

Predicting the Growth of Vibrio parahaemolyticus in Oysters under Varying Ambient Temperature

Abstract
1. Introduction
2. Materials and Methods
2.1. OLS and LOESS Regression Models for V. parahaemolyticus Growth and Inactivation Processes
2.2. Growth and Inactivation Rates for the Model
2.3. The Model
2.3.1. Model Description, Mathematical Theory, and Assumptions
2.3.2. Model Equations
2.3.3. Model Verification and Evaluation
2.3.4. Modeling Scenarios
2.4. Risk of Illness
2.5. Model Limitations
3. Results
3.1. OLS and LOESS Regression Models for V. parahaemolyticus
3.2. Growth and Inactivation Rates for the Model
3.3. Model Verification and Evaluation
3.4. Modeling Scenarios under Varying Temperature
3.4.1. Simulations 9–11: Summer, Water 30 °C, Air (Max) 40 °C
3.4.2. Simulations 12–14, Summer: Water 25 °C, Air (Max) 32 °C
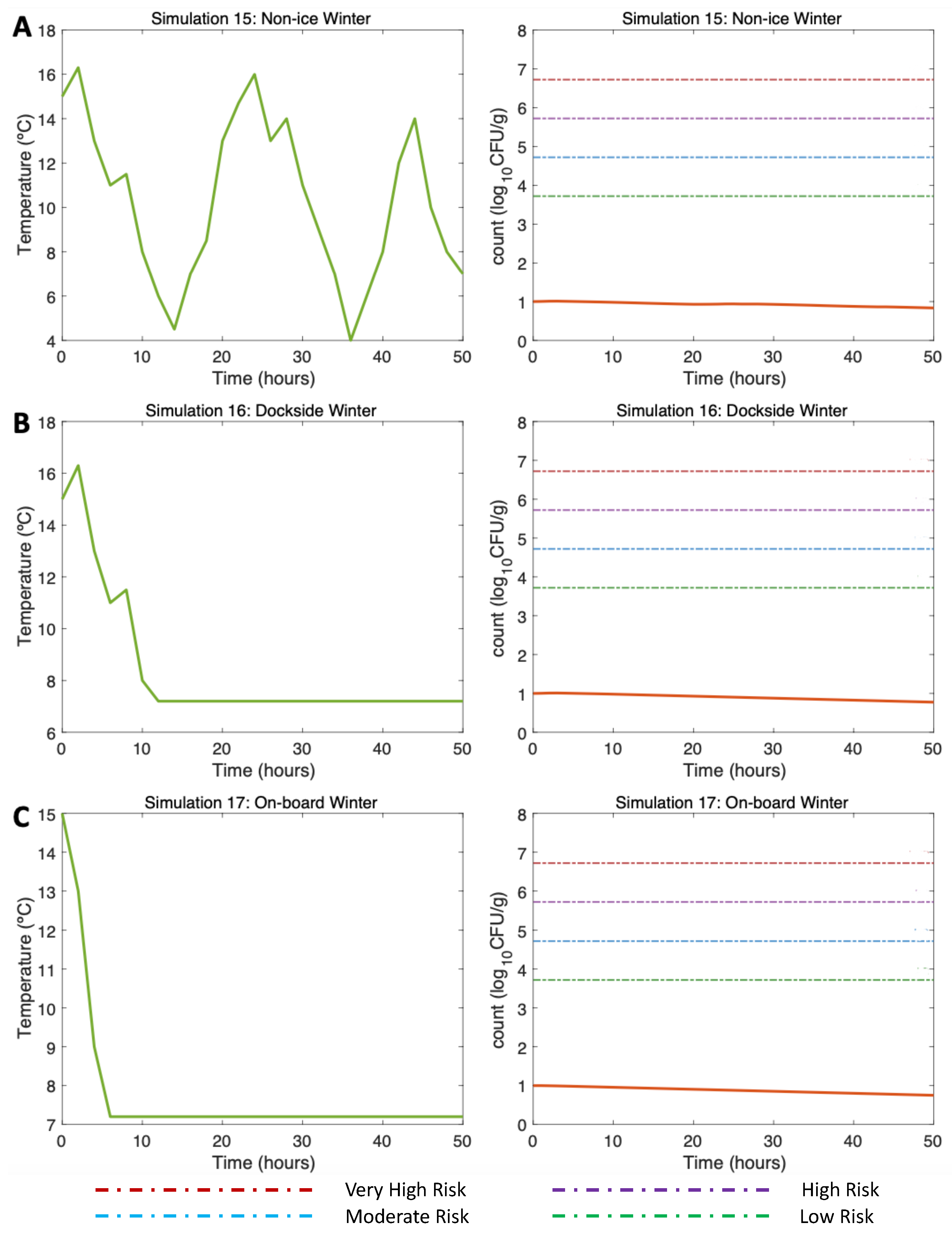
3.4.3. Simulations 15–17: Winter, Water 15 °C Air (Max) 16 °C
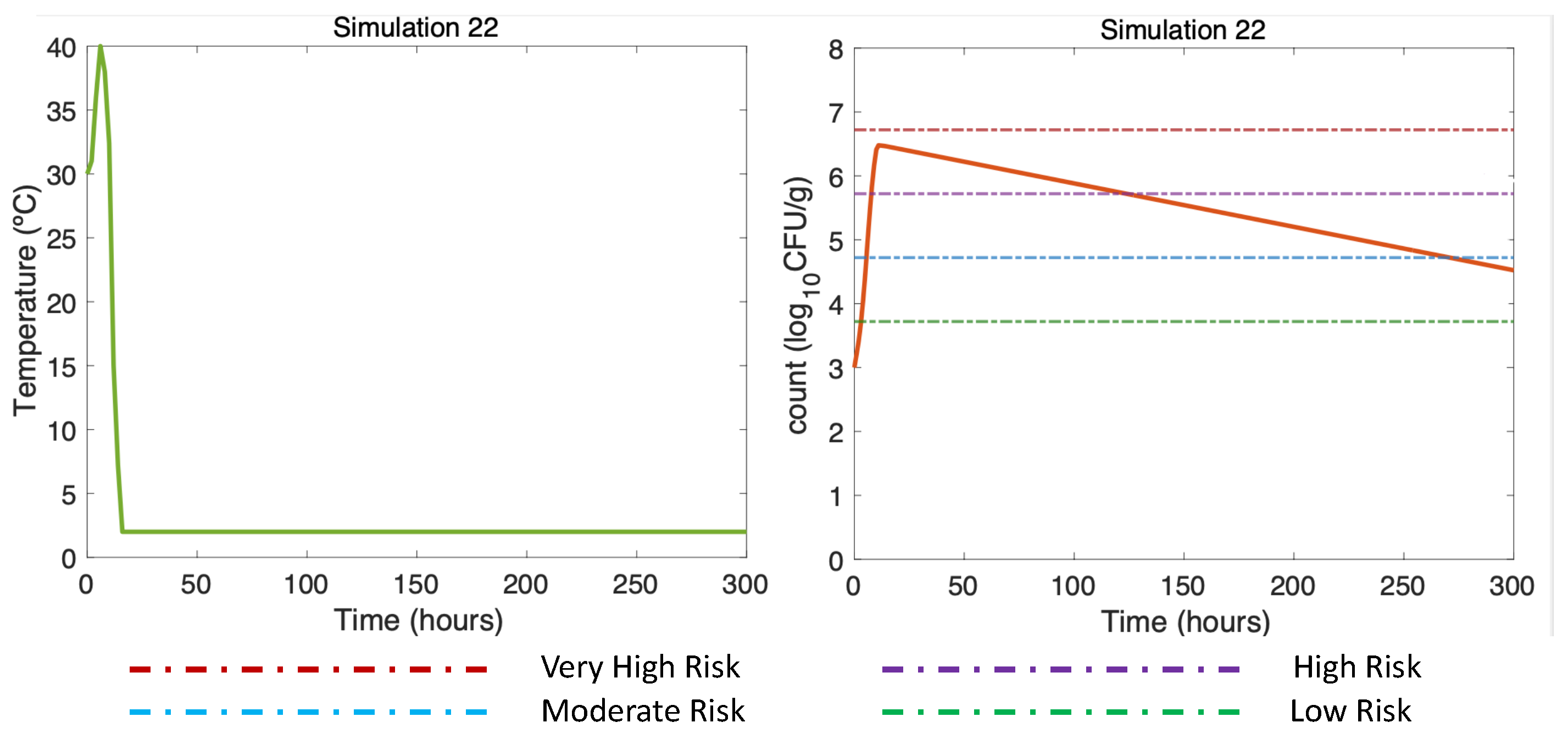
3.4.4. Other Simulations of Interest: Simulations 18–22
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NMFS. Status of the Eastern Oyster, Crassostrea virginica; National Marine Fisheries Service: Silver Spring, MD, USA, 2007. [Google Scholar]
- Su, Y.C.; Liu, C. Vibrio parahaemolyticus: A Concern Seafood Safety. Food Microbiol. 2007, 24, 549–558. [Google Scholar] [CrossRef]
- Odeyemi, O.A. Incidence and prevalence of Vibrio parahaemolyticus Seafood: A Systematic Review and Meta-Analysis. Springerplus 2016, 5, 1–17. [Google Scholar] [CrossRef]
- Joseph, S.W.; Colwell, R.R.; Kaper, J.B. Vibrio parahaemolyticus and related halophilic vibrios. CRC Crit. Rev. Microbiol. 1982, 10, 77–124. [Google Scholar] [CrossRef]
- Jay, J.M.; Loessner, M.J.; Golden, D.A. Foodborne gastroenteritis caused by Vibrio, Yersinia, and Campylobacter species. In Modern Food Microbiology; Food Science Texts Series; Springer: Boston, MA, USA, 2005; pp. 657–678. [Google Scholar]
- Desmarchelier, P.M. Pathogenic vibrios. In Foodborne Microorganisms of Public Health Significance; Australian Institute of Food Science and Technology Incorporated (AIFST Inc.): Marriockville, Australia, 2003; pp. 333–358. [Google Scholar]
- Bruner, D.M.; Huth, W.L.; McEvoy, D.M.; Morgan, O.A. Consumer valuation of food safety: The case of postharvest processed oysters. Agric. Resour. Econ. Rev. 2014, 43, 300–318. [Google Scholar] [CrossRef]
- WHO. Risk Assessment of Vibrio parahaemolyticus in Seafood: Interpretative Summary and Technical Report; World Health Organization: Geneva, Switzerland, 2011.
- CDC. Preliminary FoodNet Data on the incidence of infection with pathogens transmitted commonly through food–10 States, 2008. Centers Dis. Control Prev. MMWR. Morb. Mortal. Wkly. Rep. 2009, 58, 333–337. [Google Scholar]
- Wang, R.; Zhong, Y.; Gu, X.; Yuan, J.; Saeed, A.F.; Wang, S. The pathogenesis, detection, and prevention of Vibrio parahaemolyticus. Front. Microbiol. 2015, 6, 144. [Google Scholar] [CrossRef]
- Shen, X.; Su, Y.C.; Liu, C.; Oscar, T.; DePaola, A. Efficacy of Vibrio parahaemolyticus Depuration Oysters (Crassostrea gigas). Food Microbiol. 2019, 79, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Trinanes, J.; Martinez-Urtaza, J. Future scenarios of risk of Vibrio infections in a warming planet: A global mapping study. Lancet Planet. Health 2021, 5, e426–e435. [Google Scholar] [CrossRef] [PubMed]
- Vezzulli, L.; Grande, C.; Reid, P.C.; Hélaouët, P.; Edwards, M.; Höfle, M.G.; Brettar, I.; Colwell, R.R.; Pruzzo, C. Climate influence on Vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc. Natl. Acad. Sci. USA 2016, 113, E5062–E5071. [Google Scholar] [CrossRef]
- Lydon, K.A.; Farrell-Evans, M.; Jones, J.L. Evaluation of ice slurries as a control for postharvest growth of Vibrio spp. in oysters and potential for filth contamination. J. Food Prot. 2015, 78, 1375–1379. [Google Scholar] [CrossRef]
- Yoon, K.; Min, K.; Jung, Y.; Kwon, K.; Lee, J.; Oh, S. A model of the effect of temperature on the growth of pathogenic and nonpathogenic Vibrio parahaemolyticus isolated from oysters in Korea. Food Microbiol. 2008, 25, 635–641. [Google Scholar] [CrossRef]
- DePaola, A.; Nordstrom, J.L.; Bowers, J.C.; Wells, J.G.; Cook, D.W. Seasonal abundance of total and pathogenic Vibrio parahaemolyticus Alabama Oysters. Appl. Environ. Microbiol. 2003, 69, 1521–1526. [Google Scholar] [CrossRef]
- Ndraha, N.; Hsiao, H.I. The risk assessment of Vibrio parahaemolyticus Raw Oysters Taiwan under the Seasonal Variations, Time Horizons, and Climate Scenarios. Food Control 2019, 102, 188–196. [Google Scholar] [CrossRef]
- Takemura, A.F.; Chien, D.M.; Polz, M.F. Associations and dynamics of Vibrionaceae in the environment, from the genus to the population level. Front. Microbiol. 2014, 5, 38. [Google Scholar] [CrossRef]
- Audemard, C.; Ben-Horin, T.; Kator, H.I.; Reece, K.S. Vibrio vulnificus and Vibrio parahaemolyticus in Oysters under Low Tidal Range Conditions: Is Seawater Analysis Useful for Risk Assessment? Foods 2022, 11, 4065. [Google Scholar] [CrossRef]
- Froelich, B.; Bowen, J.; Gonzalez, R.; Snedeker, A.; Noble, R. Mechanistic and statistical models of total Vibrio abundance in the Neuse River Estuary. Water Res. 2013, 47, 5783–5793. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Piquer, J.; Bowman, J.P.; Ross, T.; Tamplin, M.L. Predictive Models for the Effect of Storage Temperature on Vibrio parahaemolyticus Viability and Counts of Total Viable Bacteria in Pacific Oysters (Crassostrea gigas). Appl. Environ. Microbiol. 2011, 77, 8687–8695. [Google Scholar] [CrossRef]
- FDA. Quantitative Risk Assessment on the Public Health Impact of Pathogenic Vibrio parahaemolyticus, 2005th ed.; U.S. Department of Health and Human Services: Washington, DC, USA, 2005. [Google Scholar]
- Mudoh, M.F.; Parveen, S.; Schwarz, J.; Rippen, T.; Chaudhuri, A. The Effects of Storage Temperature on the Growth of Vibrio parahaemolyticus and Organoleptic Properties in Oysters. Front. Public Health 2014, 2, 45. [Google Scholar] [CrossRef] [PubMed]
- Pruente, V.L.; Jones, J.L.; Steury, T.D.; Walton, W.C. Effects of tumbling, refrigeration and subsequent resubmersion on the abundance of Vibrio vulnificus and Vibrio parahaemolyticus in cultured oysters (Crassostrea virginica). Int. J. Food Microbiol. 2020, 335, 108858. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, S.; Hwang, I.; Yoon, K. Effect of Temperature on Growth of Vibrio Paraphemolyticus Vibrio Vulnificus Flounder, Salmon Sashimi Oyster Meat. Int. J. Environ. Res. Public Health 2012, 9, 4662–4675. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.y.; Fodjo, E.K.; Kong, C.; Gu, R.R.; Han, F.; Shen, X.S. Temperature effect study on growth and survival of pathogenic Vibrio parahaemolyticus Jinjiang Oyster (Crassostrea rivularis) Rapid Count Method. J. Food Qual. 2018, 2018, 2060915. [Google Scholar] [CrossRef]
- Bernhardt, J.R.; Sunday, J.M.; Thompson, P.L.; O’Connor, M.I. Nonlinear averaging of thermal experience predicts population growth rates in a thermally variable environment. Proc. R. Soc. B Biol. Sci. 2018, 285, 20181076. [Google Scholar] [CrossRef]
- Fernandez-Piquer, J.; Bowman, J.P.; Ross, T.; Estrada-Flores, S.; Tamplin, M.L. Preliminary stochastic model for managing Vibrio parahaemolyticus and total viable bacterial counts in a Pacific oyster (Crassostrea gigas) supply chain. J. Food Prot. 2013, 76, 1168–1178. [Google Scholar] [CrossRef]
- Love, D.C.; Kuehl, L.M.; Lane, R.M.; Fry, J.P.; Harding, J.; Davis, B.J.; Clancy, K.; Hudson, B. Performance of cold chains and modeled growth of Vibrio parahaemolyticus for farmed oysters distributed in the United States and internationally. Int. J. Food Microbiol. 2020, 313, 108378. [Google Scholar] [CrossRef]
- Galton, F. Regression towards mediocrity in hereditary stature. J. Anthropol. Inst. Great Br. Irel. 1886, 15, 246–263. [Google Scholar] [CrossRef]
- Cleveland, W.S.; Devlin, S.J. Locally weighted regression: An approach to regression analysis by local fitting. J. Am. Stat. Assoc. 1988, 83, 596–610. [Google Scholar] [CrossRef]
- Ratkowsky, D.A.; Olley, J.; McMeekin, T.; Ball, A. Relationship between temperature and growth rate of bacterial cultures. J. Bacteriol. 1982, 149, 1–5. [Google Scholar] [CrossRef]
- He, F.; Yeung, L.F.; Brown, M. Discrete-time model representations for biochemical pathways. Trends Intell. Syst. Comput. Eng. 2008, 6, 255–271. [Google Scholar]
- Verhulst, P. Deuxième mémoire sur la loi d’accroissement de la population. Nouv. Acad. R. Sci. Lett. Des-Beaux-Arts Belg. 1847, 20, 1–32. [Google Scholar]
- AEMET. Agencia Estatal de Meteorología. Available online: https://www.aemet.es/ (accessed on 23 January 2023).
- NCEI. National Center of Environmental Information NOAA. Available online: https://www.ncei.noaa.gov/ (accessed on 12 October 2022).
- McLaughlin, J.B.; DePaola, A.; Bopp, C.A.; Martinek, K.A.; Napolilli, N.P.; Allison, C.G.; Murray, S.L.; Thompson, E.C.; Bird, M.M.; Middaugh, J.P. Outbreak of Vibrio parahaemolyticus gastroenteritis associated with Alaskan oysters. N. Engl. J. Med. 2005, 353, 1463–1470. [Google Scholar] [CrossRef] [PubMed]
- Levy, S. Warming Trend: How Climate Shapes Vibrio Ecol. Environ. Health Perspect. 2015, 123, A82–A89. [Google Scholar] [CrossRef]
- Sestelo, M.; Villanueva, N.M.; Meira-Machado, L.; Roca-Pardiñas, J. An R Package for Non-parametric Estimation and Inference in Life Sciences. J. Stat. Softw. 2017, 82, 1–27. [Google Scholar] [CrossRef]
- Gajewski, Z.; Stevenson, L.A.; Pike, D.A.; Roznik, E.A.; Alford, R.A.; Johnson, L.R. Predicting the growth of the amphibian chytrid fungus in varying temperature environments. Ecol. Evol. 2021, 11, 17920–17931. [Google Scholar] [CrossRef]
- Fu, S.; Shen, J.; Liu, Y.; Chen, K. A predictive model of Vibrio cholerae for combined temperature and organic nutrient in aquatic environments. J. Appl. Microbiol. 2013, 114, 574–585. [Google Scholar] [CrossRef]
- Ellett, A.N.; Rosales, D.; Jacobs, J.M.; Paranjpye, R.; Parveen, S. Growth Rates of Vibrio parahaemolyticus Sequence Type 36 Strains in Live Oysters and in Culture Medium. Microbiol. Spectr. 2022, 10, e02112-22. [Google Scholar] [CrossRef]
- Chen, P.; Wang, J.J.; Hong, B.; Tan, L.; Yan, J.; Zhang, Z.; Liu, H.; Pan, Y.; Zhao, Y. Characterization of mixed-species biofilm formed by Vibrio parahaemolyticus and Listeria monocytogenes. Front. Microbiol. 2019, 10, 2543. [Google Scholar] [CrossRef]
- Chae, M.; Cheney, D.; Su, Y.C. Temperature effects on the depuration of Vibrio parahaemolyticus Vibrio vulnificus from the American Oyster (Crassostrea virginica). J. Food Sci. 2009, 74, M62–M66. [Google Scholar] [CrossRef]
- Melody, K.P. Two Post-Harvest Treatments for the Reduction of Vibrio vulnificus and Vibrio parahaemolyticus in Eastern Oysters (Crassostrea virginica); LSU Digital Commons; Louisiana State University and Agricultural & Mechanical College ProQuest Dissertations Publishing: Baton Rouge, LA, USA, 2008. [Google Scholar]
- Andrews, L.S.; Park, D.L.; Chen, Y.P. Low temperature pasteurization to reduce the risk of vibrio infections from raw shell-stock oysters. Food Addit. Contam. 2000, 17, 787–791. Available online: http://xxx.lanl.gov/abs/ (accessed on 10 November 2022). [CrossRef]
- Martinez-Urtaza, J.; Bowers, J.C.; Trinanes, J.; DePaola, A. Climate anomalies and the increasing risk of Vibrio parahaemolyticus Vibrio vulnificus Illnesses. Food Res. Int. 2010, 43, 1780–1790. [Google Scholar] [CrossRef]
- Groner, M.L.; Maynard, J.; Breyta, R.; Carnegie, R.B.; Dobson, A.; Friedman, C.S.; Froelich, B.; Garren, M.; Gulland, F.M.; Heron, S.F.; et al. Managing marine disease emergencies in an era of rapid change. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150364. [Google Scholar] [CrossRef]
- Maynard, J.; Van Hooidonk, R.; Harvell, C.D.; Eakin, C.M.; Liu, G.; Willis, B.L.; Williams, G.J.; Groner, M.L.; Dobson, A.; Heron, S.F.; et al. Improving marine disease surveillance through sea temperature monitoring, outlooks and projections. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150208. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Simulation | Scenario | Expected Results |
|---|---|---|
| Simulation 1 | T = 18.4 °C, Vp = 3.4 CFU/g | Logistic (extension) 3.4 to 5.5 CFU/g |
| Simulation 2 | T = 20 °C, Vp = 3.4 CFU/g | Logistic (extension) 3.4 to 5.5 CFU/g |
| Simulation 3 | T = 25.7 °C, Vp = 3.4 CFU/g | Logistic (extension) 3.4 to 7.5 CFU/g |
| Simulation 4 | T = 30.4 °C, Vp = 3.4 CFU/g | Logistic (extension) 3.4 to 6.75 CFU/g |
| Simulation 5 | T = 3.6 °C, Vp = 5.8 CFU/g | Linear 5.8 to 3.0 CFU/g |
| Simulation 6 | T = 6.2 °C, Vp = 5.5 CFU/g | Linear 5.5 to 4.0 CFU/g |
| Simulation 7 | T = 9.6 °C, Vp = 5.1 CFU/g | Linear 5.1 to 3.0 CFU/g |
| Simulation 8 | T = 12.6 °C, Vp = 5.3 CFU/g | Linear 5.3 to 4.0 CFU/g |
| Simulation | Scenario |
|---|---|
| Simulation 9 | NI, Summer, W = 30 °C, A = 40 °C, Vp = 3 CFU/g |
| Simulation 10 | DS, Summer, W = 30 °C, A = 40 °C, Vp = 3 CFU/g |
| Simulation 11 | OB, Summer, W = 30 °C, A = 40 °C, Vp = 3 CFU/g |
| Simulation 12 | NI, Summer, W = 25 °C, A = 32 °C, Vp = 3 CFU/g |
| Simulation 13 | DS, Summer, W = 25 °C, A = 32 °C, Vp = 3 CFU/g |
| Simulation 14 | OB, Summer, W = 25 °C, A = 32 °C, Vp = 3 CFU/g |
| Simulation 15 | NI, Winter, W = 15 °C, A = 16 °C, Vp = 1 CFU/g |
| Simulation 16 | DS, Winter, W = 15 °C, A = 16 °C, Vp = 1 CFU/g |
| Simulation 17 | OB, Winter, W = 15 °C, A = 16 °C, Vp = 1 CFU/g |
| Simulation 18 | DS, BCC Summer, W = 30 °C, A = 32 °C, Vp = 3 CFU/g |
| Simulation 19 | OB, BCC, Summer, W = 30 °C, A = 32 °C, Vp = 3 CFU/g |
| Simulation 20 | DS, BCC, Winter, W = 15 °C, A = 18 °C, Vp = 1 CFU/g |
| Simulation 21 | OB, BCC, Winter, W = 15 °C, A = 18 °C, Vp = 1 CFU/g |
| Simulation 22 | DS, Summer, W = 25 °C, A = 40 °C, Vp = 3 CFU/g |
| CFU per Serving | Log CFU/g | P (G) | Risk (G) | P (S) | Risk (S) |
|---|---|---|---|---|---|
| 1.72 | Extremely low | Extremely low | |||
| 2.72 | Very low | Extremely low | |||
| 3.72 | Low | Extremely low | |||
| 4.72 | Moderate | Extremely low | |||
| 5.72 | High | Very low | |||
| 6.72 | Very high | Very low | |||
| 7.72 | Extremely high | Very low | |||
| 8.72 | Extremely high | Very low |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Vélez, I.; Bidegain, G.; Ben-Horin, T. Predicting the Growth of Vibrio parahaemolyticus in Oysters under Varying Ambient Temperature. Microorganisms 2023, 11, 1169. https://doi.org/10.3390/microorganisms11051169
Fernández-Vélez I, Bidegain G, Ben-Horin T. Predicting the Growth of Vibrio parahaemolyticus in Oysters under Varying Ambient Temperature. Microorganisms. 2023; 11(5):1169. https://doi.org/10.3390/microorganisms11051169
Chicago/Turabian StyleFernández-Vélez, Iker, Gorka Bidegain, and Tal Ben-Horin. 2023. "Predicting the Growth of Vibrio parahaemolyticus in Oysters under Varying Ambient Temperature" Microorganisms 11, no. 5: 1169. https://doi.org/10.3390/microorganisms11051169
APA StyleFernández-Vélez, I., Bidegain, G., & Ben-Horin, T. (2023). Predicting the Growth of Vibrio parahaemolyticus in Oysters under Varying Ambient Temperature. Microorganisms, 11(5), 1169. https://doi.org/10.3390/microorganisms11051169