
Fate of Horizontal-Gene-Transfer Markers and Beta-Lactamase Genes during Thermophilic Composting of Human Excreta

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Composting Trial and Sampling
2.2. DNA Extraction and Purification
2.3. Quantitative RealTime PCR
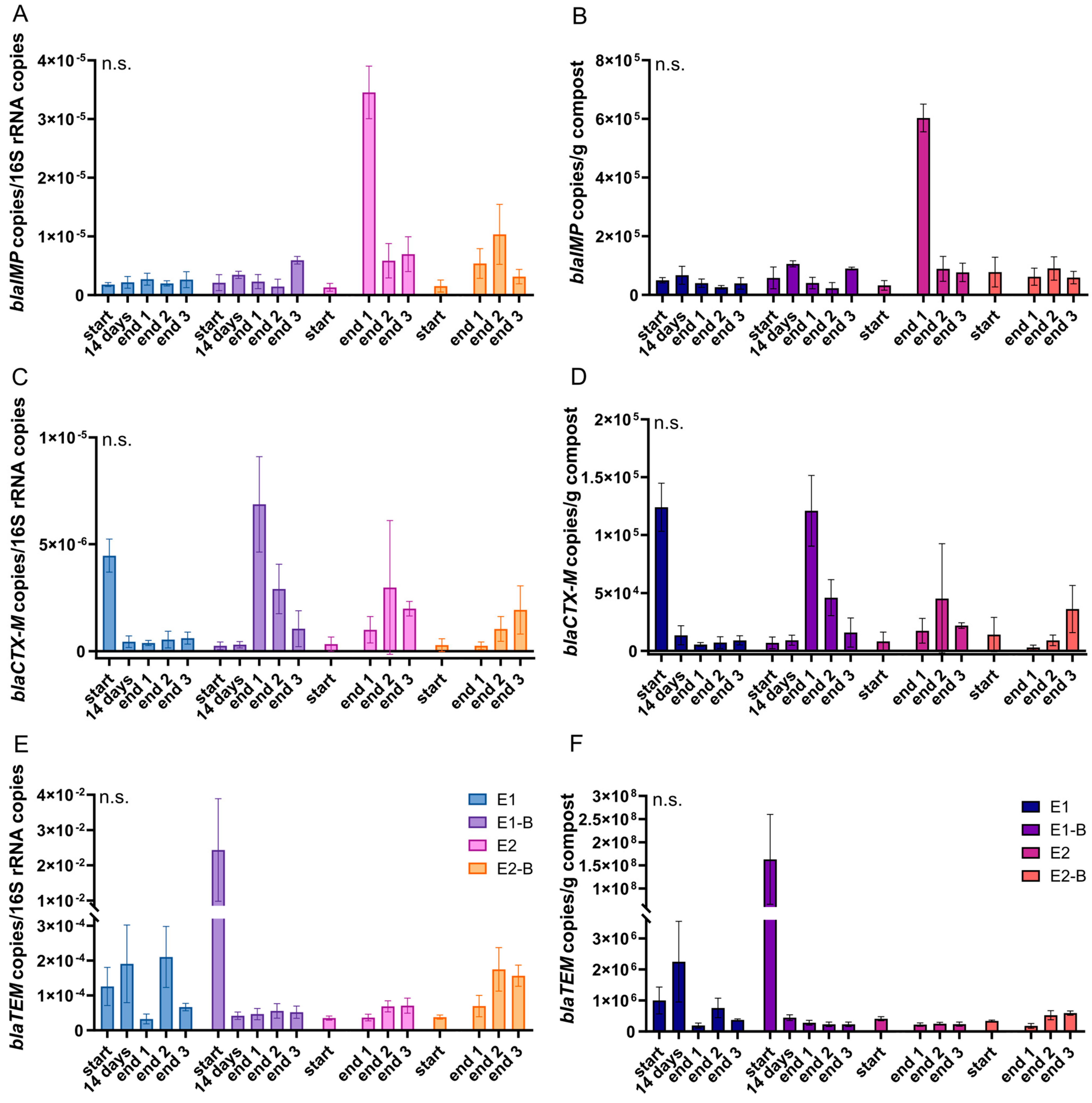
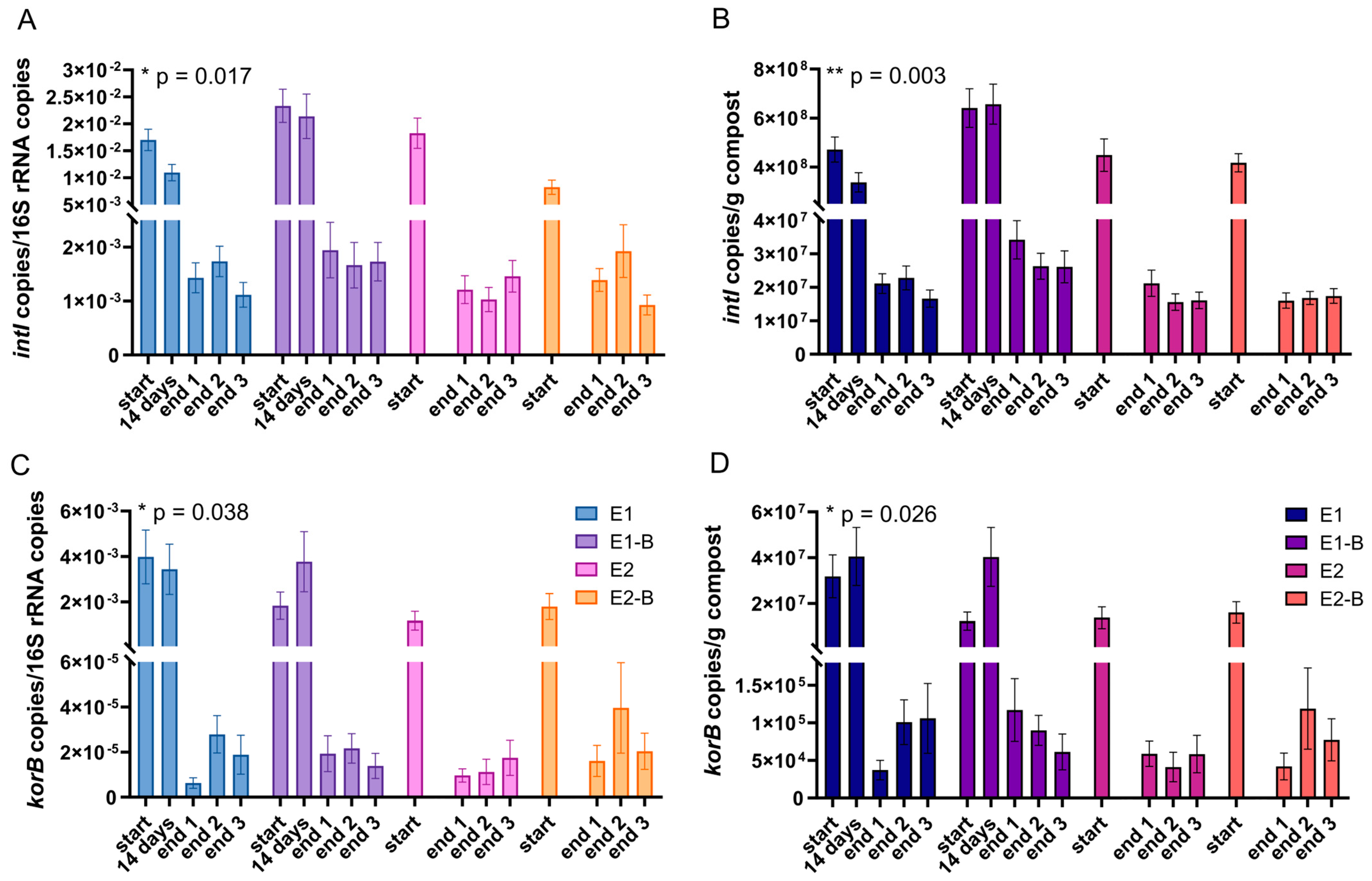
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rose, C.; Parker, A.; Jefferson, B.; Cartmell, E. The characterization of feces and urine: A review of the literature to inform advanced treatment technology. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1827–1879. [Google Scholar] [CrossRef] [PubMed]
- Ryals, R.; Bischak, E.; Porterfield, K.K.; Heisey, S.; Jeliazovski, J.; Kramer, S.; Pierre, S. Toward zero hunger through coupled ecological sanitation-agriculture systems. Front. Sustain. Food Syst. 2021, 5, 716140. [Google Scholar] [CrossRef]
- Bernal, M.P.; Alburquerque, J.A.; Moral, R. Composting of animal manures and chemical criteria for compost maturity assessment. A review. Bioresour. Technol. 2009, 100, 5444–5453. [Google Scholar] [CrossRef] [PubMed]
- Ryckeboer, J.; Mergaert, J.; Vaes, K.; Klammer, S.; De Clercq, D.; Coosemans, J.; Insam, H.; Swings, J. A survey of bacteria and fungi occurring during composting and self-heating processes. Ann. Microbiol. 2003, 53, 349–410. [Google Scholar]
- Godlewska, P.; Schmidt, H.P.; Ok, Y.S.; Oleszczuk, P. Biochar for composting improvement and contaminants reduction. A review. Bioresour. Technol. 2017, 246, 193–202. [Google Scholar] [CrossRef]
- Sanchez-Monedero, M.A.; Cayuela, M.L.; Roig, A.; Jindo, K.; Mondini, C.; Bolan, N. Role of biochar as an additive in organic waste composting. Bioresour. Technol. 2018, 247, 1155–1164. [Google Scholar] [CrossRef]
- Gurtler, J.B.; Doyle, M.P.; Erickson, M.C.; Jiang, X.; Millner, P.; Sharma, M. Composting to inactivate foodborne pathogens for crop soil application: A review. J. Food Prot. 2018, 81, 1821–1837. [Google Scholar] [CrossRef]
- Kelova, M.E.; Ali, A.M.; Eich-Greatorex, S.; Dörsch, P.; Kallenborn, R.; Jenssen, P.D. Small-scale on-site treatment of fecal matter: Comparison of treatments for resource recovery and sanitization. Environ. Sci. Pollut. Res. 2021, 28, 63945–63964. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.M.; Wertheim, H.F.L.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic resistance-the need for global solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef]
- O’Neill, J. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations, London, UK. 2014. Available online: https://wellcomecollection.org/works/rdpck35v (accessed on 8 January 2021).
- Lartigue, M.F.; Poirel, L.; Aubert, D.; Nordmann, P. In vitro analysis of ISEcp1B-mediated mobilization of naturally occurring β-lactamase gene blaCTX-M of Kluyvem ascorbata. Antimicrob. Agents Chemother. 2006, 50, 1282–1286. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, J.; Sui, Q.; Wan, H.; Tong, J.; Chen, M.; Wei, Y.; Wei, D. Effect of red mud addition on tetracycline and copper resistance genes and microbial community during the full scale swine manure composting. Bioresour. Technol. 2016, 216, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Gu, J.; Wang, X.; Zhang, R.; Yin, Y.; Sun, W.; Tuo, X.; Zhang, L. Effects of superabsorbent polymers on the abundances of antibiotic resistance genes, mobile genetic elements, and the bacterial community during swine manure composting. Bioresour. Technol. 2017, 244, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, Y.; Wen, Q. Effects of chlortetracycline on the fate of multi-antibiotic resistance genes and the microbial community during swine manure composting. Environ. Pollut. 2018, 237, 977–987. [Google Scholar] [CrossRef]
- Esperón, F.; Albero, B.; Ugarte-Ruíz, M.; Domínguez, L.; Carballo, M.; Tadeo, J.L.; del Mar Delgado, M.; Moreno, M.Á.; de la Torre, A. Assessing the benefits of composting poultry manure in reducing antimicrobial residues, pathogenic bacteria, and antimicrobial resistance genes: A field-scale study. Environ. Sci. Pollut. Res. 2020, 27, 27738–27749. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Duan, M.; Gu, J.; Zhang, Y.; Qian, X.; Ma, J.; Zhang, R.; Wang, X. Effects of bamboo charcoal on antibiotic resistance genes during chicken manure composting. Ecotoxicol. Environ. Saf. 2017, 140, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Feng, Y.; Liu, Y.; Xue, J.; Li, Z. Dynamics of oxytetracycline, sulfamerazine, and ciprofloxacin and related antibiotic resistance genes during swine manure composting. J. Environ. Manag. 2019, 230, 102–109. [Google Scholar] [CrossRef]
- Duan, M.; Zhang, Y.; Zhou, B.; Wang, Q.; Gu, J.; Liu, G.; Qin, Z.; Li, Z. Changes in antibiotic resistance genes and mobile genetic elements during cattle manure composting after inoculation with Bacillus subtilis. Bioresour. Technol. 2019, 292, 122011. [Google Scholar] [CrossRef]
- Lu, C.; Gu, J.; Wang, X.; Liu, J.; Zhang, K.; Zhang, X.; Zhang, R. Effects of coal gasification slag on antibiotic resistance genes and the bacterial community during swine manure composting. Bioresour. Technol. 2018, 268, 20–27. [Google Scholar] [CrossRef]
- Cui, E.; Wu, Y.; Zuo, Y.; Chen, H. Effect of different biochars on antibiotic resistance genes and bacterial community during chicken manure composting. Bioresour. Technol. 2016, 203, 11–17. [Google Scholar] [CrossRef]
- Cantón, R.; Novais, A.; Valverde, A.; Machado, E.; Peixe, L.; Baquero, F.; Coque, T.M. Prevalence and spread of extended-spectrum β-lactamase-producing Enterobacteriaceae in Europe. Clin. Microbiol. Infect. 2008, 14, 144–153. [Google Scholar] [CrossRef]
- Elshamy, A.A.; Aboshanab, K.M. A review on bacterial resistance to carbapenems: Epidemiology, detection and treatment options. Futur. Sci. OA 2020, 6, FSO438. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, VMBF-0016-2015. [Google Scholar] [CrossRef]
- Watanabe, M.; Iyobe, S.; Inoue, M.; Mitsuhashi, S. Transferable imipenem resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1991, 35, 147–151. [Google Scholar] [CrossRef]
- Logan, L.K.; Weinstein, R.A. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J. Infect. Dis. 2017, 215 (Suppl. S1), S28–S36. [Google Scholar] [CrossRef]
- Karlowsky, J.A.; Lob, S.H.; Kazmierczak, K.M.; Badal, R.E.; Young, K.; Motyl, M.R.; Sahm, D.F. In Vitro Activity of Imipenem against Carbapenemase-Positive Enterobacteriaceae Isolates Collected by the SMART Global Surveillance Program from 2008 to 2014. J. Clin. Microbiol. 2017, 55, 1638–1649. [Google Scholar] [CrossRef]
- Matsumura, Y.; Peirano, G.; Bradford, A.; Motyl, M.R.; DeVinney, R.; Pitout, J.D.D. Genomic characterization of IMP and VIM carbapenemase-encoding transferable plasmids of Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 3034–3038. [Google Scholar] [CrossRef]
- Cohen, R.; Paikin, S.; Rokney, A.; Rubin-Blum, M.; Astrahan, P. Multidrug-resistant enterobacteriaceae in coastal water: An emerging threat. Antimicrob. Resist. Infect. Control 2020, 9, 169. [Google Scholar] [CrossRef]
- Kittinger, C.; Lipp, M.; Folli, B.; Kirschner, A.; Baumert, R.; Galler, H.; Grisold, A.J.; Luxner, J.; Weissenbacher, M.; Farnleitner, A.H.; et al. Enterobacteriaceae Isolated from the River Danube: Antibiotic Resistances, with a Focus on the Presence of ESBL and Carbapenemases. PLoS ONE 2016, 11, e0165820. [Google Scholar] [CrossRef]
- Gorrasi, S.; Pasqualetti, M.; Franzetti, A.; Gonzalez-Martinez, A.; Gonzalez-Lopez, J.; Muñoz-Palazon, B.; Fenice, M. Persistence of Enterobacteriaceae Drawn into a Marine Saltern (Saline di Tarquinia, Italy) from the Adjacent Coastal Zone. Water 2021, 13, 1443. [Google Scholar] [CrossRef]
- Glushakova, A.M.; Kachalkin, V.; Prokof’eva, T.V.; Lysak, L.V. Enterobacteriaceae in soils and atmospheric dust aerosol accumulations of Moscow city. Curr. Res. Microb. Sci. 2022, 3, 100124. [Google Scholar] [CrossRef]
- Cernava, T.; Erlacher, A.; Soh, J.; Sensen, C.W.; Grube, M.; Berg, G. Enterobacteriaceae dominate the core microbiome and contribute to the resistome of arugula (Eruca sativa Mill.). Microbiome 2019, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Pintor-Cora, A.; Álvaro-Llorente, L.; Otero, A.; Rodríguez-Calleja, J.M.; Santos, J.A. Extended-Spectrum Beta-Lactamase-Producing Enterobacteriaceae in Fresh Produce. Foods 2021, 10, 2609. [Google Scholar] [CrossRef] [PubMed]
- Al Atrouni, A.; Joly-Guillou, M.-L.; Hamze, M.; Kempf, M. Reservoirs of Non-baumannii Acinetobacter Species. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Adewoyin, M.A.; Okoh, A.I. The natural environment as a reservoir of pathogenic and non-pathogenic Acinetobacter species. Rev. Environ. Health 2018, 33, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Biehl, L.M.; Schmidt-Hieber, M.; Liss, B.; Cornely, O.A.; Vehreschild, M.J.G.T. Colonization and infection with extended spectrum beta-lactamase producing Enterobacteriaceae in high-risk patients—Review of the literature from a clinical perspective. Crit. Rev. Microbiol. 2016, 42, 1–16. [Google Scholar] [CrossRef]
- Jiang, A.-M.; Liu, N.; Zhao, R.; Zheng, H.-R.; Chen, X.; Fan, C.-X.; Zhang, R.; Zheng, X.-Q.; Fu, X.; Yao, Y.; et al. Clinical outcomes and prognostic factors in bloodstream infections due to extended-spectrum β-lactamase-producing Enterobacteriaceae among patients with malignancy: A meta-analysis. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 53. [Google Scholar] [CrossRef]
- Jiang, A.-M.; Shi, X.; Liu, N.; Gao, H.; Ren, M.-D.; Zheng, X.-Q.; Fu, X.; Liang, X.; Ruan, Z.-P.; Yao, Y.; et al. Nosocomial infections due to multidrug-resistant bacteria in cancer patients: A six-year retrospective study of an oncology Center in Western China. BMC Infect. Dis. 2020, 20, 452. [Google Scholar] [CrossRef]
- Datta, N.; Kontomichalou, P. Penicillinase Synthesis Controlled by Infectious R Factors in Enterobacteriaceae. Nature 1965, 208, 239–241. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. Epidemiology of β-Lactamase-Producing Pathogens. Clin. Microbiol. Rev. 2020, 33, e00047-19. [Google Scholar] [CrossRef]
- Castanheira, M.; Simner, J.; Bradford, P.A. Extended-spectrum β-lactamases: An update on their characteristics, epidemiology and detection. JAC Antimicrobial Resist. 2021, 3, dlab092. [Google Scholar] [CrossRef]
- Ghaly, T.M.; Gillings, M.R.; Penesyan, A.; Qi, Q.; Rajabal, V.; Tetu, S.G. The Natural History of Integrons. Microorganisms 2021, 9, 2212. [Google Scholar] [CrossRef] [PubMed]
- Gillings, M.R.; Gaze, W.H.; Pruden, A.; Smalla, K.; Tiedje, J.M.; Zhu, Y.-G. Using the class 1 integron-integrase gene as a proxy for anthropogenic pollution. ISME J. 2015, 9, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Gillings, M.R. Integrons: Past, present, and future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, M.; Jagura-Burdzy, G. Spread and survival of promiscuous IncP-1 plasmids. Acta Biochim. Pol. 2003, 50, 425–453. [Google Scholar]
- Klümper, U.; Riber, L.; Dechesne, A.; Sannazzarro, A.; Hansen, L.H.; Sørensen, S.J.; Smets, B.F. Broad host range plasmids can invade an unexpectedly diverse fraction of a soil bacterial community. ISME J. 2015, 9, 934–945. [Google Scholar] [CrossRef]
- Willms, I.M.; Yuan, J.; Penone, C.; Goldmann, K.; Vogt, J.; Wubet, T.; Schöning, I.; Schrumpf, M.; Buscot, F.; Nacke, H. Distribution of Medically Relevant Antibiotic Resistance Genes and Mobile Genetic Elements in Soils of Temperate Forests and Grasslands Varying in Land Use. Genes 2020, 11, 150. [Google Scholar] [CrossRef]
- Wolters, B.; Jacquiod, S.; Sørensen, S.J.; Widyasari-Mehta, A.; Bech, T.B.; Kreuzig, R.; Smalla, K. Bulk soil and maize rhizosphere resistance genes, mobile genetic elements and microbial communities are differently impacted by organic and inorganic fertilization. FEMS Microbiol. Ecol. 2018, 94, fiy027. [Google Scholar] [CrossRef]
- Jechalke, S.; Schreiter, S.; Wolters, B.; Dealtry, S.; Heuer, H.; Smalla, K. Widespread dissemination of class 1 integron components in soils and related ecosystems as revealed by cultivation-independent analysis. Front. Microbiol. 2014, 4, 420. [Google Scholar] [CrossRef]
- Shintani, M.; Nour, E.; Elsayed, T.; Blau, K.; Wall, I.; Jechalke, S.; Spröer, C.; Bunk, B.; Overmann, J.; Smalla, K. Plant Species-Dependent Increased Abundance and Diversity of IncP-1 Plasmids in the Rhizosphere: New Insights Into Their Role and Ecology. Front. Microbiol. 2020, 11, 590776. [Google Scholar] [CrossRef]
- Binh, C.T.T.; Heuer, H.; Kaupenjohann, M.; Smalla, K. Piggery manure used for soil fertilization is a reservoir for transferable antibiotic resistance plasmids. FEMS Microbiol. Ecol. 2008, 66, 25–37. [Google Scholar] [CrossRef]
- Schlüter, A.; Szczepanowski, R.; Pühler, A.; Top, E.M. Genomics of IncP-1 antibiotic resistance plasmids isolated from wastewater treatment plants provides evidence for a widely accessible drug resistance gene pool. FEMS Microbiol. Rev. 2007, 31, 449–477. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz Barrón, M.; Merlin, C.; Guilloteau, H.; Montargès-Pelletier, E.; Bellanger, X. Suspended Materials in River Waters Differentially Enrich Class 1 Integron- and IncP-1 Plasmid-Carrying Bacteria in Sediments. Front. Microbiol. 2018, 9, 1443. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.S.; Lázaro, B.; Azevedo, J.S.N.; Henriques, I.; Almeida, A.; Correia, A. New molecular variants of epsilon and beta IncP-1 plasmids are present in estuarine waters. Plasmid 2012, 67, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Shintani, M.; Takahashi, Y.; Yamane, H.; Nojiri, H. The Behavior and Significance of Degradative Plasmids Belonging to Inc Groups in Pseudomonas within Natural Environments and Microcosms. Microbes Environ. 2010, 25, 253–265. [Google Scholar] [CrossRef]
- Popowska, M.; Krawczyk-Balska, A. Broad-host-range IncP-1 plasmids and their resistance potential. Front. Microbiol. 2013, 4, 1–8. [Google Scholar] [CrossRef]
- Thomas, C.M. Paradigms of plasmid organization. Mol. Microbiol. 2000, 37, 485–491. [Google Scholar] [CrossRef]
- Jechalke, S.; Dealtry, S.; Smalla, K.; Heuer, H. Quantification of IncP-1 Plasmid Prevalence in Environmental Samples. Appl. Environ. Microbiol. 2013, 79, 1410–1413. [Google Scholar] [CrossRef]
- Taggar, G.; Rehman, M.A.; Boerlin, P.; Diarra, M. Molecular Epidemiology of Carbapenemases in Enterobacteriales from Humans, Animals, Food and the Environment. Antibiotics 2020, 9, 693. [Google Scholar] [CrossRef]
- Halat, D.H.; Moubareck, C.A. The Intriguing Carbapenemases of Pseudomonas aeruginosa: Current Status, Genetic Profile, and Global Epidemiology. Yale J. Biol. Med. 2022, 95, 507–515. [Google Scholar]
- Werner, K.A.; Poehlein, A.; Schneider, D.; El-Said, K.; Wöhrmann, M.; Linkert, I.; Hübner, T.; Brüggemann, N.; Prost, K.; Daniel, R.; et al. Thermophilic Composting of Human Feces: Development of Bacterial Community Composition and Antimicrobial Resistance Gene Pool. Front. Microbiol. 2022, 13, 824834. [Google Scholar] [CrossRef]
- Werner, K.A.; Schneider, D.; Poehlein, A.; Diederich, N.; Feyen, L.; Axtmann, K.; Hübner, T.; Brüggemann, N.; Prost, K.; Daniel, R.; et al. Metagenomic Insights Into the Changes of Antibiotic Resistance and Pathogenicity Factor Pools Upon Thermophilic Composting of Human Excreta. Front. Microbiol. 2022, 13, 826071. [Google Scholar] [CrossRef] [PubMed]
- de Bertoldi, M.; Sequi, P.; Lemmes, B.; Papi, T. (Eds.) The Science of Composting; Springer: Dordrecht, The Netherlands, 1996. [Google Scholar]
- Hartmann, A.; Locatelli, A.; Amoureux, L.; Depret, G.; Jolivet, C.; Gueneau, E.; Neuwirth, C. Occurrence of CTX-M producing Escherichia coli in soils, cattle, and farm environment in France (Burgundy region). Front. Microbiol. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pollett, S.; Miller, S.; Hindler, J.; Uslan, D.; Carvalho, M.; Humphries, R.M. Phenotypic and molecular characteristics of carbapenem-resistant Enterobacteriaceae in a health care system in Los Angeles, California, from 2011 to 2013. J. Clin. Microbiol. 2014, 52, 4003–4009. [Google Scholar] [CrossRef]
- Lachmayr, K.L.; Cavanaugh, C.M.; Kerkhof, L.J.; DiRienzo, A.G.; Ford, T.E. Quantifying nonspecific tem β-lactamase (blatem) genes in a wastewater stream. Appl. Environ. Microbiol. 2009, 75, 203–211. [Google Scholar] [CrossRef]
- Barraud, O.; Baclet, M.C.; Denis, F.; Ploy, M.C. Quantitative multiplex real-time PCR for detecting class 1, 2 and 3 integrons. J. Antimicrob. Chemother. 2010, 65, 1642–1645. [Google Scholar] [CrossRef] [PubMed]
- Klymus, K.E.; Merkes, C.M.; Allison, M.J.; Goldberg, C.S.; Helbing, C.C.; Hunter, M.E.; Jackson, C.A.; Lance, R.F.; Mangan, A.M.; Monroe, E.M.; et al. Reporting the limits of detection and quantification for environmental DNA assays. Environ. DNA 2020, 2, 271–282. [Google Scholar] [CrossRef]
- Tooke, C.L.; Hinchliffe, P.; Bragginton, E.C.; Colenso, C.K.; Hirvonen, V.H.A.; Takebayashi, Y.; Spencer, J. β-Lactamases and β-Lactamase Inhibitors in the 21st Century. J. Mol. Biol. 2019, 431, 3472–3500. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Mao, D.; Zhou, H.; Luo, Y. Prevalence and Fate of Carbapenemase Genes in a Wastewater Treatment Plant in Northern China. PLoS ONE 2016, 11, e0156383. [Google Scholar] [CrossRef]
- Liu, B.; Yu, K.; Ahmed, I.; Gin, K.; Xi, B.; Wei, Z.; He, Y.; Zhang, B. Key factors driving the fate of antibiotic resistance genes and controlling strategies during aerobic composting of animal manure: A review. Sci. Total Environ. 2021, 791, 148372. [Google Scholar] [CrossRef]
- Kaszab, E.; Szoboszlay, S.; Dobolyi, C.; Háhn, J.; Pék, N.; Kriszt, B. Antibiotic resistance profiles and virulence markers of Pseudomonas aeruginosa strains isolated from composts. Bioresour. Technol. 2011, 102, 1543–1548. [Google Scholar] [CrossRef]
- Pellegrini, C.; Mercuri, P.S.; Celenza, G.; Galleni, M.; Segatore, B.; Sacchetti, E.; Volpe, R.; Amicosante, G.; Perilli, M. Identification of blaIMP-22 in Pseudomonas spp. in urban wastewater and nosocomial environments: Biochemical characterization of a new IMP metallo-enzyme variant and its genetic location. J. Antimicrob. Chemother. 2009, 63, 901–908. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Toval, F.; Guzmán-Marte, A.; Madriz, V.; Somogyi, T.; Rodríguez, C.; García, F. Predominance of carbapenem-resistant Pseudomonas aeruginosa isolates carrying bla IMP and bla VIM metallo-β-lactamases in a major hospital in Costa Rica. J. Med. Microbiol. 2015, 64, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Cao, R.; Mo, S.; Yao, R.; Ren, Z.; Wu, J. Swine Manure Composting with Compound Microbial Inoculants: Removal of Antibiotic Resistance Genes and Their Associations with Microbial Community. Front. Microbiol. 2020, 11, 592592. [Google Scholar] [CrossRef]
- Zhou, Z.; Yao, H. Effects of Composting Different Types of Organic Fertilizer on the Microbial Community Structure and Antibiotic Resistance Genes. Microorganisms 2020, 8, 268. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Gu, J.; Wang, X.; Nasir, M.; Yu, J.; Lei, L.; Wang, Q. Elucidating the effect of microbial inoculum and ferric chloride as additives on the removal of antibiotic resistance genes from chicken manure during aerobic composting. Bioresour. Technol. 2020, 309, 122802. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Yang, X.; Li, Q.; Wu, L.; Shen, Q. Changes in antibiotic concentrations and antibiotic resistome during commercial composting of animal manures. Environ. Pollut. 2016, 219, 182–190. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, M.; Sui, Q.; Tong, J.; Jiang, C.; Lu, X.; Zhang, Y.; Wei, Y. Impacts of addition of natural zeolite or a nitrification inhibitor on antibiotic resistance genes during sludge composting. Water Res. 2016, 91, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Osińska, A.; Korzeniewska, E.; Harnisz, M.; Felis, E.; Bajkacz, S.; Jachimowicz, P.; Niestępski, S.; Konopka, I. Small-scale wastewater treatment plants as a source of the dissemination of antibiotic resistance genes in the aquatic environment. J. Hazard. Mater. 2020, 381, 121221. [Google Scholar] [CrossRef]
- Cacace, D.; Fatta-Kassinos, D.; Manaia, C.M.; Cytryn, E.; Kreuzinger, N.; Rizzo, L.; Karaolia, P.; Schwartz, T.; Alexander, J.; Merlin, C.; et al. Antibiotic resistance genes in treated wastewater and in the receiving water bodies: A pan-European survey of urban settings. Water Res. 2019, 162, 320–330. [Google Scholar] [CrossRef]
- Do, T.T.; Tamames, J.; Stedtfeld, R.D.; Guo, X.; Murphy, S.; Tiedje, J.M.; Walsh, F. Antibiotic Resistance Gene Detection in the Microbiome Context. Microb. Drug Resist. 2018, 24, 542–546. [Google Scholar] [CrossRef]
- Quirós, P.; Colomer-Lluch, M.; Martínez-Castillo, A.; Miró, E.; Argente, M.; Jofre, J.; Navarro, F.; Muniesa, M. Antibiotic Resistance Genes in the Bacteriophage DNA Fraction of Human Fecal Samples. Antimicrob. Agents Chemother. 2014, 58, 606–609. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhang, A.; Guo, T.; Zhu, Y.; Shao, Y. Biochar and hyperthermophiles as additives accelerate the removal of antibiotic resistance genes and mobile genetic elements during composting. Materials 2021, 14, 5428. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Lu, X.; Rensing, C.; Friman, V.P.; Geisen, S.; Chen, Z.; Yu, Z.; Wei, Z.; Zhou, S.; Zhu, Y. Hyperthermophilic composting accelerates the removal of antibiotic resistance genes and mobile genetic elements in sewage sludge. Environ. Sci. Technol. 2018, 52, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Zhou, G.; Wang, H.; Wu, X. The behavior of antibiotic-resistance genes and their relationships with the bacterial community and heavy metals during sewage sludge composting. Ecotoxicol. Environ. Saf. 2021, 216, 112190. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Xie, S.; Zeng, M.; Xu, X.; Li, Y.; Liu, X.; Wang, X. Impacts of pile temperature on antibiotic resistance, metal resistance and microbial community during swine manure composting. Sci. Total Environ. 2020, 744, 140920. [Google Scholar] [CrossRef]
- Dealtry, S.; Ding, G.-C.; Weichelt, V.; Dunon, V.; Schlüter, A.; Martini, M.C.; De Papa, M.F.; Lagares , A.; Amos, G.C.A.; Wellington, E.M.H.; et al. Cultivation-Independent Screening Revealed Hot Spots of IncP-1, IncP-7 and IncP-9 Plasmid Occurrence in Different Environmental Habitats. PLoS ONE 2014, 9, e89922. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Target Gene | Primer/ Probe | Sequence (5′→3′) | Amplicon Size [bp] | Positive Controls | Reference |
|---|---|---|---|---|---|
| blaCTX-M | q_CTXM-F | CAGCTGGGAGACGAAACGTT | 63 | E. coli 1058_16 * | [64] |
| q_CTXM-R | CCGGAATGGCGGTGTTTA | ||||
| q_CTXM-P | 6FAM- CGTCTCGACCGTACCGAGCCGAC-TAMRA | ||||
| blaIMP | q_blaIMP-F | ATTTTCATAGTGACAGCACGGGC | 105 | Klebsiella pneumoniae R77 IMP-4 | [65] |
| q_blaIMP-R | CCTTACCGTCTTTTTTAAGCAGCTCATTAG | ||||
| q_blaIMP-P | 6FAM- TTCTCAACTCATCCCCACGTATGC-TAMRA | ||||
| blaTEM | q_TEM-F | CACTATTCTCAGAATGACTTGGT | 85 | E. coli 832_16_2 * | [66] |
| q_TEM-R | TGCATAATTCTCTTACTGTCATG | ||||
| q_TEM-P | 6FAM- CCAGTCACAGAAAAGCATCTTACGG-TAMRA | ||||
| intI1 | q_intI-F | GCCTTGATGTTACCCGAGAG | 196 | E. coli pKJK5 #337 | [67] |
| q_intI-R | GATCGGTCGAATGCGTGT | ||||
| q_intI-P | 6FAM-ATTCCTGGCCGTGGTTCTGGGTTTT-TAMRA | ||||
| korB | q_korB-F | TCATCGACAACGACTACAACG | 118 | E. coli RP4 (IncP-1α) E. coli JM109 pB10 (IncP-1ß) E. coli JM109 pQKH545 (IncP-1γ) E. coli JM109 pKJK5 (IncP-1ε) | [58] |
| q_korB-Fz | TCGTGGATAACGACTACAACG | ||||
| q_korB-R | TTCTTCTTGCCCTTCGCCAG | ||||
| q_korB-Rge | TTYTTCYTGCCCTTGGCCAG | ||||
| q_korB-Rd | TTCTTGACTCCCTTCGCCAG | ||||
| q_korB-P | 6FAM- TCAGYTCRTTGCGYTGCAGGTTCTCVAT-TAMRA | ||||
| q_korB-Pgz | 6FAM- TSAGGTCGTTGCGTTGCAGGTTYTCAAT-TAMRA |
| ARG | Normalization | Decrease from Start to End of Composting [%] * | ||||
|---|---|---|---|---|---|---|
| E1 | E1-B | E2 | E2-B | Mean | ||
| blaIMP | 16S rRNA | +36.1 | +52.6 | +1096.2 | +308.2 | +373.3 |
| g compost | −29.5 | −12.2 | +689.3 | −9.5 | +159.5 | |
| blaCTX-M | 16S rRNA | −88.6 | +1323.3 | +499.9 | +285.8 | +505.1 |
| g compost | −94.1 | +773.9 | +245.8 | +14.1 | +234.9 | |
| blaTEM | 16S rRNA | −18.1 | −99.8 | +66.9 | +252.5 | +50.4 |
| g compost | −56.0 | −99.8 | −42.1 | +27.4 | −42.6 | |
| intI1 | 16S rRNA | −91.6 | −92.4 | −93.2 | −82.9 | −90.0 |
| g compost | −95.7 | −95.5 | −96.1 | −96.0 | −95.8 | |
| korB | 16S rRNA | −99.6 | −99.0 | −98.9 | −98.6 | −99.0 |
| g compost | −99.7 | −99.3 | −99.6 | −99.5 | −99.5 | |
| ARG | Mean Copy Numbers per Gram Compost | |
|---|---|---|
| Start | End | |
| blaIMP | 5.5 × 104 | 1.0 × 105 |
| blaCTX-M | 3.8 × 104 | 2.8 × 104 |
| blaTEM | 4.1 × 107 | 3.4 × 105 |
| intI1 | 5.0 × 108 | 2.1 × 107 |
| korB | 1.9 × 107 | 7.6 × 104 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Werner, K.A.; Feyen, L.; Hübner, T.; Brüggemann, N.; Prost, K.; Grohmann, E. Fate of Horizontal-Gene-Transfer Markers and Beta-Lactamase Genes during Thermophilic Composting of Human Excreta. Microorganisms 2023, 11, 308. https://doi.org/10.3390/microorganisms11020308
Werner KA, Feyen L, Hübner T, Brüggemann N, Prost K, Grohmann E. Fate of Horizontal-Gene-Transfer Markers and Beta-Lactamase Genes during Thermophilic Composting of Human Excreta. Microorganisms. 2023; 11(2):308. https://doi.org/10.3390/microorganisms11020308
Chicago/Turabian StyleWerner, Katharina A., Lara Feyen, Tobias Hübner, Nicolas Brüggemann, Katharina Prost, and Elisabeth Grohmann. 2023. "Fate of Horizontal-Gene-Transfer Markers and Beta-Lactamase Genes during Thermophilic Composting of Human Excreta" Microorganisms 11, no. 2: 308. https://doi.org/10.3390/microorganisms11020308
APA StyleWerner, K. A., Feyen, L., Hübner, T., Brüggemann, N., Prost, K., & Grohmann, E. (2023). Fate of Horizontal-Gene-Transfer Markers and Beta-Lactamase Genes during Thermophilic Composting of Human Excreta. Microorganisms, 11(2), 308. https://doi.org/10.3390/microorganisms11020308