
A Simple Assay to Assess Salmonella Typhimurium Impact on Performance and Immune Status of Growing Pigs after Different Inoculation Doses

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Housing and Management
2.2. Experimental Design
2.3. Data and Sample Collection
2.4. Statistical Analysis
3. Results
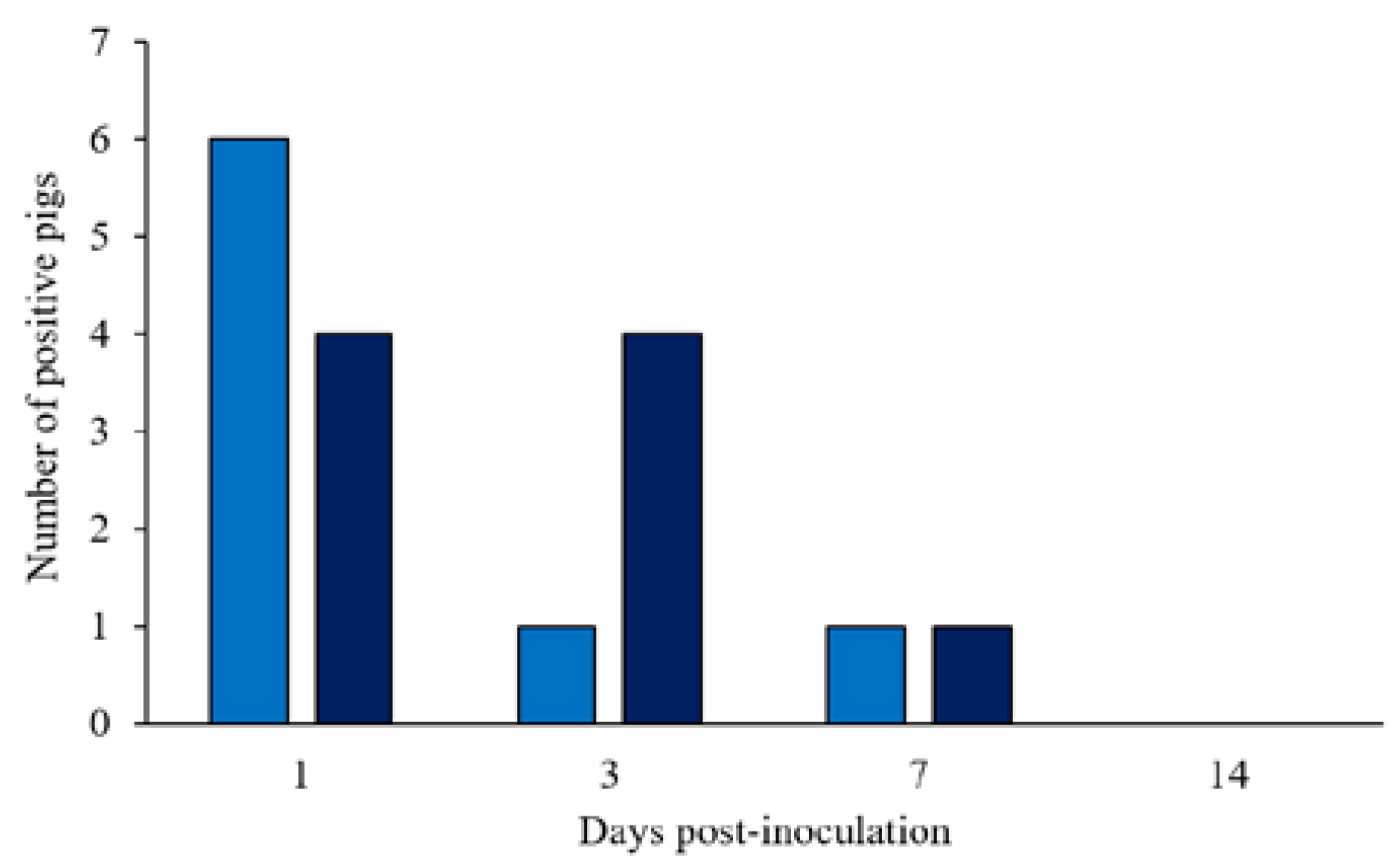
3.1. Salmonella Typhimurium Fecal Shedding, Rectal Temperature, and Fecal Score
3.2. Hematological Parameters
3.3. Growth Performance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campos, P.H.; Floc’h, L.; Noblet, J.; Renaudeau, D. Physiological responses of growing pigs to high ambient temperature and/or inflammatory challenges. Rev. Bras. Zootec. 2017, 46, 537–544. [Google Scholar] [CrossRef]
- Qin, X.; Yang, M.; Cai, H.; Liu, Y.; Gorris, L.; Aslam, M.Z.; Jia, K.; Sun, T.; Wang, X.; Dong, Q. Antibiotic Resistance of Salmonella Typhimurium Monophasic Variant 1,4,[5],12:I:-In China: A Systematic Review and Meta-Analysis. Antibiotics 2022, 11, 532. [Google Scholar] [CrossRef]
- Won, Y.; Kim, S.; Han, J. The protective effect of dietary supplementation of Salmonella-specific bacteriophages in post-weaning piglets challenged with Salmonella typhimurium. J. Adv. Veter. Anim. Res. 2021, 8, 440–447. [Google Scholar] [CrossRef]
- Deane, A.; Murphy, D.; Leonard, F.C.; Byrne, W.; Clegg, T.; Madigan, G.; Griffin, M.; Egan, J.; Prendergast, D.M. Prevalence of Salmonella spp. in slaughter pigs and carcasses in Irish abattoirs and their antimicrobial resistance. Ir. Veter. J. 2022, 75, 4. [Google Scholar] [CrossRef]
- Gomes, V.T.M.; Moreno, L.Z.; Silva, A.P.S.; Thakur, S.; La Ragione, R.M.; Mather, A.E.; Moreno, A.M. Characterization of Salmonella Enterica Contamination in Pork and Poultry Meat from São Paulo/Brazil: Serotypes, Genotypes and Antimicrobial Resistance Profiles. Pathogens 2022, 11, 358. [Google Scholar] [CrossRef]
- Morningstar-Shaw, B.R.; Mackie, T.A.; Barker, D.K.; Palmer, E.A. Salmonella Serotypes Isolated from Animals and Related Sources January; CDC: Ames, IA, USA, 2016; pp. 1–4. [Google Scholar]
- Kempf, F.; Cordoni, G.; Chaussé, A.-M.; Drumo, R.; Brown, H.; Horton, D.L.; Paboeuf, F.; Denis, M.; Velge, P.; La Ragione, R.; et al. Inflammatory Responses Induced by the Monophasic Variant of Salmonella Typhimurium in Pigs Play a Role in the High Shedder Phenotype and Fecal Microbiota Composition. Msystems 2023, 11, e00852-22. [Google Scholar] [CrossRef] [PubMed]
- Larivière-Gauthier, G.; Thibodeau, A.; Yergeau, É.; Fravalo, P. Sows affect their piglets’ faecal microbiota until fattening but not their Salmonella enterica shedding status. Lett. Appl. Microbiol. 2021, 72, 113–120. [Google Scholar] [CrossRef]
- Hankel, J.; Chuppava, B.; Wilke, V.; Hartung, C.B.; Muthukumarasamy, U.; Strowig, T.; Knudsen, K.E.B.; Kamphues, J.; Visscher, C. High Dietary Intake of Rye Affects Porcine Gut Microbiota in a Salmonella Typhimurium Infection Study. Plants 2022, 11, 2232. [Google Scholar] [CrossRef] [PubMed]
- Griffith, R.; Schawrtz, K.; Meyerholz, D. Salmonella. In Diseases of Swine, 1st ed.; Straw, B., Zimmerman, J., D’Allaire, S., Taylor, D., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2006; pp. 739–754. [Google Scholar]
- Rodrigues, L.A.; Wellington, M.O.; González-Vega, J.C.; Htoo, J.K.; van Kessel, A.G.; Columbus, D.A. Functional amino acid supplementation, regardless of dietary protein content, improves growth performance and immune status of weaned pigs challenged with Salmonella Typhimurium. J. Anim. Sci. 2021, 99, 1–13. [Google Scholar] [CrossRef]
- Rostagno, M.H.; Eicher, S.D.; Lay, D.C.J. Immunological, physiological, and behavioral effects of Salmonella enterica carriage and shedding in experimentally infected finishing pigs. Foodborne Path. Dis. 2011, 8, 623–630. [Google Scholar] [CrossRef]
- Nielsen, B.; Baggesen, D.; Bager, F.; Haugegaard, J.; Lind, P. The serological response to Salmonella serovars typhimurium and infantis in experimentally infected pigs. The time course followed with an indirect anti-LPS ELISA and bacteriological examinations. Veter. Microbiol. 1995, 47, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.C.; Rostagno, M.H.; Gardiner, G.E.; Sutton, A.L.; Richert, B.T.; Radcliffe, J.S. Controlling Salmonella infection in weanling pigs through water delivery of direct-fed microbials or organic acids. Part I: Effects on growth performance, microbial populations, and immune status1. J. Anim. Sci. 2012, 90, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Moura, E.; Silva, D.; Turco, C.; Sanches, T.; Storino, G.; Almeida, H.; Mechler-Dreibi, M.; Rabelo, I.; Sonalio, K.; Oliveira, L. Salmonella Bacterin Vaccination Decreases Shedding and Colonization of Salmonella Typhimurium in Pigs. Microorganisms 2021, 9, 1163. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.M.; Wallace, K.P.; Penn, M.J.C.; Petry, A.L.; Broadway, R.; Sanchez, N.C.B.; Carroll, J.A.; Ballou, M.A. A Dose-Response Investigation of a Micronized Porous Ceramic Particle to Improve the Health and Performance of Post-weaned Pigs Infected With Salmonella enterica Serotype Typhimurium. Front. Anim. Sci. 2022, 3, 1–17. [Google Scholar] [CrossRef]
- Balaji, R.; Wright, K.J.; Hill, C.M.; Dritz, S.S.; Knoppel, E.L.; Minton, J.E. Acute phase responses of pigs challenged orally with Salmonella typhimurium. J. Anim. Sci. 2000, 78, 1885–1891. [Google Scholar] [CrossRef]
- Spiehs, M.; Shurson, G.; Johnston, L. Effects of two direct-fed microbials on the ability of pigs to resist an infection with Salmonella enterica serovar Typhimurium. J. Swine Health Prod. 2008, 16, 27–36. [Google Scholar]
- Casey, P.G.; Gardiner, G.E.; Casey, G.; Bradshaw, B.; Lawlor, P.G.; Lynch, P.B.; Leonard, F.C.; Stanton, C.; Ross, R.P.; Fitzgerald, G.F.; et al. A five-strain probiotic combination reduces pathogen shedding and alleviates disease signs in pigs challenged with Salmonella enterica serovar Typhimurium. Appl. Environ. Microbiol. 2007, 73, 1858–1863. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Swine, 11th ed.; National Academies Press: Washington, DC, USA, 2012; p. 420. [Google Scholar]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Wood, R.L.; Rose, R.; Coe, N.E.; Ferris, K.E. Experimental establishment of persistent infection in swine with a zoonotic strain of Salmonella newport. Am. J. Veter. Res. 1991, 52, 813–819. [Google Scholar]
- Oliveira, L.G.; Carvalho, L.F.O.S.; Masson, G.C.I.H.; Feliciano, M.A.R. Experimental Infection by Salmonella enterica ssp. enterica serovar Panama and Tentative of Nose-to-Nose Transmission in Weaned Pigs. Arq. Bras. Med. Vet. Zootec. 2010, 62, 1340–1347. [Google Scholar] [CrossRef]
- Proux, K.; Cariolet, R.; Fravalo, P.; Houdayer, C.; Keranflech, A.; Madec, F. Contamination of pigs by nose-to-nose contact or airborne transmission of Salmonella Typhimurium. Veter. Res. 2001, 32, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.L.; Fraser, N.; Burkey, T.E.; Skjolaas, K.A.; Dritz, S.S.; Johnson Pin, B.J.; Minton, J.E. Oral insulation with Salmonella enterica serovar Typhimurium or Choleraesuis promotes divergent al responses in the somatotropic growth axis of swine. J. Anim. Sci. 2010, 88, 1642–1648. [Google Scholar] [CrossRef] [PubMed]
- Scherer, K.; Szabó, I.; Rösler, U.; Appel, B.; Hensel, A.; Nöckler, K. Time Course of Infection with Salmonella Typhimurium and Its Influence on Fecal Shedding, Distribution in Inner Organs, and Antibody Response in Fattening Pigs. J. Food Prot. 2008, 71, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Szabó, I.; Wieler, L.H.; Tedin, K.; Scharek-Tedin, L.; Taras, D.; Hensel, A.; Appel, B.; Nöckler, K. Influence of a Probiotic Strain of Enterococcus faecium on Salmonella enterica Serovar Typhimurium DT104 Infection in a Porcine Animal Infection Model. Appl. Environ. Microbiol. 2009, 75, 2621–2628. [Google Scholar] [CrossRef]
- Dlabac, V.; Trebichavský, I.; Reháková, Z.; Hofmanová, B.; Splíchal, I.; Cukrowska, B. Pathogenicity and protective effect of rough mutants of Salmonella species in germ-free piglets. Infect. Immun. 1997, 65, 5238–5243. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, C.; Wu, G.; Sun, Y.; Wang, B.; He, B.; Dai, Z.; Wu, Z. Glutamine Enhances Tight Junction Protein Expression and Modulates Corticotropin-Releasing Factor Signaling in the Jejunum of Weanling Piglets. J. Nutr. 2015, 145, 25–31. [Google Scholar] [CrossRef]
- Balsbaugh, R.K.; Curtis, S.E.; Meyer, R.C. Body Weight, Total Body Water and Hematocrit in Diarrheic Piglets. J. Anim. Sci. 1986, 62, 307–314. [Google Scholar] [CrossRef]
- Barba-Vidal, E.; Roll, V.F.B.; Manzanilla, E.G.; Torrente, C.; Muñoz, J.A.M.; Pérez, J.F.; Martín-Orúe, S.M. Blood parameters as biomarkers in a Salmonella spp. disease model of weaning piglets. PLoS ONE 2017, 12, e0186781. [Google Scholar] [CrossRef]
- Smolin, N.; Oleinikova, A.; Brovchenko, I.; Geiger, A.; Winter, R. Properties of Spanning Water Networks at Protein Surfaces. J. Phys. Chem. B 2005, 109, 10995–11005. [Google Scholar] [CrossRef]
- van Milgen, J.; Dourmad, J.Y. Concept and application of ideal protein for pigs. J. Anim. Sci. Biotechnol. 2015, 6, 1–11. [Google Scholar] [CrossRef]
- Adedokun, S.A.; Olojede, O.C. Optimizing Gastrointestinal Integrity in Poultry: The Role of Nutrients and Feed Additives. Front. Veter. Sci. 2018, 5, 348. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Inoculation Level, CFU | SEM | p-Value | ||
|---|---|---|---|---|---|
| 0 (Basal) | 1 × 108 | 1.5 × 108 | |||
| BW, 0 dpi | 27.25 | 27.23 | 27.42 | 1.66 | 0.99 |
| BW, 7 dpi | 32.95 | 31.94 | 31.90 | 1.80 | 0.89 |
| BW, 14 dpi | 38.10 | 37.79 | 37.74 | 2.04 | 0.99 |
| 0 to 7 dpi | |||||
| ADG, kg | 0.81 a | 0.67 b | 0.64 b | 0.03 | <0.01 |
| ADFI, kg | 1.55 | 1.52 | 1.54 | 0.07 | 0.95 |
| G:F, kg/kg | 0.52 a | 0.45 b | 0.42 b | 0.02 | <0.01 |
| 0 to 14 dpi | |||||
| ADG, kg | 0.84 | 0.75 | 0.74 | 0.04 | 0.26 |
| ADFI, kg | 1.66 | 1.63 | 1.68 | 0.07 | 0.86 |
| G:F, kg/kg | 0.46 | 0.47 | 0.44 | 0.01 | 0.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves da Cunha Valini, G.; Arnaut, P.R.; Barbosa, L.G.; de Azevedo, P.H.A.; Melo, A.D.B.; Marçal, D.A.; Campos, P.H.R.F.; Hauschild, L. A Simple Assay to Assess Salmonella Typhimurium Impact on Performance and Immune Status of Growing Pigs after Different Inoculation Doses. Microorganisms 2023, 11, 446. https://doi.org/10.3390/microorganisms11020446
Alves da Cunha Valini G, Arnaut PR, Barbosa LG, de Azevedo PHA, Melo ADB, Marçal DA, Campos PHRF, Hauschild L. A Simple Assay to Assess Salmonella Typhimurium Impact on Performance and Immune Status of Growing Pigs after Different Inoculation Doses. Microorganisms. 2023; 11(2):446. https://doi.org/10.3390/microorganisms11020446
Chicago/Turabian StyleAlves da Cunha Valini, Graziela, Pedro Righetti Arnaut, Larissa Gonçalves Barbosa, Paulo Henrique Amadeu de Azevedo, Antonio Diego Brandão Melo, Danilo Alves Marçal, Paulo Henrique Reis Furtado Campos, and Luciano Hauschild. 2023. "A Simple Assay to Assess Salmonella Typhimurium Impact on Performance and Immune Status of Growing Pigs after Different Inoculation Doses" Microorganisms 11, no. 2: 446. https://doi.org/10.3390/microorganisms11020446
APA StyleAlves da Cunha Valini, G., Arnaut, P. R., Barbosa, L. G., de Azevedo, P. H. A., Melo, A. D. B., Marçal, D. A., Campos, P. H. R. F., & Hauschild, L. (2023). A Simple Assay to Assess Salmonella Typhimurium Impact on Performance and Immune Status of Growing Pigs after Different Inoculation Doses. Microorganisms, 11(2), 446. https://doi.org/10.3390/microorganisms11020446