 4, 375–387. [Google Scholar] [CrossRef]
- Riera, N.; Handique, U.; Zhang, Y.; Dewdney, M.M.; Wang, N. Characterization of Antimicrobial-Producing Beneficial Bacteria Isolated from Huanglongbing Escape Citrus Trees. Front. Microbiol. 2017, 8, 2415. [Google Scholar] [CrossRef]
- Sagaram, U.S.; Deangelis, K.M.; Trivedi, P.; Andersen, G.L.; Lu, S.E.; Wang, N. Bacterial Diversity Analysis of Huanglongbing Pathogen-Infected Citrus, Using PhyloChip Arrays and 16S RRNA Gene Clone Library Sequencing. Appl. Environ. Microbiol. 2009, 75, 1566–1574. [Google Scholar] [CrossRef]
- Trivedi, P.; Spann, T.; Wang, N. Isolation and Characterization of Beneficial Bacteria Associated with Citrus Roots in Florida. Microb. Ecol. 2011, 62, 324–336. [Google Scholar] [CrossRef]
- Trivedi, P.; Duan, Y.; Wang, N. Huanglongbing, a Systemic Disease, Restructures the Bacterial Community Associated with Citrus Roots. Appl. Environ. Microbiol. 2010, 76, 3427–3436. [Google Scholar] [CrossRef]
- Ding, C.; He, J. Effect of Antibiotics in the Environment on Microbial Populations. Appl. Microbiol. Biotechnol. 2010, 87, 925–941. [Google Scholar] [CrossRef] [PubMed]
- Kalia, A.; Gosal, S.K. Effect of Pesticide Application on Soil Microorganisms. Arch. Agron. Soil Sci. 2011, 57, 569–596. [Google Scholar] [CrossRef]
- Seneviratne, G. Collapse of Beneficial Microbial Communities and Deterioration of Soil Health: A Cause for Reduced Crop Productivity. Curr. Sci. 2009, 96, 633. [Google Scholar]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–Microbiome Interactions: From Community Assembly to Plant Health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, J.; Berg, G.; Schulz, B. Isolation Procedures for Endophytic Microorganisms. In Microbial Root Endophytes Soil Biology; Springer: Berlin/Heidelberg, Germany, 2006; Volume 9. [Google Scholar] [CrossRef]
- Shetty, K.G.; Rivadeneira, D.V.; Jayachandran, K.; Walker, D.M. Erratum to: Isolation and Molecular Characterization of the Fungal Endophytic Microbiome from Conventionally and Organically Grown Avocado Trees in South Florida (Mycol Progress, 10.1007/S11557-016-1219-3). Mycol. Prog. 2016, 15, 987–989. [Google Scholar] [CrossRef]
- Fagen, J.R.; Leonard, M.T.; McCullough, C.M.; Edirisinghe, J.N.; Henry, C.S.; Davis, M.J.; Triplett, E.W. Comparative Genomics of Cultured and Uncultured Strains Suggests Genes Essential for Free-Living Growth of Liberibacter. PLoS ONE 2014, 9, e84469. [Google Scholar] [CrossRef]
- Vignesh, M.; Shankar, S.R.M.; MubarakAli, D.; Hari, B.N.V. A Novel Rhizospheric Bacterium: Bacillus Velezensis NKMV-3 as a Biocontrol Agent Against Alternaria Leaf Blight in Tomato. Appl. Biochem. Biotechnol. 2022, 194, 1–17. [Google Scholar] [CrossRef]
- Duan, Y.; Zhou, L.; Hall, D.G.; Li, W.; Doddapaneni, H.; Lin, H.; Liu, L.; Vahling, C.M.; Gabriel, D.W.; Williams, K.P.; et al. Complete Genome Sequence of Citrus Huanglongbing Bacterium, “Candidatus Liberibacter Asiaticus” Obtained Through Metagenomics. Mol. Plant-Microbe Interact. 2009, 22, 1011–1020. [Google Scholar] [CrossRef]
- Stover, E.D.; Stange, R.R.; McCollum, T.G.; Jaynes, J.; Irey, M.; Mirkov, E. Screening Antimicrobial Peptides In Vitro for Use in Developing Transgenic Citrus Resistant to Huanglongbing and Citrus Canker. J. Am. Soc. Hortic. Sci. 2013, 138, 142–148. [Google Scholar] [CrossRef]
- Fagen, J.R.; Leonard, M.T.; Coyle, J.F.; McCullough, C.M.; Davis-Richardson, A.G.; Davis, M.J.; Triplett, E.W. Liberibacter Crescens Gen. Nov., Sp. Nov., the First Cultured Member of the Genus Liberibacter. Int. J. Syst. Evol. Microbiol. 2014, 64, 2461–2466. [Google Scholar] [CrossRef]
- Valgas, C.; Machado De Souza, S.; Smânia, E.F.A.; Smânia, A. Screening Methods to Determine Antibacterial Activity of Natural Products. Braz. J. Microbiol. 2007, 38, 369–380. [Google Scholar] [CrossRef]
- Etxeberria, E.; Gonzalez, P.; Singerman, A.; Ebert, T. An Improved Method to Track Changes of Candidatus Liberibacter Asiaticus Titer in HLB-Affected Citrus Trees. HortScience 2019, 54, 1357–1360. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Krystel, J.; Shi, Q.; Shaw, J.; Gupta, G.; Hall, D.; Stover, E. An in Vitro Protocol for Rapidly Assessing the Effects of Antimicrobial Compounds on the Unculturable Bacterial Plant Pathogen, Candidatus Liberibacter Asiaticus. Plant Methods 2019, 15, 85. [Google Scholar] [CrossRef]
- Nocker, A.; Cheung, C.-Y.; Camper, A.K. Comparison of Propidium Monoazide with Ethidium Monoazide for Differentiation of Live vs. Dead Bacteria by Selective Removal of DNA from Dead Cells. J. Microbiol. Methods 2006, 67, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Tian, Z.; He, H.; Long, C.A.; Jiang, F. Bacillus Species as Potential Biocontrol Agents against Citrus Diseases. Biological. Control. 2020, 151, 104419. [Google Scholar] [CrossRef]
- Zhang, M.; Guo, Y.; Powell, C.A.; Doud, M.S.; Yang, C.; Duan, Y. Effective Antibiotics against “Candidatus Liberibacter Asiaticus” in HLB-Affected Citrus Plants Identified via the Graft-Based Evaluation. PLoS ONE 2014, 9, 17–21. [Google Scholar] [CrossRef]
- Hu, J.; Akula, N.; Wang, N. Development of a Microemulsion Formulation for Antimicrobial SecA Inhibitors. PLoS ONE 2016, 11, e0150433. [Google Scholar] [CrossRef] [PubMed]
- Leonard, M.T.; Fagen, J.R.; Davis-Richardson, A.G.; Davis, M.J.; Triplett, E.W. Complete Genome Sequence of Liberibacter Crescens BT-1. Stand Genomic. Sci. 2012, 7, 271–283. [Google Scholar] [CrossRef]
- Krystel, J.; Liu, H.; Hartung, J.; Stover, E. Novel Plantibodies Show Promise to Protect Citrus from Greening Disease. J. Am. Soc. Hortic. Sci. 2021, 146, 377–386. [Google Scholar] [CrossRef]
- Steinbuch, K.B.; Fridman, M. Mechanisms of Resistance to Membrane-Disrupting Antibiotics in Gram-Positive and Gram-Negative Bacteria. Medchemcomm 2016, 7, 86–102. [Google Scholar] [CrossRef]
- Hurdle, J.G.; O’Neill, A.J.; Chopra, I.; Lee, R.E. Targeting Bacterial Membrane Function: An Underexploited Mechanism for Treating Persistent Infections. Nat. Rev. Microbiol. 2011, 9, 62. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Oh, H.-S.; Park, S.-C.; Chun, J. Towards a Taxonomic Coherence between Average Nucleotide Identity and 16S RRNA Gene Sequence Similarity for Species Demarcation of Prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulou, A.; Theologidis, I.; Varympopi, A.; Papafotis, D.; Mermigka, G.; Tzima, A.; Panopoulos, N.J.; Skandalis, N. Shifting Perspectives of Translational Research in Bio-Bactericides: Reviewing the Bacillus Amyloliquefaciens Paradigm. Biology 2021, 10, 1202. [Google Scholar] [CrossRef] [PubMed]
- Nan, J.; Zhang, S.; Jiang, L. Antibacterial Potential of Bacillus Amyloliquefaciens GJ1 against Citrus Huanglongbing. Plants 2021, 10, 261. [Google Scholar] [CrossRef]
- Luo, L.; Zhao, C.; Wang, E.; Raza, A.; Yin, C. Bacillus Amyloliquefaciens as an Excellent Agent for Biofertilizer and Biocontrol in Agriculture: An Overview for Its Mechanisms. Microbiol. Res. 2022, 259, 127016. [Google Scholar] [CrossRef]
- Tang, J.; Ding, Y.; Nan, J.; Yang, X.; Liang, S.U.N.; Zhao, X.; Jiang, L. Transcriptome Sequencing and ITRAQ Reveal the Detoxification Mechanism of Bacillus GJ1, a Potential Biocontrol Agent for Huanglongbing. PLoS ONE 2018, 13, e0200427. [Google Scholar] [CrossRef]
- Alvarez, F.; Anastassiadou, M.; Arena, M.; Auteri, D.; Brancato, A.; Bura, L.; Carrasco Cabrera, L.; Castoldi, A.F.; Chaideftou, E.; Chiusolo, A.; et al. Peer Review of the Pesticide Risk Assessment of the Active Substance Bacillus Amyloliquefaciens Strain IT-45. EFSA J. 2021, 19, 6594. [Google Scholar] [CrossRef]
- Dimopoulou, A.; Theologidis, I.; Benaki, D.; Koukounia, M.; Zervakou, A.; Tzima, A.; Diallinas, G.; Hatzinikolaou, D.G.; Skandalis, N. Direct Antibiotic Activity of Bacillibactin Broadens the Biocontrol Range of Bacillus Amyloliquefaciens MBI600. Msphere 2021, 6, e0037621. [Google Scholar] [CrossRef]
- Reva, O.N.; Dixelius, C.; Meijer, J.; Priest, F.G. Taxonomic Characterization and Plant Colonizing Abilities of Some Bacteria Related to Bacillus Amyloliquefaciens and Bacillus Subtilis. FEMS Microbiol. Ecol. 2004, 48, 249–259. [Google Scholar] [CrossRef]
- Porwal, S.; Lal, S.; Cheema, S.; Kalia, V.C. Phylogeny in Aid of the Present and Novel Microbial Lineages: Diversity in Bacillus. PLoS ONE 2009, 4, e4438. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Target | Type | Sequence |
|---|---|---|---|
| LLjs-F | CLas 16S rDNA | Forward | 5′-GCATGGAAACGTGTGCTAATAC-3′ |
| LLjs-R | CLas 16S rDNA | Reverse | 5′-AGGCTCTTACCCTACCAACTA-3′ |
| LLjs-Probe | CLas 16S rDNA | Probe | 5′/65-FAM/TTGGAGAGA/ZEN/GATGAGCCTGCGTTG/3IABkFQ/-3′ |
| CDjs-F | citrus dehydrogenase | Forward | 5′-GCAGCAGTTTCTTTGTCCTTATC-3′ |
| CDjs-R | citrus dehydrogenase | Reverse | 5′-CCAAGGAGAAGAAGGGCATATT-3′ |
| CDjs-Probe | citrus dehydrogenase | Probe | 5′-/5Cy5/ACCACCCAAAGTCTGAGGACGAAA/3IAbRQSp/-3′ |
| Isolate Number | Inhibition of S. meliloti in Dual Culture Assay | Inhibition of L. crescens by CFCS in Diffusion Assay |
|---|---|---|
| B-1 | + | − |
| B-2 | + | − |
| B-3 | + | − |
| B-4 | + | − |
| B-5 | + | − |
| B-6 | ++ | + |
| B-7 | + | − |
| B-8 | + | − |
| B-9 | ++ | ++ |
| B-10 | + | − |
| B-11 | + | − |
| B-12 | + | − |
| B-13 | + | + |
| B-14 | + | − |
| B-15 | + | − |
| B-16 | + | − |
| B-17 | ++ | ++ |
| B-18 | + | − |
| B-19 | + | − |
| B-20 | + | + |
| B-21 | + | − |
| B-22 | ++ | + |
| B-23 | + | − |
| B-24 | ++ | ++ |
| B-25 | ++ | ++ |
| B-26 | + | − |
| B-27 | ++ | ++ |
| B-28 to B-179 | − | N/A |
| F-1 to F-163 | − | N/A |
| Plant Species | Concentration | |||
|---|---|---|---|---|
| Scientific Name | Common Name | 100% | 10% | 1% |
| Artemisia annua | Wormwood | ++ | + | − |
| Cryptolepis sanguinolenta | Nibima | + | − | − |
| Alchornea cordifolia | Christmas bush | ++ | + | + |
| Bidens pilosa | Cobbler’s pegs | − | − | − |
| Dispsacus fullonum | Teasel | − | − | − |
| Uncaria tomentosa | Cat’s claw | − | − | − |
| Origanum vulgare | Oregano | ++ | ++ | + |
| Thymus vulgaris | Thyme | ++ | + | + |
| Cinnamomum aromaticum | Cinnamon | ++ | ++ | + |
| Curcuma longa | Turmeric | ++ | ++ | + |
| Ocimum tenuiflorum | Basil | + | − | − |
| Usnea longissima | Bearded lichen | + | + | − |
| Otoba parvifolia | Banderol | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dominguez, J.; Jayachandran, K.; Stover, E.; Krystel, J.; Shetty, K.G. Endophytes and Plant Extracts as Potential Antimicrobial Agents against Candidatus Liberibacter Asiaticus, Causal Agent of Huanglongbing. Microorganisms 2023, 11, 1529. https://doi.org/10.3390/microorganisms11061529
Dominguez J, Jayachandran K, Stover E, Krystel J, Shetty KG. Endophytes and Plant Extracts as Potential Antimicrobial Agents against Candidatus Liberibacter Asiaticus, Causal Agent of Huanglongbing. Microorganisms. 2023; 11(6):1529. https://doi.org/10.3390/microorganisms11061529
Chicago/Turabian StyleDominguez, Jessica, Krishnaswamy Jayachandran, Ed Stover, Joseph Krystel, and Kateel G. Shetty. 2023. "Endophytes and Plant Extracts as Potential Antimicrobial Agents against Candidatus Liberibacter Asiaticus, Causal Agent of Huanglongbing" Microorganisms 11, no. 6: 1529. https://doi.org/10.3390/microorganisms11061529
APA StyleDominguez, J., Jayachandran, K., Stover, E., Krystel, J., & Shetty, K. G. (2023). Endophytes and Plant Extracts as Potential Antimicrobial Agents against Candidatus Liberibacter Asiaticus, Causal Agent of Huanglongbing. Microorganisms, 11(6), 1529. https://doi.org/10.3390/microorganisms11061529