Abstract
The exposure of microorganisms to conventional plastics is a relatively recent occurrence, affording limited time for evolutionary adaptation. As part of the EU-funded project BioICEP, this study delves into the plastic degradation potential of microorganisms isolated from sites with prolonged plastic pollution, such as plastic-polluted forests, biopolymer-contaminated soil, oil-contaminated soil, municipal landfill, but also a distinctive soil sample with plastic pieces buried three decades ago. Additionally, samples from Arthropoda species were investigated. In total, 150 strains were isolated and screened for the ability to use plastic-related substrates (Impranil dispersions, polyethylene terephthalate, terephthalic acid, and bis(2-hydroxyethyl) terephthalate). Twenty isolates selected based on their ability to grow on various substrates were identified as Streptomyces, Bacillus, Enterococcus, and Pseudomonas spp. Morphological features were recorded, and the 16S rRNA sequence was employed to construct a phylogenetic tree. Subsequent assessments unveiled that 5 out of the 20 strains displayed the capability to produce polyhydroxyalkanoates, utilizing pre-treated post-consumer PET samples. With Priestia sp. DG69 and Neobacillus sp. DG40 emerging as the most successful producers (4.14% and 3.34% of PHA, respectively), these strains are poised for further utilization in upcycling purposes, laying the foundation for the development of sustainable strategies for plastic waste management.
1. Introduction
Over the past few decades, there has been significant interest in identifying microorganisms capable of degrading plastic and the enzymes involved in plastic degradation. To date, only a limited number of microbial agents and their enzymes have been recognized and studied for plastic degradation, yet with minimal effectiveness. This could be attributed to the prevalence of uncultivated microbial species, which consequently remain unexplored in the context of the respective plastic-degrading environment [1]. Despite the high-molecular-weight polymers (e.g., polypropylene (PP), polystyrene (PS)), potential degraders are widely spread across the microbial tree of life, and evidence for plastic degradation by a majority of these taxa is still limited [2]. However, when it comes to polyethylene terephthalate (PET), various bacteria (e.g., Thermobifida fusca, Thermomonospora curvata, and Ideonella sakaiensis) [3,4,5] and fungi (e.g., Fusarium solani, Humicola insolens, and Aspergillus oryzae) [6,7,8] have been proven to possess enzymatic machinery to degrade this polymer.
Microorganisms have been exposed to conventional plastics only recently, allowing limited time for their evolution. However, microbes capable of breaking down plastic can be found in various environments, such as soil, seawater, and plastic-polluted sites [9]. Landfill sites exhibit significant heterogeneity owing to the complexity of substrates and the abundance of organic and inorganic matter, making them a recognized reservoir of microbial diversity [10]. Hence, many research groups are reporting results on plastic degraders discovered in environments involving severe or moderate plastic pollution [11,12,13]. Ever since Bowditch et al. [14]. reported the ability of Lepidoptera and Coleoptera to partially degrade polyvinyl chloride and PP packaging films, the scientific community has turned their attention to this unconventional source of plastic degraders. Kim et al. [15] found bacteria in the intestines of adult bees with the ability to degrade PET, while Pham et al. [16] reported the ability of mealworm microbiota to degrade various consumer plastics. The existing information regarding the involvement of insects and their microbiota in plastic decomposition is still quite restricted, leaving numerous questions unanswered about the intricate process of plastic breakdown facilitated by insects. The specific mechanisms and the role of enzymes in this degradation process are yet to be conclusively determined. However, insects certainly do exhibit the capability to degrade compounds that are typically challenging and potentially can find practical applications in waste management programs.
In recent years, notable attention and interest in bioplastics have risen, reflecting a worldwide trend toward the preference for biodegradable products by consumers. Bioplastics stand out due to their inherent biodegradability, sustainability, and environmentally friendly properties, presenting numerous advantages over conventional petrochemical-based plastics [17]. However, it is important to point out that not all biodegradable materials are biobased. Biobased materials are derived from renewable resources, from living (or once-living) organisms, while biodegradable materials possess the ability to decompose in a given environment over a reasonable amount of time [18]. The most extensively studied materials, both biobased and biodegradable, are polyhydroxyalkanoates (PHAs). These materials are poised to replace certain contemporary petrochemically derived plastics due to their biodegradable nature and favourable thermoplastic and mechanical properties. These include versatility, elasticity, flexibility, and other desirable characteristics [19]. PHA production using waste streams from several industries, municipal waste, and households is already well examined and established [20,21].
Nevertheless, recent studies have indicated that plastic waste can be used for the same goal [22,23]. The biological process of plastic upcycling involves utilizing plastic waste streams as a carbon substrate for biotechnological processes, following a methodology employed for lignocellulosic feedstocks [24]. Numerous enzymes with hydrolytic properties have been identified, capable of depolymerizing specific plastics with the possibility or specifically engineered to transform depolymerized plastics into high-value products, such as biopolymers [22,25,26]. However, the effectiveness of the biological route is often impeded by the inherent recalcitrance of plastics, typically associated with hydrophobicity and crystallinity.
Presently, recycling technologies predominantly yield lower-value products from plastic waste (downcycling) or, at best, products of similar value. To propel the development of technologies facilitating plastic upcycling, the European Commission has allocated significant funding to research and innovation projects, encompassing both chemical and biological processes. Bio innovation of a circular economy for plastics (BioICEP) is one of EU-funded projects tackling plastic waste management and upcycling. The overarching aim of the project is to illustrate a streamlined and sustainable approach toward achieving a circular economy for plastics. This involves the creation of an advanced, energy-efficient, and cost-effective method for biotransforming waste plastic into bioproducts and bioplastics that are in high demand in the market. As a part of the BioICEP project, the first steps of this study involved the creation of microbial collection from soil samples tainted with plastic waste and samples gathered from two types of insects. Based on the literature reported earlier in this section, this approach was selected to increase the probability of isolating microbes equipped with the requisite enzymatic machinery for plastic degradation. Overall, this study delved into the microbial diversity of plastic-polluted sites, exploring their proficiency in degrading various plastic-related substrates as sole carbon and energy sources. The ability of these strains to produce high-value molecules such as biosurfactants and PHA was analysed. Furthermore, the confirmed capacity of selected strains to utilize plastic-related substrates and generate biopolymers added depth to our understanding of their ecological roles in plastic degradation and possible use for plastic upcycling.
2. Materials and Methods
2.1. Collection of Samples
Various sampling sites were chosen to be examined as a source of microorganisms with potential ability to degrade plastic-related substrates, such as plastic-polluted sites, landfills, oil-polluted sites, but also, arthropods (isopods and arachnids) were collected as a sampling material. The number of sites, their origin, and nature are presented in Table 1.

Table 1.
Summary of sites used for sample collection for this study.
2.2. Bacterial Isolation
Microbial isolation was carried out via the serial dilution method using different selective media in order to facilitate various microbial species growth. Thus, 1 g/L of samples was used as a starting point. For arachnids and isopods, samples were washed with 70% (v/v) ethanol, rinsed in distilled water, and followed up with an overnight freeze at −80 °C. Samples were thoroughly macerated, and 1 g/L was used to prepare serial dilutions. For these purposes, Lysogeny broth (LA: 10 g/L tryptone, 5 g/L yeast extract, 10 g/L NaCl, 15 g/L agar), Sabouraud Dextrose (SAB: 40 g/L glucose, 10 g/L peptone, 15 g/L agar), and Tryptic Soy Agar (TSA: 17 g/L casein peptone (pancreatic), 2.5 g/L, dipotassium hydrogen phosphate, 2.5 g/L glucose, 5 g/L sodium chloride, 3 g/L soya peptone, 15 g/L agar) were used. After 10 days of incubation at 30 °C and 37 °C, morphologically distinct strains were selected and separated. Obtained pure cultures were stored in glycerol (20%, v/v), maintained at −80 °C, and used for the inoculation of cultures for further experiments. Morphological characteristics of selected pure cultures were recorded on TSA plates using Stereo Microscope Olympus szx10, 10× magnification (Boston, MA, USA) [27,28].
2.3. Agar-Based Screening Methodology
The ability to utilize different plastic monomers and oligomers as a sole carbon and energy source was investigated using Mineral Salt Medium agar plates (MSM): 15 g/L agar, 9 g/L Na2HPO4 × 12H2O, 1.5 g/L KH2PO4, 1 g/L NH4Cl, 0.2 g/L MgSO4 × 7H2O, 0.2 g/L CaCl2 × 2H2O, 0.1% trace elements solution, 0.025% N-Z amine and carbon source 5 g/L. Plastic-related substrates were used as a sole carbon source: terephthalic acid powder (TPA; Sigma Aldrich, London, UK), bis(2-hydroxyethyl) terephthalate powder (BHET; Sigma Aldrich, UK), polyethylene terephthalate powder (PET; Sigma Aldrich, UK), anionic aliphatic polyester-polyurethane dispersion (Impranil DLN-SD, Covestro Ltd., London, UK), and anionic polycarbonate polyurethane dispersion (Impranil DL2077; Covestro Ltd., London, UK), whereas glucose was used as a positive control. The applied method is based on the combination of several references and previous experience [29,30,31].
2.4. 16S rRNA Sequencing
Strains selected as the best performers in screening assays were identified via 16S rRNA sequence analysis. These strains were cultivated in tryptone soy broth (TSB) (17 g/L casein peptone (pancreatic), 2.5 g/L, dipotassium hydrogen phosphate, 2.5 g/L glucose, and 5 g/L sodium chloride, 3 g/L soya peptone) at 30 °C and 37 °C for 4 days shaking at 180 rpm for DNA isolation [32]. Strains were identified by 16S rRNA sequence analysis using universal bacterial primer set: 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′). PCR amplification was performed in a 2720 Thermal Cycler (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA) using Phusion High-Fidelity PCR Master Mix (F-531S) (Thermo Scientific, Waltham, MA, USA) following the manufacturer’s protocol. PCR products were purified using a PCR purification kit (Qiagen, Hilden, Germany). Sequencing was performed using the Sanger sequencing method at Source Bioscience (Cambridge, UK). 16S rRNA sequences were identified using BLASTN program [33].
2.5. Phylogenetic Analysis
The taxonomic verification of the isolated bacterial strains was carried out through an examination of the 16S rRNA sequence data. These sequences were compared with those deposited in the GenBank database of NCBI using the BLAST similarity search tool “https://blast.ncbi.nlm.nih.gov/Blast.cgi” (accessed on 10 September 2023). The sequence with the highest total score, substantial percentage query coverage, high identity percentage, and lowest E-value was chosen from all the identified homologous nucleotide sequences. Sequences exhibiting a sequence similarity of >98% to their closest phylogenetic neighbor over an average range of 1300–1400 bp were classified at the species level. Sequences with <98.0% sequence similarity were categorized at the genus level, and the percentage sequence similarity is indicated in brackets. The 16S rRNA sequences were submitted to GenBank and assigned the following accession numbers: OR693299, OR693300, OR693301, OR693302, OR693303, OR693304, OR693305, OR693306, OR693307, OR693308, OR693309, OR693310, OR693311, OR693312, OR693313, OR693314, OR693315, OR693316, OR693317, and OR693318. The evolutionary relationship was inferred using the neighbor-joining method [34]. The optimal tree is shown. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) is shown next to the branches [35]. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Tamura–Nei method and are in the units of the number of base substitutions per site [36]. This analysis involved 24 nucleotide sequences. Codon positions included were 1st + 2nd + 3rd + Noncoding. All ambiguous positions were removed for each sequence pair (pairwise deletion option). There were a total of 1405 positions in the final dataset. Evolutionary analyses were conducted in MEGA11 [37].
2.6. Upcycling Capabilities of Selected Isolates
2.6.1. Emulsification Assay
For emulsification activity determination, 2 mL of cell-free supernatant and 2 mL of sunflower oil were mixed in test tubes. The mixtures were vigorously vortexed for 1 min and allowed to stand for 24 h at room temperature. By adopting the formula given below, the emulsification index (EI, %) was calculated (Equation (1)). Polysorbate 80 was used as a positive control (EI 100%).
2.6.2. Reactive Extrusion
PET depolymerization experiment via reactive extrusion (REX) was conducted on a benchtop PrismTM twin-screw extruder (Thermo Electron GmbH, Karlsruhe, Germany) using a modified procedure to that of Biermann et al. (2021) [38]. Post-consumer PET material (PET tray, provided by Novelplast, Gibstown, Ireland) was mixed with solid NaOH in a ratio (2:1) in a sealed plastic bag, and the mixture was dispensed through the main shaft into the barrel. The temperature of the barrel was kept constant at 250 °C, and the rotational speed of the screws was adjusted to 20 rpm. Unreacted porous polymer residue was used for PHA production experiments as a sole carbon source.
2.6.3. Bacterial Cultivation for PHA Production
The cultures were reactivated from cryopreserved stock cultures (in 20% (v/v) glycerol, at −80 °C) by plating into solid Luria Bertani (LB) medium (tryptone, 10 g/L; yeast extract, 5 g/L; NaCl, 10 g/L; agar, 15 g/L; pH 7.0). The plates were incubated at 30 °C until isolated colonies formed. The inoculum for the shake flask experiments was prepared by inoculating a single colony in 50 mL liquid LB medium (tryptone, 10 g/L; yeast extract, 5 g/L; NaCl, 10 g/L; pH 7.0) and incubation in an orbital shaker (200 rpm), at 30 °C, for 24 h. For the shake flask assays, modified Medium E* (pH 7.0) was used, with the following composition: (NH4)2HPO4, 1.1 g/L; K2HPO4, 5.8 g/L; KH2PO4, 3.7 g/L; 10 mL/L of a 100 mM MgSO4 solution and 1 mL/L of a micronutrients’ solution. The micronutrient solution was composed of (per litre of 1 N HCl): FeSO4⋅7H2O, 2.78 g; MnCl2⋅4H2O, 1.98 g; CoSO4⋅7H2O, 2.81 g; CaCl2⋅2H2O, 1.67 g; CuCl2⋅2H2O, 0.17 g; and ZnSO4⋅7H2O, 0.29 g. The medium was supplemented with REX residuals (REX PET) at a concentration of 20 g/L. The assays were performed in 500 mL baffled shake flasks containing 200 mL of Medium E* supplemented with REX PET, which were inoculated with 20 mL of the inoculum prepared as described above. Incubation was performed in an orbital shaker (200 rpm) at 30 °C.
2.6.4. Quantitative and Qualitative Analysis of Derived PHAs
Cell growth was monitored during the cultivation by measuring the culture broth’s optical density (OD) at 660 nm. The presence of intracellular granules of PHA was assessed by visualization using fluorescence microscopy (BX51, Olympus, Boston, MA, USA) of the cells stained with Nile Blue A (Sigma Aldrich, UK), as described by Silvestre et al. [39]. At the end of the run, the cell pellet was collected by centrifuging the culture broth (10,956× g, 15 min, 4 °C) and lyophilized. The resulting dry biomass was used for the gravimetric determination of the cell dry weight (CDW) and for PHA analysis by gas chromatography (GC), as described by Rebocho et al. [40]. PHA content in the biomass and its composition were determined by GC after acidic methanolysis of dried cells. Samples were mixed with 2 mL of a sulphuric acid 20% (v/v) (Sigma- Aldrich, HPLC grade, UK) solution in methanol (Fisher Chemical, HPLC grade, UK) and 2 mL of benzoic acid (1 g/L) in chloroform (Sigma Aldrich, HPLC grade, UK) solution, at 100 °C, for 4 h. Benzoic acid acted as internal standard. The calibration curve was prepared with standards of P(HB-co-HV) (Sigma Aldrich) composed of 86 mol% 3-hydroxybutyrate (3HB) and 14 mol% 3-hydroxyvalerate (3 HV), at concentrations ranging from 0.1 to 1.0 g/L. The obtained methyl esters were analysed in a gas chromatograph equipped with a flame ionization detector (FID) (Sigma Aldrich, UK) and a Restek column (Crossbond, Stabilwax, Tokyo, Japan) at constant pressure (96 kPa) using helium as carrier gas. Splitless injection was used. The oven temperature ramp was the following: 20 °C/min until 100 °C; 3 °C/min until 155 °C; and, finally, 20 °C/min until 220 °C. The yields of purity obtained for the biopolymers were calculated from Equation (2):
The polymer was extracted from the dried biomass by Soxhlet extraction with chloroform (~5 g biomass for 250 mL chloroform), at 80 °C, for 48 h. Cell debris were removed by filtration with filters with a pore size of 0.45 μm (GxF, GHPmembrane, PALL, UK), and the polymer was precipitated in ice-cold ethanol (1:10, v/v) under vigorous stirring. The polymer was recovered and dried at room temperature.
2.6.5. Estimation of TPA Content in Cultivation Media by HPLC Analysis
Cultivation broth was centrifuged (13,000 rpm, 15 min) and supernatants were filtered (0.2 μm, VWR) and diluted with 30 mM NaOH in order to assess the amount of TPA. HPLC method was applied using anion exchange column (Ionpac AS11-HC 4.6 × 250 mm equipped with a pre-column) coupled to a conductivity detector. Analysis was performed at 30 °C, and 30 mM NaOH was used as an eluent at the 1.5 mL/min flow rate. TPA standards in concentration range from 0.006 to 1.0 g/L were used for calibration curve construction.
3. Results and Discussion
3.1. Isolation and Screening of Microbial Strains from Plastic-Polluted Sites
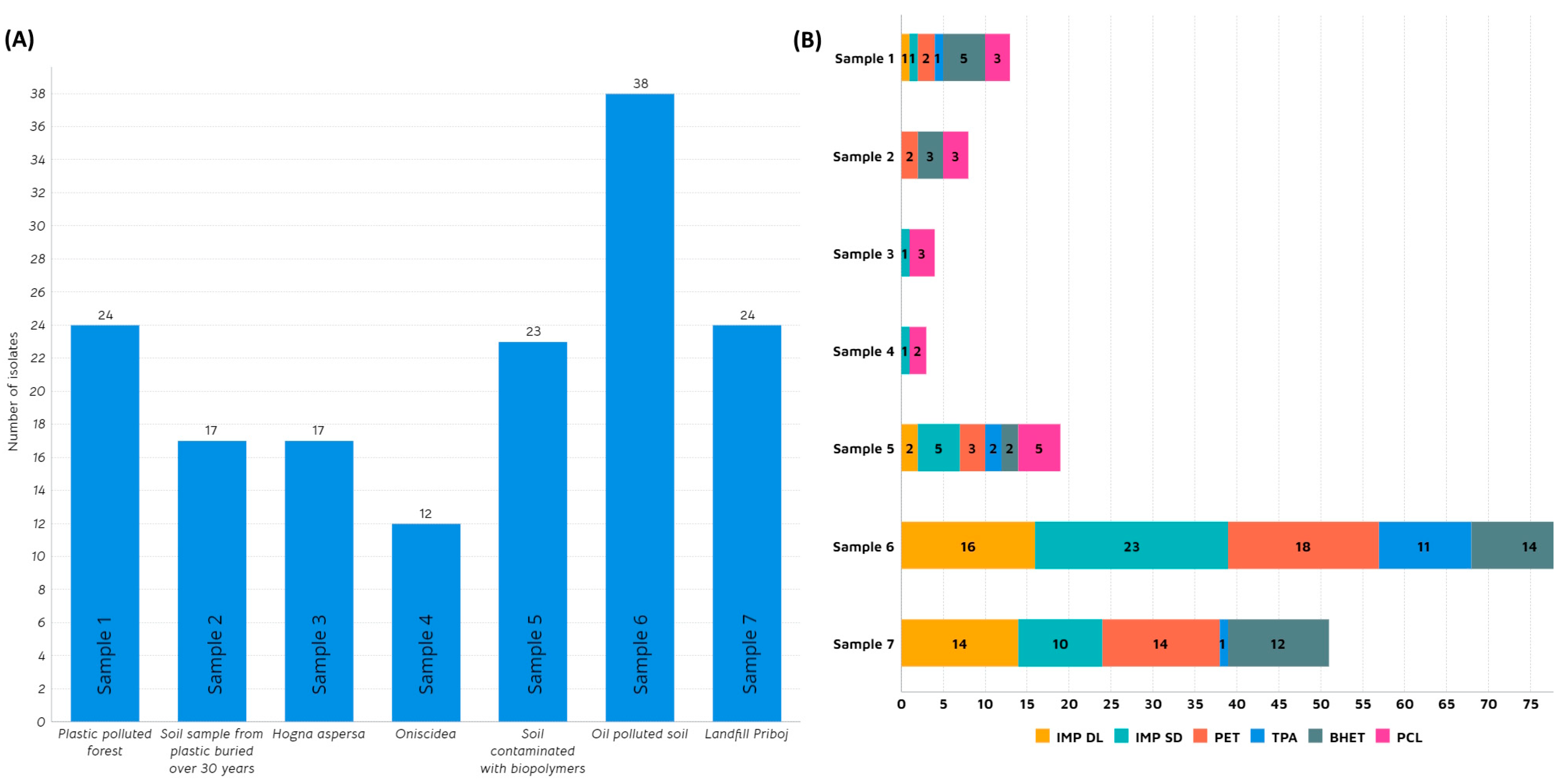
Microorganisms possess inherent or acquired capacities to endure in environments with extreme conditions, such as adaptive changes in transcriptomic, metabolic, and signalling pathways [41]. Adapting to plastic-contaminated surroundings includes biofilm formation [42], utilization of different pathways [43,44], and, predominantly, the secretion of specific depolymerizing enzymes that enable them to use persistent plastic pollution as an additional carbon source, giving them a selective advantage [45]. In this study, we tried to harness the potential of microbial communities to degrade plastic-related substrates by targeting sites where plastic pollution is present for a significant amount of time. A total of seven samples were collected and analysed (Table 1). Samples 1, 5, and 6 were obtained from a plastic-polluted forest, biopolymer-polluted soil, and municipal landfill, respectively. Sample 2 was provided by an Irish farmer who, predicting the plastic pollution problem, buried plastic pieces in soil 3 decades ago. Following up on the concept of insects as a biodegrading tool for synthetic polymers [46], samples 3 and 4 were collected from Arthropoda species: Oniscidea (woodlice) and Hogna aspersa (garden wolf spiders), respectively. On the other hand, sample 7 was retrieved from oil-polluted soil in Mexico. According to the research performed by Gambarini et al. [2], the majority of presumed plastic degraders were isolated from soil samples (27.8%), followed by plastic waste disposal sites (9.6%) and composts (5.3%). Additionally, a significant portion was sourced from culture collections of microorganisms (15.9%). As illustrated in Figure 1A, the highest number of morphologically diverse isolates, proficient in degrading plastic, was identified in sample 6 from landfill (25%).
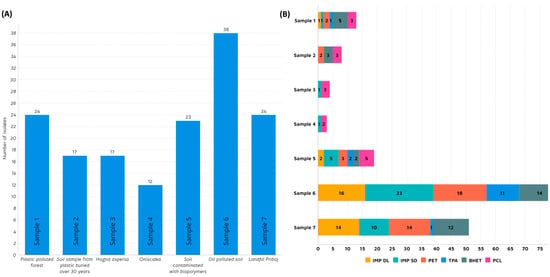
Figure 1.
(A) Number of isolated strains per sample; (B): Number of positive isolates per substrate in agar plate screening: Impranil® DL 2077 (IMP DL), Impranil® DLN-SD (IMP SD), Polyethylene terephthalate (PET), Terephthalic acid (TPA), Bis(2-hydroxyethyl) terephthalate (BHET) and Polycaprolactone (PCL).
This landfill, situated in the small city of Priboj in Western Serbia, poses a significant ecological challenge for the region due to the volume of waste and inadequate disposal and incineration methods. Having been covered with plastic waste for several decades, this area emerged as an ideal starting point in the quest for plastic-degrading microbes. Kopecka et al. (2022) examined isolates from plastic waste deposited in the municipal landfill over 2–17 years and discovered several bacterial strains with the ability to degrade high-density polyethylene (HDPE) [47]. In this study, out of 38 isolates, 23 showed the ability to grow on anionic aliphatic polyester-polyurethane dispersion (IMP DLN-SD) while 16 grew on anionic polycarbonate polyurethane dispersion (IMP DL) as a sole carbon source (Figure 1B). This potential discovery holds promise for addressing polyurethane (PU)-related plastic waste, which constitutes 9% of global plastic production, according to Geyer et al. [48]. A noteworthy proportion of isolates from this source demonstrated the capability to utilize PET and PET-related substrates (TPA and BHET). Another sample that showed interesting potential in this study came from a site constantly polluted by oil, located in Poza Rica, east-central Mexico (Sample 7). Further, 24 strains were isolated in total, out of which 58% showed the ability to grow on impranil and PET. It is interesting to note that none of these strains were capable of using PCL as a carbon source. However, in sample 5 (soil polluted with biopolymers), growth on PCL was detected for five strains. Samples related to insects (samples 3 and 4) did not yield exceptional results, except for five strains displaying the ability to grow on PCL. Sample 1 was derived from a forest mildly polluted with plastics and, thus, considered relatively pristine. Despite isolating 24 different strains, only a handful exhibited potential in utilizing plastic-related substrates, with five strains for BHET, three for PCL, and two for PET.
3.2. Most Promising Strains Selection and Characterization
Based on agar screening results, 20 isolates were selected for further investigation, taking into consideration the following factors: their ability to grow on at least three out of five examined substrates as a single carbon and energy source; prominent growth on at least one substrate; incubation period (Table 2). These isolates were further identified through 16S rRNA sequencing, and their morphological characteristics were recorded and summarized (Figure 2).
Table 2.
Selected strains’ performance on examined substrates.
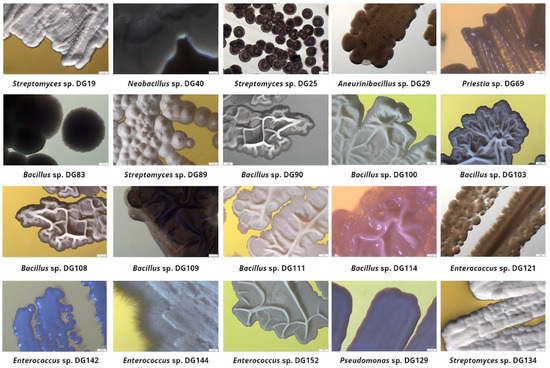
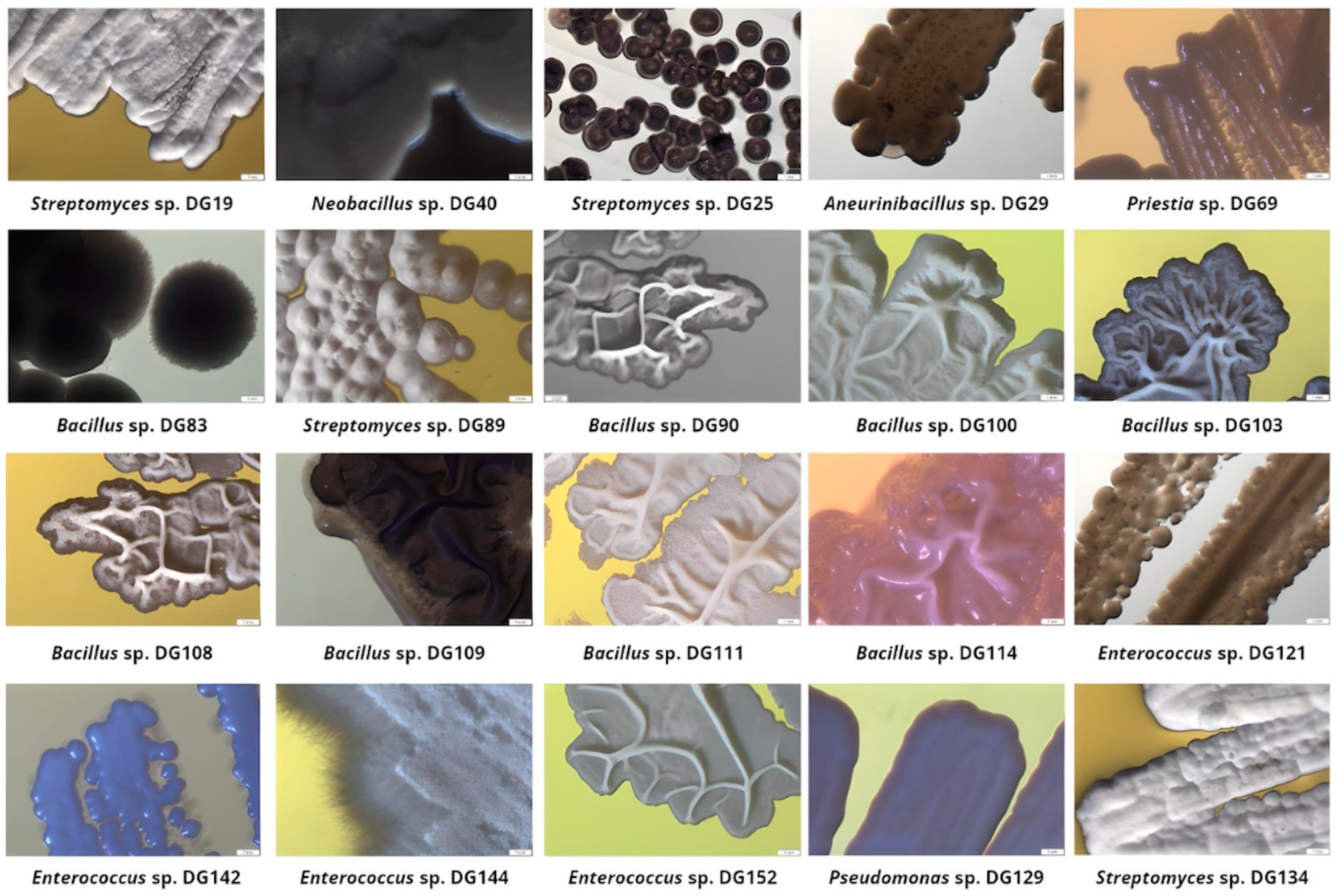
Figure 2.
Selected isolates’ colony morphology after incubation on Tryptone Soy Agar plates (magnification 10×).
Sample 1: Plastic-polluted forest
Strain 19 expressed moderate growth on all examined substrates apart from TPA. This strain 16S rRNA sequence (1375 bp length) showed 100% similarity with Streptomyces strains according to NCBI Blast Nucleotide base analysis. The Streptomyces genus exhibited significant potential in several industries; their enzymatic toolbox and the ability to degrade polysaccharides [49] position them as a compelling candidate for bioremediation and biological control across diverse ecosystems. Results reported by Rodríguez-Fonseca et al. demonstrated the significant potential of Streptomyces spp. in the degradation of synthetic polymers, especially polyolephins [50]. Streptomyces sp. DG19 exhibits typical behavior of this genus when incubated on the TPA plate, forming compact white colonies with smooth edges [51].
Strain 40 expressed the ability to grow on four out of five examined substrates with moderate growth on BHET. This strain was selected due to the ability to grow fast in examined conditions. According to 16S rRNA sequence analysis, 1204 bp aligned with the highest percentage (99.67%) with Neobacillus spp. These species, when present as a part of rhizosphere microbiota, were found to promote plant performance in saline–alkali soil under plastic film mulching. Janakiev et al. isolated and examined species related to this genus from bacteriobiota associated with the gut of Chironomus riparius larvae and discovered potential candidates for the formulation of eco-friendly approaches to break down organic pollutants and microplastics within freshwater ecosystems [52]. Neobacillus sp. DG 40 showed the ability to grow extremely fast on TSA plates, forming round, smooth, and cream colonies, orange in liquid culture.
Sample 2: Soil sample from plastic buried over 30 years ago
Isolate 69 was selected for further research due to its ability to grow on four out of five offered substrates (all but TPA), especially on Impranil. This strain developed silky-smooth colonies typical for Bacillus spp., and BLAST analysis of a 1178 bp sequence suggested the highest similarity with Priestia megaterium sp. (basonym: Bacillus megaterium) [53]. In the literature, this strain is described as important in biotechnology due to its role as an expression platform for recombinant proteins and vitamins, as well as contributing to bioremediation activities [54]. Meng Tan et al. efficiently used this strain for polystyrene degradation [55]. In addition, this strain is well known as a significant PHB producer under high-salt conditions [56].
Sample 5: Soil contaminated with biopolymers
Isolate 83 exhibited prominent growth on Impranil and PET. After incubation on the TSA plate, it formed round, fuzzy grey colonies. 16S rRNA sequence analysis (1398 bp) indicated 100% similarity to the Bacillus murallis. This strain was identified as an LDPE-degrading bacteria and PE waste-colonizing bacteria in the literature [57,58].
Isolate 89 exhibited strong growth on PET substrate. Fuzzy-white, round colonies with smooth edges on the TSA plate suggested this isolate belongs to the Streptomyces genus, which was further confirmed by 16S rRNA analysis (1378 bp, 100% similarity).
Another strain obtained from this sample, selected for further research, was isolate 90, with the ability to grow on all examined substrates. Opaque, cream-colored and slightly convex colonies, with irregular margins, were observed after incubation on TSA plates. Thus, the 1377 bp 16S sequence showed 100% similarity with Bacillus circulans according to BLAST analysis. In a study performed by Saikia et al., this strain was found as part of the community in the gut of the greater wax moth (Galleria mellonella linnaeus), able to degrade polycyclic aromatic hydrocarbons, low-density polyethylene, and 2-methylphenanthrene [59]. This strain is also known as a PHA producer [60,61].
Sample 6: Oil-polluted soil
All selected isolates from this sample were identified as Bacillus spp. Among the publications employing naturally occurring PET-degrading microorganisms, Bacillus spp. was found to be the most frequent bacterial genus [62,63]. All selected strains exhibited the ability to grow on all examined substrates apart from the 114 strain that did not utilize TPA. Isolates 100, 103, and 108 looked similar when grown in TSA and TSB media, but due to their distinctive behavior on MSM plates and in the presence of various substrates, it was presumed that these strains are morphologically similar but not identical. These isolates’ colonies were observed as white, flat, opaque, rough, and wrinkled, with irregular edges on the TSA medium. According to 16S rRNA analysis, these three strains are over 99% similar to each other. The sequence of isolates 100 and 103 (1418 and 1414 bp, respectively) showed 99.86% and 100% similarity to Bacillus velezensis, respectively. It is well established that this Bacillus strain has the capacity to generate diverse enzymes, including protease, cellulase, amylase, and glucanase. Additionally, it secretes antibacterial substances, effectively inhibiting the growth of pathogenic microorganisms [64,65]. Gui et al. discovered a deep-sea B. velezensis with the ability to break down waterborne polyurethane [66]. Also, it has been reported that in the community with Stenotrophononas maltophilia and Acinetobacter radioresistens, this Bacillus strain can affect the decomposition of polystyrene microplastics [66]. Isolate 108 (1333 bp) had the highest similarity to Bacillus subtilis (100%). B. subtilis is highly resistant to challenging environments and has been used for the secretion of proteins, capable of degrading various pollutants [67]. This quality positions it as a promising microbial tool for biodegradation. Other than PET’s degrading potential [68], Yao et al. reported the ability of B. subtilis ATCC6051 to degrade PE [69], and it has been listed as a potential PU degrader [70].
Isolate 109 formed cream-colored colonies, irregular in shape, with undulate margins on the TSA medium. It was established that this strain sequence (1369 bp) has 100% similarity to Bacillus licheniformis, a well-established bio-surfactant producer [71]. It was found that in the presence of organoclays, this strain can accelerate the degradation of PLA [72], but also when combined with other Bacillus spp., it utilizes PP and is able to degrade PP-PLA blends [73].
When grown on the TSA medium, isolate 111 formed milky-white colonies with undulate edges and a wrinkled surface. Further, 100% similarity to Bacillus subtilis, subsp. stercoris, and Bacillus safensis was detected by BLAST analysis of 16S rRNA sequence (1387 bp). B. safensis is known for its ability to degrade PLA [74] but, moreover, it was found by Waquas et al. that this strain can degrade 18.6% LDPE in 30 days [75].
Isolate 114 showed remarkable ability to grow on Impranil-related substrates, exhibiting a halo zone around the irregularly shaped, white colonies with smooth edges. The obtained sequence of this isolate with 1405 bp aligned with B. licheniformis and several partial sequences of Bacillus spp. isolates (100%).
Sample 7: Landfill in Priboj
All strains selected from sample 7 had the ability to efficiently grow on all examined substrates. Isolates 121 and 142 formed cream, round colonies with smooth edges on TSA medium. Despite being morphologically similar, these strains exhibited different behavior under the examined conditions: 121 expressed profound growth on TPA with a distinctive halo around colonies. Isolate 144, on the other hand, formed white, textured colonies with filiform margins, while 152 had wrinkled grey colonies with smooth margins when grown on TSA. All four of these isolates showed the highest similarity to the Enterococcus hirae (100%). Enterococcus spp. is rarely connected with plastic degradation research. The ability of E. faecailis to degrade polyamide 6 was examined, but the results were not as significant as with Alcaligene faecalis [76]. Their ability to colonize LDPE and biopolymers was also investigated with the aim to highlight potential concerns for human health due to the fast pathogens’ biofilm development, not in light of the biodegradation assessment [77]. They have been described as biopolymer producers in the literature [78]. For example, Bhuwal et al. described them as a good candidate for PHB industrial production from cardboard industry wastewater [79]. In this study, isolated Enterococcus spp. exhibited an amazing ability to utilize plastic-related substrates as a sole carbon and energy source, making these strains interesting candidates for further research.
Isolate 129 showed exquisite performance during the screen. It was able to grow on all examined substrates, forming a halo zone with the TPA substrate as a sole carbon source. Colonies were disc-shaped with ridges radiating from the center when grown on TSA medium. 16S rRNA sequence (1392 bp) had 100% similarity to Pseudomonas stutzeri. Pseudomonas spp. is well described in the literature regarding plastic degradation. Highly effective degradation of high-molecular-weight polyethylene glycols (PEGs) achieved through pure cultures of P. stutzeri was reported by Obradors et al. in 1991 [80]. This strain was found to degrade the PET monomers terephthalic acid and ethylene glycol [81], but also affect the deterioration of HDPE microplastics [82]. Uefuji et al. reported that this strain secretes PHB depolymerase [83].
Isolate 134 was selected due to its excellent performance in the initial screen. Morphologically, this strain behaved as previously described for Streptomyces spp.: white, round colonies with smooth edges and textured surface. This was further confirmed by BLAST analysis, where the 16S rRNA sequence (1374 bp) had 99.93% similarity to Streptomyces spp.
3.3. Phylogenetic Analysis
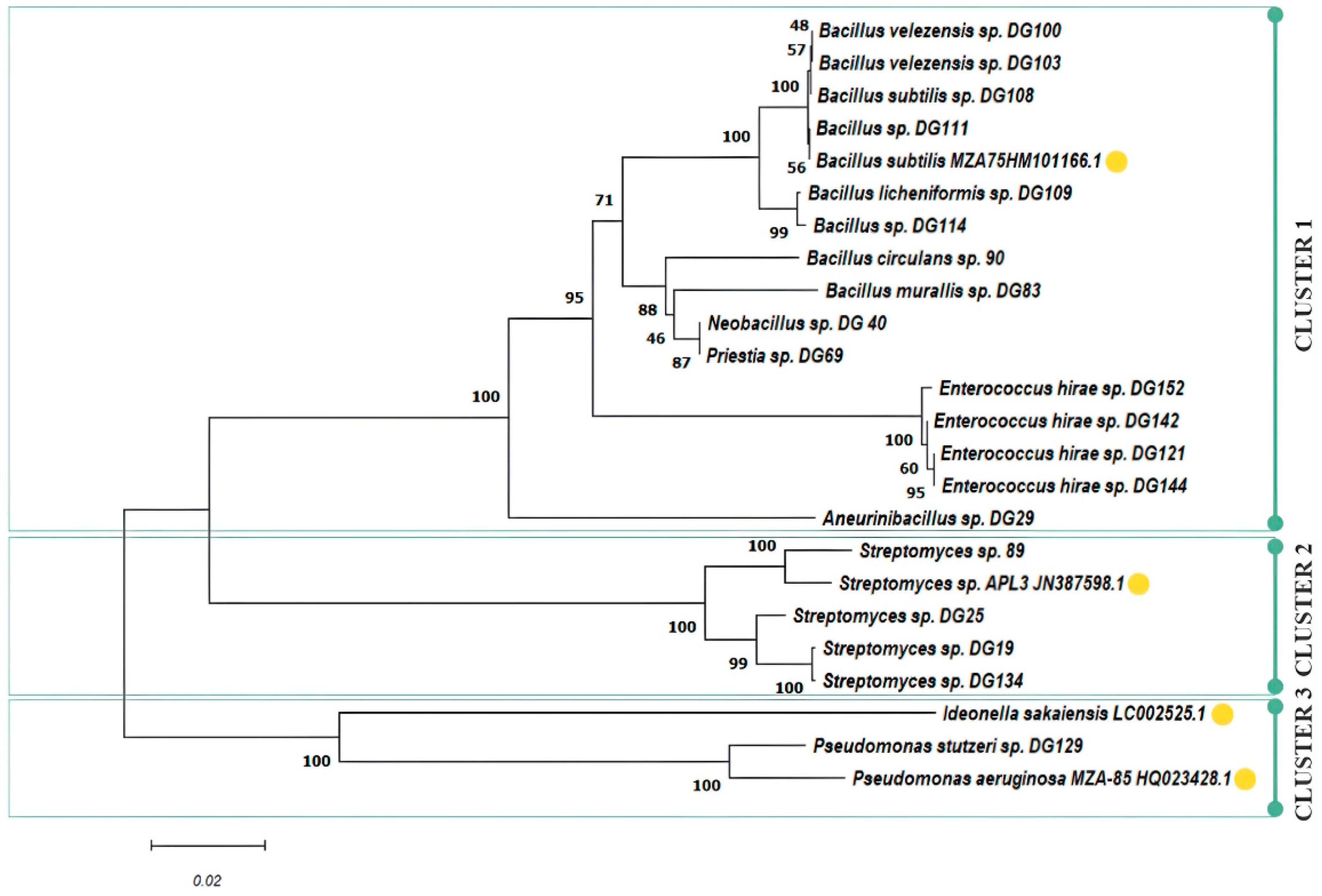
The number of plastic-degrading microorganisms reported is rapidly increasing, making it possible to explore the conservation and distribution of presumed plastic-degrading traits across the diverse microbial tree of life. Presumed degraders of conventional high-molecular-weight polymers, like polyamide, polystyrene, polyvinyl chloride, and polypropylene, are distributed extensively across bacterial and fungal branches of the tree of life [84]. As of April 2020, the cumulative count of species documented to possess plastic-degrading capabilities, using specified search terms by Gambarini et al., reached 436 [2], with the initial mention dating back to a publication in 1974 [85]. The same authors reported a total of 16,170 potential plastic degradation orthologs identified across 6000 distinct microbial strains. These strains span 12 different phyla, with 5 of them lacking reported species capable of plastic degradation. In this study, 10 Bacillus and Bacillus-related strains, 4 Enterococcus spp., 4 Streptomyces spp., and 1 Pseudomonas sp. were isolated from various sources and selected for further research. A phylogenetic tree was constructed, including several bacterial strains that are widely known as plastic degraders in the literature to assess the genetic relationship of isolated stains: B. subtilis MZA-75 (polyurethane degrader) [86], P. aeruginosa MZA-85 (polyurethane degrader) [87], Ideonella sakaiensis (PET degrader), and Streptomyces sp. APL3 (polyester-based plastics degrader) [88]. The phylogenetic tree constructed based on neighbour-joining analysis of 16S rRNA clustered 24 examined isolates in three clusters and eight subclusters (Figure 3).
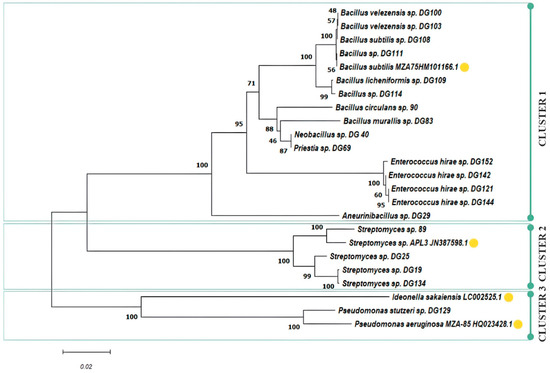
Figure 3.
Phylogenetic tree of selected isolates including literature examples of known plastic degraders (yellow circle).
Cluster 1 forms an assemblage of Bacillus- and Enterococcus-related species, with four distinctive subclusters. In subcluster 1, isolates 100, 103, 108, 109, 111, and 114 closely align with the previously characterized PU degrader, B. subtilis MZ-75. This result is in accordance with previously presented screening results, where these isolates exhibited similar behaviour, with an emphasis on PU-related substrate consumption under the examined conditions. Subcluster 2 encompasses isolates 90, 83, 40, and 69. Meanwhile, subcluster 3 consolidates all Enterococcus isolates, namely 121, 142, 144, and 152. Notably, isolate 29 takes a divergent path, forming a separate branch as subcluster 4. Cluster 2 gathers Streptomyces spp., and two subclusters can be observed: isolate 89 that exhibited excellent performance on PET substrate was closely grouped with Streptomyces sp. ALP3 (polyester degrader), while other Streptomyces isolates (19, 25, 134) that were more effective on PET monomers were grouped in a separate subcluster. The last cluster contains Pseudomonas spp., grouped together with the already well-known PET degrader I. sakaiensis, including our isolate 129, capable of using all PET-related substrates, especially TPA, where a halo zone was observed around colonies according to the screening experiment.
Our study reveals that 20 isolates consisted of four different genera, indicating great potential for species diversity among bacteria exhibiting the ability to degrade polymers. Notably, distinctive patterns emerge concerning the substrate range. While it is essential to approach the interpretation with caution due to site-specific limitations and sample size constraints, these findings are of significant importance in understanding the organisms responsible for the in situ degradation of plastics.
3.4. Upcycling Capabilities of Isolated Strains
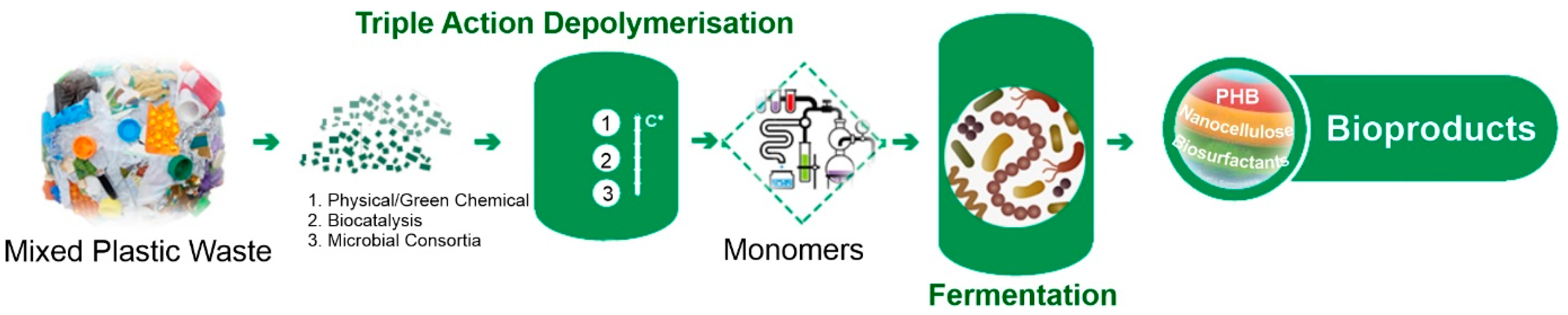
The primary aim of the BioICEP project is to showcase a sustainable pathway towards a circular economy for plastics. This involves the development of an advanced, energy-efficient, low-carbon, and cost-effective process for transforming waste plastics into high-demand bioproducts and bioplastics (Figure 4). In order to estimate if the selected isolates can be used for plastic waste conversion into high-value products, their emulsification activities and PHA production abilities were assessed. Moreover, pre-treated post-consumer PET (mixture of PET-related substrates) was used to assess PHA production.
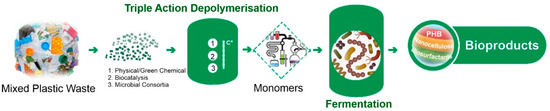
Figure 4.
BioICEP project overview.
3.4.1. Emulsification Assay
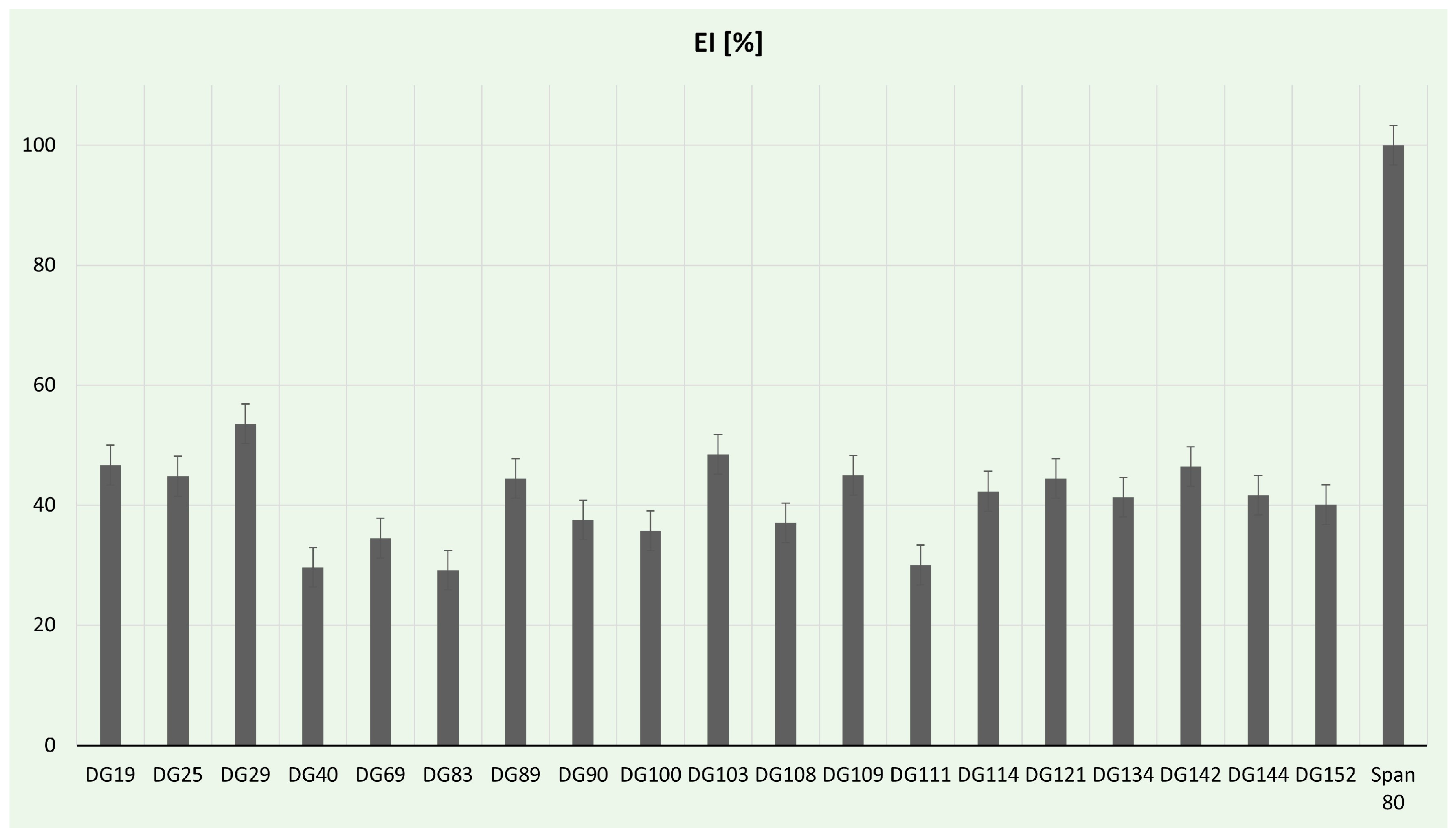
Biosurfactants are gaining prominence as potential natural surfactants, presenting numerous advantages compared to chemical surfactants. These benefits include low toxicity, inherent biodegradability, and ecological acceptability [89]. In our study, the emulsification activity of selected isolates was determined (Figure 5). In comparison to the positive control, the highest (53.57%) emulsification index was for Aneurinibacillus sp. DG29. These strains are already reported in the literature as promising biosurfactant producers. For example, A. aneurinilyticus was previously described as a potent extracellular and cell-bound lipopeptide producer by Lopez-Prieto et al. [90], while A. thermoaerophilus HAK01 isolated from municipal landfill sites was able to produce about 4.9 g/L of lipopeptide at 45 °C [91]. In this study, a notable emulsification index, over 40%, was achieved for Streptomyces spp., while the lowest values (below 30%) were obtained from Neobacillus sp. DG40, Bacillus murallis sp. DG83, and Bacillus sp. DG111. Streptomyces spp. has been reported as a successful biosurfactant producer in the literature. For example, Streptomyces enissocaesilis HRB1 isolated from the plastic-polluted site emerged as a promising source for the economical production of glycolipid biosurfactants, showcasing potential applications in both biomedical and environmental domains [92]. Zambry et al. reported Streptomyces sp. PBD-410L strain with an EI of 46.6% and the ability to effectively produce lipopeptide biosurfactant using palm oil as a sole carbon source [93]. The significance of developing green biosurfactant production methods cannot be overstated, especially considering the critical role of green strategies in the food and health sectors [94]. Utilizing food and plastic wastes can contribute to the generally safe production of biosurfactants; therefore, future studies will be focused on optimization processes, including carbon source exchange with plastic waste.
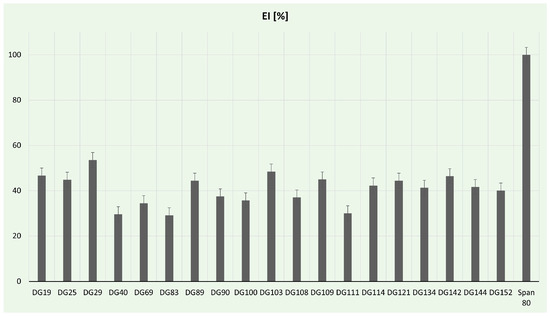
Figure 5.
Emulsification assay percentages for tested bacterial strains.
3.4.2. PHA Production
PHAs represent a promising alternative to certain petrochemical-based plastics. These polyesters are synthesized and stored within the cytoplasm of various bacteria and archaea as water-insoluble inclusions. Typically, the production of PHAs occurs when these microbes are cultured under conditions of nutrient limitation, specifically with low concentrations of nitrogen, phosphorus, sulphur, or oxygen, and ample carbon sources. Industries have strategically optimized fermentation conditions to minimize the cost of commercially producing PHAs. In industrial settings, these biodegradable polyesters are obtained through microbial fermentation processes, employing a variety of carbon sources [95,96]. Some examples include the use of discarded plant oils [96], molasses derived from the sugar industry [97], lignocellulosic materials [98], oil palm shells [99], pressed fruit fibre [100], biodiesel waste [101], and waste animal oils [102]. In this study, the ability of selected isolates to produce PHA using pre-treated post-consumer PET as a sole carbon source was examined. Fluorescence was detected in six isolates (Figure S1), and PHA presence and composition were determined by GC analysis (Table 3).
Table 3.
Evaluation of PHA materials obtained after cultivation of selected isolates using residues after PET reactive extrusion as a sole carbon source.
According to GC analysis, the highest biomass yield was obtained by Streptomyces sp. DG19 (1.39 g/L), followed by Priestia sp. DG69 (1.06 g/L). Priestia sp. DG69 and Neobacillus sp. DG40 were the most successful producers with 4.14 and 3.34% of PHA, respectively. The ability of these strains to produce PHAs has been previously recognized. For example, the ability of Priestia spp. to produce these molecules using sugarcane molasses as a sole carbon source has been reported, even without a sterilization process prior to inoculation [103,104]. In the study of Sivakuvamar et al., Streptomyces sp., isolated from the Bay of Bengal, proved to be a potential source of PHB, utilizing natural carbon sources, such as paddy straw, wheat bran, and rice bran [105]. Co-cultivation of these strains was also reported as successful in PHA production using plant biomass as a carbon source [106].
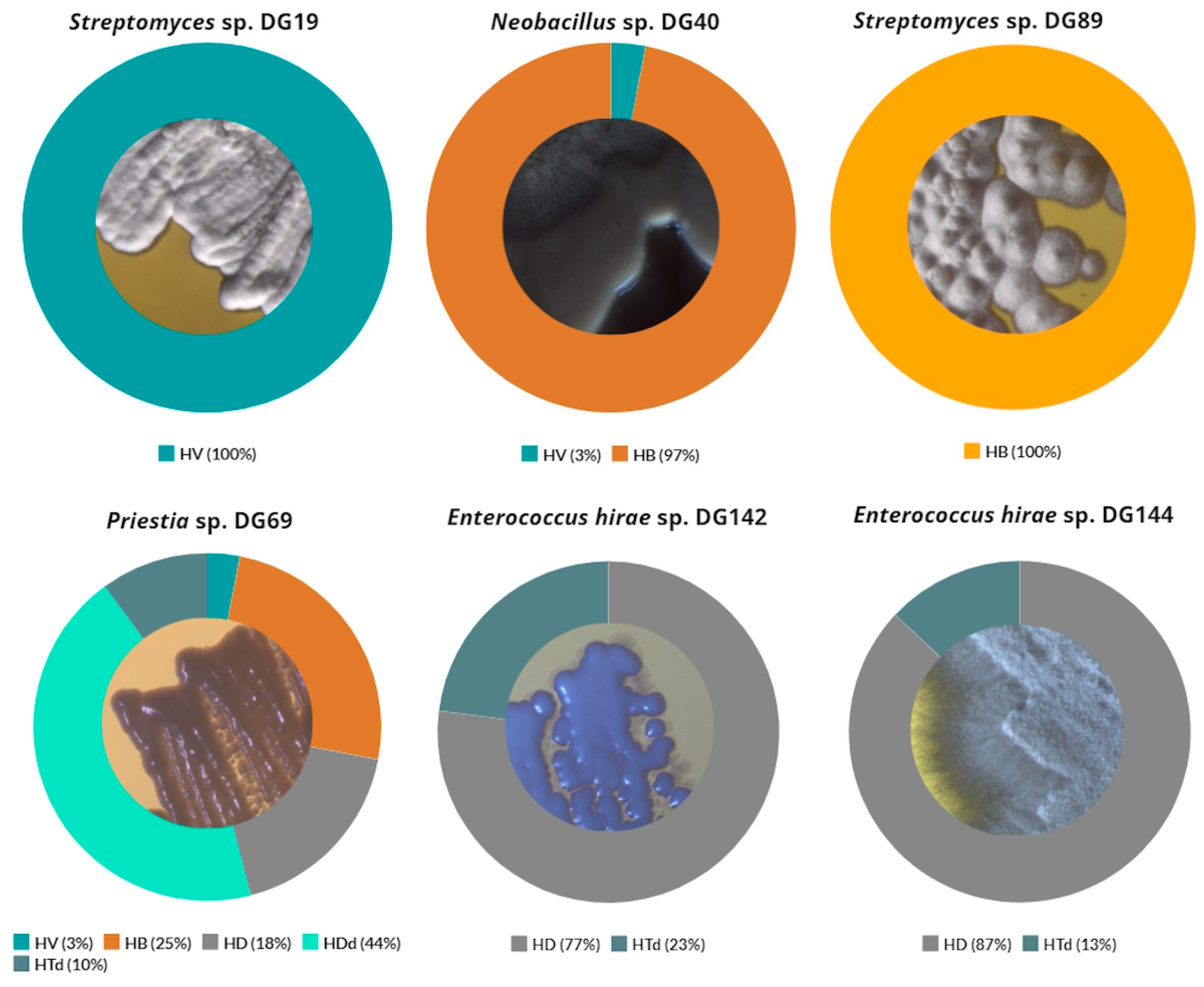
The composition of produced PHAs depends on several factors, including the type of microorganism, nature of carbon source, and culture conditions. In reality, numerous naturally occurring microorganisms have the capacity to produce PHAs with a diverse monomeric composition, influencing the material properties of the resulting PHA [107]. For this study, additional analysis revealed that a high percentage of 3-hydroxybutyrate (HB) monomers was detected for Neobacillus sp. DG40 and Priestia sp. DG69 strains: 97 and 25%, respectively (Figure 6). In addition to these, 3-hydroxyvalerate (HV) and 3-hydroxydecanoate (HD) monomers were also detected for these strains in lower percentages. A similar composition was reported by Kenny et al. for the examined Pseudomonas spp. [108], with a higher percentage of 3-hydroxypentadecanoate (HDd) in comparison to the results derived in this study. Strain Streptomyces sp. DG19 produced 100% of HV monomer, while high percentages of HD monomers were found after cultivation of Enterococcus hirae sp. DG142 and Enterococcus hirae sp. DG144: 77 and 87%, respectively. High percentages of these monomers were also observed during the cultivation of Ralstonia eutropha H16, 80% for HD and 90% of HV monomer, depending on the carbon source type [109].
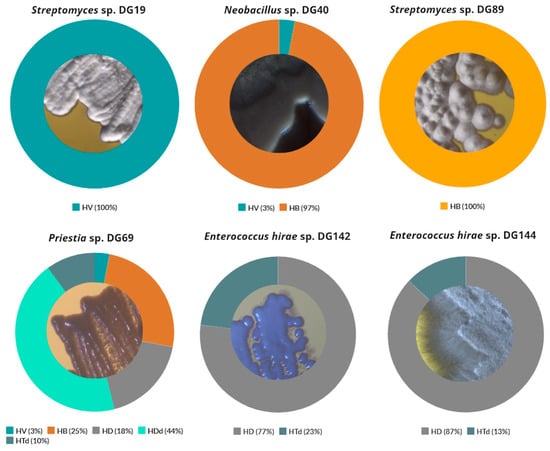
Figure 6.
Composition of specific PHA monomers detected after selected isolates cultivation. HB: polyhydroxybutyrate; HV: 3-hydroxyvalerate; HD: 3-hydroxydecanoate; HDd: 3-hydroxypentadecanoate; HTd: 3-hydroxyhexadecanoate.
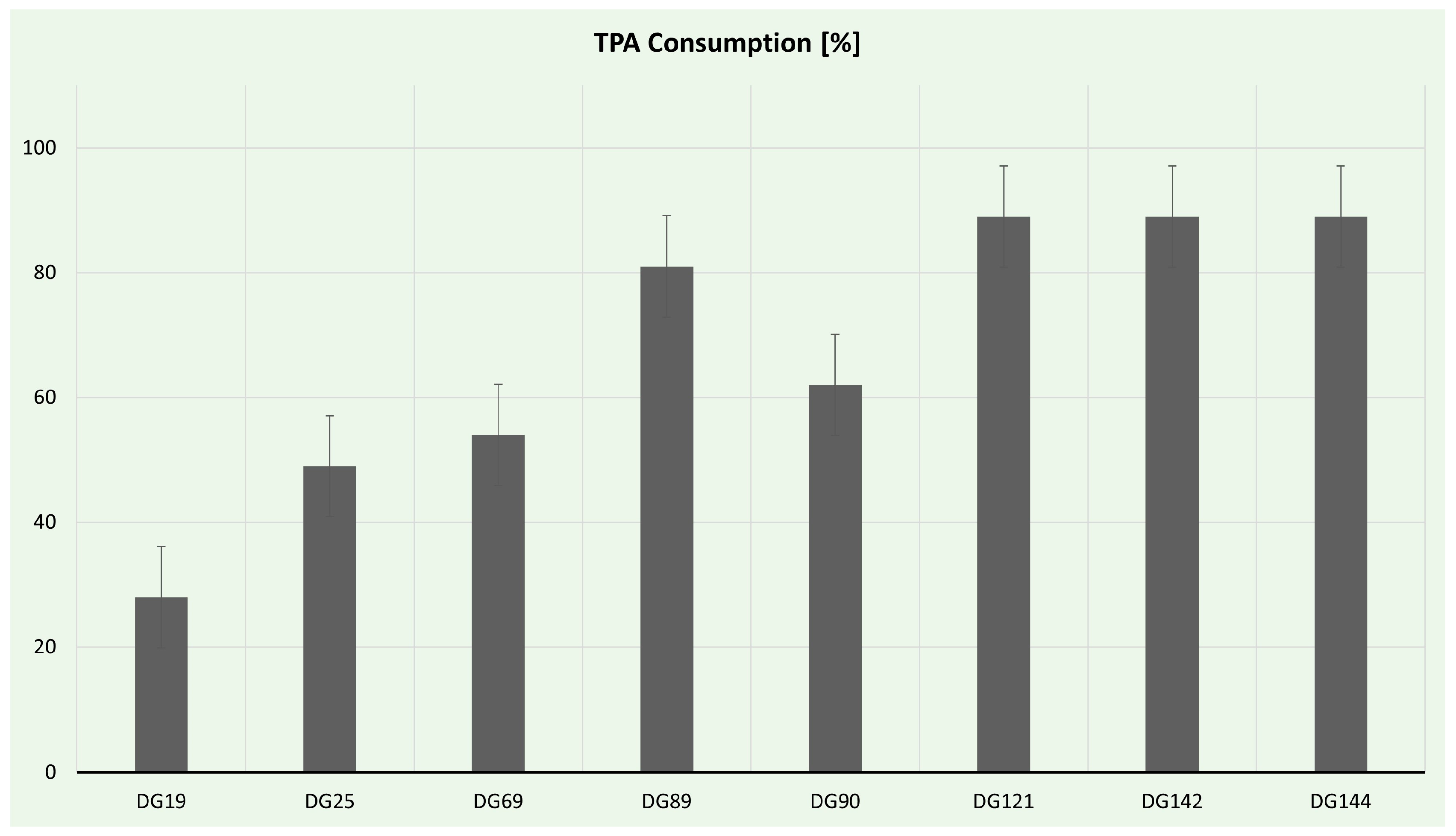
TPA consumption was estimated via HPLC analysis, and the results are presented in Figure 7. Eight isolates had the ability to use TPA as a sole carbon source in liquid culture; nevertheless, only five produced PHAs. The most effective strains for TPA utilization were Enterococcus strains. Enterococcus hirae spp. DG142 and DG144 consumed almost 90% of the offered TPA. Interestingly, Enterococcus hirae sp. DG121, despite being the only strain capable of forming clearing halos on TPA plates, consumed the same amount of TPA in the time frame examined. Although all of the isolated Enterococcus strains possess the necessary enzymatic machinery to utilize TPA, only strain DG121 can secrete these enzymes. Additionally, Streptomyces strains DG89 and DG90 consumed >60% of TPA, probably owing to the rich oxidoreduction enzyme arsenal of Streptomyces strains [110].
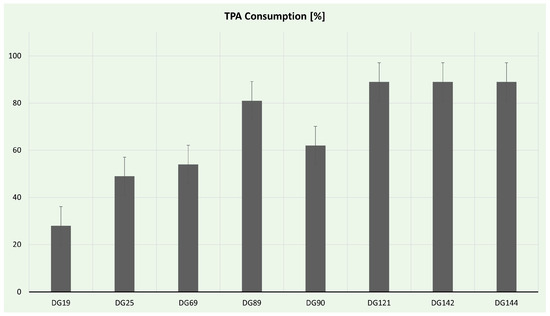
Figure 7.
Terephthalic acid consumption during selected isolate cultivation.
Recently there has been a surge in reports highlighting microbial strains with the capability to degrade plastic materials and transform them into valuable products, such as PHA. This development opens possibilities for advancing microbial and enzymatic technologies for the treatment of plastic waste, thereby contributing to the progress of plastics circularity. However, certain challenges persist, requiring additional research. A crucial aspect involves gaining a more profound comprehension of the various pathways of plastic degradation to enhance the development of advanced biotechnological techniques.
4. Conclusions
Plastic pollution poses a significant threat to ecosystems and human health, necessitating innovative solutions for its remediation. The findings presented in this research are crucial in advancing our understanding of how microorganisms can be harnessed to combat plastic pollution. By isolating and characterizing strains capable of degrading various plastic substrates, this study lays the foundation for the development of eco-friendly and sustainable strategies for plastic waste management. Several promising strains capable of degrading plastics, particularly PET and PU, were isolated (Table 4). These strains belong to the Bacillus, Enterococcus, Streptomyces, and Pseudomonas genera. Phylogenetic analysis demonstrated the diversity of these strains across the microbial tree of life. Additionally, emulsification activity assessment indicated their biosurfactant-producing capabilities making them potentially valuable for various applications, including bioremediation and the food and health industries. Finally, some of the strains exhibited significant potential for PHA production from pre-treated post-consumer PET, contributing to the circular economy for plastics.
Table 4.
The best-performing isolates’ performance and characteristics.
This research provides valuable insights into the microbial degradation of plastics, offering a sustainable pathway for plastic waste management. In summary, this research has broad implications for the fields of environmental science, biotechnology, and sustainable waste management, offering a path toward an eco-conscious and resilient future.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms11122914/s1, Figure S1: Visualization of PHA granules in isolates via confocal microscopy.
Author Contributions
Conceptualization, M.M., M.B.F. and J.N.-R.; methodology, D.A.G.H., B.P., A.J., C.T. and M.B.; validation, B.P., C.C., F.F., C.T., P.M. and M.M.; formal analysis, D.A.G.H., A.J. and M.B.; investigation, B.P. and M.M.; data curation, D.A.G.H., A.J., C.C., C.T. and M.B.; writing—original draft preparation, M.M. and D.A.G.H.; writing—review and editing, M.M., J.N.-R., F.F. and P.M.; visualization, M.M.; supervision, M.M., J.N.-R., P.M. and M.B.F.; project administration, M.M.; funding acquisition, M.B.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Technological University of The Shannon through the President Seed Fund, the Government of Ireland International Education Scholarship 2020/2021, the European Union’s Horizon 2020 Research and Innovation program (grant number: 870292 (BioICEP)); European Union’s Horizon Europe EIC Pathfinder program (grant number: 101046758 (EcoPlastiC)). This work was supported by national funds from FCT—Fundação para a Ciência e a Tecnologia, I.P. (Portugal), in the scope of projects UIDP/04378/2020 and UIDB/04378/2020 of the Research Unit on Applied Molecular Biosciences—UCIBIO and LA/P/0140/2020 of the Associate Laboratory Institute for Health and Bioeconomy—i4HB.
Data Availability Statement
Data are contained within the article and supplementary materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Purohit, J.; Chattopadhyay, A.; Teli, B. Metagenomic Exploration of Plastic Degrading Microbes for Biotechnological Application. Curr. Genom. 2020, 21, 253–270. [Google Scholar] [CrossRef] [PubMed]
- Gambarini, V.; Pantos, O.; Kingsbury, J.M.; Weaver, L.; Handley, K.M.; Lear, G. Phylogenetic Distribution of Plastic-Degrading Microorganisms. mSystems 2021, 6. [Google Scholar] [CrossRef]
- Thumarat, U.; Nakamura, R.; Kawabata, T.; Suzuki, H.; Kawai, F. Biochemical and genetic analysis of a cutinase-type polyesterase from a thermophilic Thermobifida alba AHK119. Appl. Microbiol. Biotechnol. 2011, 95, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Oeser, T.; Then, J.; Kühn, N.; Barth, M.; Schmidt, J.; Zimmermann, W. Functional characterization and structural modeling of synthetic polyester-degrading hydrolases from Thermomonospora curvata. AMB Express 2014, 4, 44. [Google Scholar] [CrossRef]
- Tanasupawat, S.; Takehana, T.; Yoshida, S.; Hiraga, K.; Oda, K. Ideonella sakaiensis sp. nov., isolated from a microbial consortium that degrades PET. Int. J. Syst. Evol. Microbiol. 2016, 66, 2813–2818. [Google Scholar] [CrossRef] [PubMed]
- Egmond, M. Fusarium solani pisi cutinase. Biochimie 2000, 82, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Ronkvist, Å.M.; Xie, W.; Lu, W.; Gross, R.A. Cutinase-Catalyzed Hydrolysis of Poly(ethylene terephthalate). Macromolecules 2009, 42, 5128–5138. [Google Scholar] [CrossRef]
- Maeda, H.; Yamagata, Y.; Abe, K.; Hasegawa, F.; Machida, M.; Ishioka, R.; Gomi, K.; Nakajima, T. Purification and characterization of a biodegradable plastic-degrading enzyme from Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2005, 67, 778–788. [Google Scholar] [CrossRef]
- Da Costa, J.P.; Nunes, A.R.; Santos, P.S.M.; Girão, A.V.; Duarte, A.C.; Rocha-Santos, T. Degradation of polyethylene microplastics in seawater: Insights into the environmental degradation of polymers. J. Environ. Sci. Health Part A 2018, 53, 866–875. [Google Scholar] [CrossRef]
- Song, L.; Wang, Y.; Tang, W.; Lei, Y. Bacterial community diversity in municipal waste landfill sites. Appl. Microbiol. Biotechnol. 2015, 99, 7745–7756. [Google Scholar] [CrossRef]
- Kumar, R.; Pandit, P.; Kumar, D.; Patel, Z.; Pandya, L.; Kumar, M.; Joshi, C.; Joshi, M. Landfill microbiome harbour plastic degrading genes: A metagenomic study of solid waste dumping site of Gujarat, India. Sci. Total Environ. 2021, 779, 146184. [Google Scholar] [CrossRef] [PubMed]
- Hiraga, K.; Taniguchi, I.; Yoshida, S.; Kimura, Y.; Oda, K. Biodegradation of waste PET. EMBO Rep. 2019, 20, e49365. [Google Scholar] [CrossRef] [PubMed]
- Malik, N.; Lakhawat, S.S.; Kumar, V.; Sharma, V.; Bhatti, J.S.; Sharma, P.K. Recent advances in the omics-based assessment of microbial consortia in the plastisphere environment: Deciphering the dynamic role of hidden players. Process Saf. Environ. Prot. 2023, 176, 207–225. [Google Scholar] [CrossRef]
- Graham Bowditch, T. Penetration of Polyvinyl Chloride and Polypropylene Packaging Films by Ephestia cautella (Lepidoptera: Pyralidae) and Plodia interpunctella (Lepidoptera: Pyralidae) Larvae, and Tribolium confusum (Coleoptera: Tenebrionidae) Adults. J. Econ. Entomol. 1997, 90, 1028–1031. [Google Scholar] [CrossRef]
- Kim, J.-H.; Lee, S.-H.; Lee, B.-M.; Son, K.-H.; Park, H.-Y. Biodegradation Potential of Polyethylene Terephthalate by the Two Insect Gut Symbionts Xanthomonas sp. HY-74 and Bacillus sp. HY-75. Polymers 2023, 15, 3546. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.Q.; Longing, S.; Siebecker, M.G. Consumption and degradation of different consumer plastics by mealworms (Tenebrio molitor): Effects of plastic type, time, and mealworm origin. J. Clean. Prod. 2023, 403, 136842. [Google Scholar] [CrossRef]
- Ong, S.Y.; Heng, K.-S.; Heng, K.-S.; Ong, S.Y.; Sudesh, K. Efficient biosynthesis and recovery of polyhydroxyalkanoate. Malays. J. Microbiol. 2016, 12, 383–398. [Google Scholar]
- Paul-Pont, I.; Ghiglione, J.-F.; Gastaldi, E.; Ter Halle, A.; Huvet, A.; Bruzaud, S.; Lagarde, F.; Galgani, F.; Duflos, G.; George, M.; et al. Discussion about suitable applications for biodegradable plastics regarding their sources, uses and end of life. Waste Manag. 2023, 157, 242–248. [Google Scholar] [CrossRef]
- Nielsen, C.; Rahman, A.; Rehman, A.U.; Walsh, M.K.; Miller, C.D. Food waste conversion to microbial polyhydroxyalkanoates. Microb. Biotechnol. 2017, 10, 1338–1352. [Google Scholar] [CrossRef]
- Surendran, A.; Lakshmanan, M.; Chee, J.Y.; Sulaiman, A.M.; Thuoc, D.V.; Sudesh, K. Can Polyhydroxyalkanoates Be Produced Efficiently From Waste Plant and Animal Oils? Front. Bioeng. Biotechnol. 2020, 8, 169. [Google Scholar] [CrossRef]
- Mannina, G.; Presti, D.; Montiel-Jarillo, G.; Carrera, J.; Suárez-Ojeda, M.E. Recovery of polyhydroxyalkanoates (PHAs) from wastewater: A review. Bioresour. Technol. 2020, 297, 122478. [Google Scholar] [CrossRef]
- Tiso, T.; Narancic, T.; Wei, R.; Pollet, E.; Beagan, N.; Schröder, K.; Honak, A.; Jiang, M.; Kenny, S.T.; Wierckx, N.; et al. Towards bio-upcycling of polyethylene terephthalate. Metab. Eng. 2021, 66, 167–178. [Google Scholar] [CrossRef]
- Johnston, B.; Adamus, G.; Ekere, A.I.; Kowalczuk, M.; Tchuenbou-Magaia, F.; Radecka, I. Bioconversion of Plastic Waste Based on Mass Full Carbon Backbone Polymeric Materials to Value-Added Polyhydroxyalkanoates (PHAs). Bioengineering 2022, 9, 432. [Google Scholar] [CrossRef] [PubMed]
- Wierckx, N.; Prieto, M.A.; Pomposiello, P.; de Lorenzo, V.; O’Connor, K.; Blank, L.M. Plastic waste as a novel substrate for industrial biotechnology. Microb. Biotechnol. 2015, 8, 900–903. [Google Scholar] [CrossRef]
- Johnston, B.; Radecka, I.; Hill, D.; Chiellini, E.; Ilieva, V.I.; Sikorska, W.; Musioł, M.; Zięba, M.; Marek, A.A.; Keddie, D.; et al. The Microbial Production of Polyhydroxyalkanoates from Waste Polystyrene Fragments Attained Using Oxidative Degradation. Polymers 2018, 10, 957. [Google Scholar] [CrossRef] [PubMed]
- Narancic, T.; Salvador, M.; Hughes, G.M.; Beagan, N.; Abdulmutalib, U.; Kenny, S.T.; Wu, H.; Saccomanno, M.; Um, J.; O’Connor, K.E.; et al. Genome analysis of the metabolically versatile Pseudomonas umsongensis GO16: The genetic basis for PET monomer upcycling into polyhydroxyalkanoates. Microb. Biotechnol. 2021, 14, 2463–2480. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.M.; Machado, I.; Nicolau, A.; Pereira, M.O. Improvements on colony morphology identification towards bacterial profiling. J. Microbiol. Methods 2013, 95, 327–335. [Google Scholar] [CrossRef]
- Breakwell, D.; MacDonald, B.; Woolverton, C.; Smith, K.; Robison, R. Colony Morphology Protocol. Am. Soc. Microbiol. 2007. Available online: https://asm.org/ASM/media/Protocol-Images/Colony-Morphology-Protocol.pdf?ext=.pdf (accessed on 15 March 2023).
- Molitor, R.; Bollinger, A.; Kubicki, S.; Loeschcke, A.; Jaeger, K.-E.; Thies, S. Agar plate-based screening methods for the identification of polyester hydrolysis by Pseudomonas species. Microb. Biotechnol. 2020, 13, 274–284. [Google Scholar] [CrossRef]
- Ibrahim MH, A.; Willems, A.; Steinbüchel, A. Isolation and characterization of new poly(3HB)-accumulating star-shaped cell-aggregates-forming thermophilic bacteria. J. Appl. Microbiol. 2010, 109, 1579–1590. [Google Scholar] [CrossRef]
- Álvarez-Barragán, J.; Domínguez-Malfavón, L.; Vargas-Suárez, M.; González-Hernández, R.; Aguilar-Osorio, G.; Loza-Tavera, H. Biodegradative Activities of Selected Environmental Fungi on a Polyester Polyurethane Varnish and Polyether Polyurethane Foams. Appl. Environ. Microbiol. 2016, 82, 5225–5235. [Google Scholar] [CrossRef]
- Nikodinovic, J.; Barrow, K.D.; Chuck, J.-A. High yield preparation of genomic DNA from Streptomyces. Biotechniques 2003, 35, 932–936. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Saltou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Brennan Fournet, M.; Attallah, O.A.; Mojicevic, M.; Chen, Y.; Major, I. High Throughput Mechanochemical Depolymerisation and Purification of Polyethylene Terephthalate and Related Polymers; World Intellectual Property Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Silvestre, S.L.; Araújo, D.; Marques, A.C.; Pires, C.; Matos, M.; Alves, V.D.; Martins, R.F.P.; Freitas, F.; Reis, M.A.M.; Fortunato, E. Microneedle Arrays of Polyhydroxyalkanoate by Laser-Based Micromolding Technique. ACS Appl. Bio Mater. 2020, 3, 5856–5864. [Google Scholar] [CrossRef]
- Rebocho, A.T.; Perreira, J.R.; Neves, L.A.; Alves, V.D.; Sevrin, C.; Grandfils, C.; Freitas, F.; Reis, M.A.M. Preparation and Characterization of Films Based on a Natural P(3HB)/mcl-PHA Blend Obtained through the Co-culture of Cupriavidus Necator and Pseudomonas Citronellolis in Apple Pulp Waste. Bioengineering 2020, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Bleuven, C.; Landry, C.R. Molecular and cellular bases of adaptation to a changing environment in microorganisms. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161458. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, I. Role of microbiome and biofilm in environmental plastic degradation. Biocatal. Agric. Biotechnol. 2022, 39, 102263. [Google Scholar] [CrossRef]
- Hara, H.; Eltis, L.D.; Davies, J.E.; Mohn, W.W. Transcriptomic Analysis Reveals a Bifurcated Terephthalate Degradation Pathway in Rhodococcus sp. Strain RHA1. J. Bacteriol. 2007, 189, 1641–1647. [Google Scholar] [CrossRef] [PubMed]
- Mooney, A.; Ward, P.G.; O’Connor, K.E. Microbial degradation of styrene: Biochemistry, molecular genetics, and perspectives for biotechnological applications. Appl. Microbiol. Biotechnol. 2006, 72, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Huang, D.; Ji, J.; Völker, C.; Wurm, F.R. Seawater-Degradable Polymers—Fighting the Marine Plastic Pollution. Adv. Sci. 2021, 8, 2001121. [Google Scholar] [CrossRef]
- Mitra, B.; Das, A. The Ability of Insects to Degrade Complex Synthetic Polymers. In Arthropods-New Advances and Perspectives [Working Title]; IntechOpen: London, UK, 2023. [Google Scholar] [CrossRef]
- Kopecká, R.; Kubínová, I.; Sovová, K.; Mravcová, L.; Vítěz, T.; Vítězová, M. Microbial degradation of virgin polyethylene by bacteria isolated from a landfill site. SN Appl. Sci. 2022, 4, 302. [Google Scholar] [CrossRef]
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J.P.; Klenk, H.P.; Clément, C.; Ouhdouch, Y.; Van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef]
- Rodríguez-Fonseca, M.F.; Sánchez-Suárez, J.; Valero, M.F.; Ruiz-Balaguera, S.; Díaz, L.E. Streptomyces as Potential Synthetic Polymer Degraders: A Systematic Review. Bioengineering 2021, 8, 154. [Google Scholar] [CrossRef]
- Taddei, A.; Rodríguez, M.J.; Márquez-Vilchez, E.; Castelli, C. Isolation and identification of Streptomyces spp. from Venezuelan soils: Morphological and biochemical studies. I. Microbiol. Res. 2006, 161, 222–231. [Google Scholar] [CrossRef]
- Janakiev, T.; Milošević, K.; Petrović, M.; Miljković, J.; Stanković, N.; Zdravković, D.S.; Dimkić, I. Chironomus riparius Larval Gut Bacteriobiota and Its Potential in Microplastic Degradation. Microb. Ecol. 2023, 86, 1909–1922. [Google Scholar] [CrossRef]
- Bergey, H.D. Bergey’s Manual of Determinative Bacteriology; Williams & Wilkins Co.: Philadelphia, PA, USA, 1975. [Google Scholar]
- Vary, P.S.; Biedendieck, R.; Fuerch, T.; Meinhardt, F.; Rohde, M.; Deckwer, W.-D.; Jahn, D. Bacillus megaterium—From simple soil bacterium to industrial protein production host. Appl. Microbiol. Biotechnol. 2007, 76, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Meng, T.K.; Kassim, A.S.B.M.; Razak, A.H.B.A.; Fauzi, N.A.B.M. Bacillus megaterium: A Potential and an Efficient Bio-Degrader of Polystyrene. Braz. Arch. Biol. Technol. 2021, 64, e21190321. [Google Scholar] [CrossRef]
- Biedendieck, R.; Knuuti, T.; Moore, S.J.; Jahn, D. The “beauty in the beast”—The multiple uses of Priestia megaterium in biotechnology. Appl. Microbiol. Biotechnol. 2021, 105, 5719–5737. [Google Scholar] [CrossRef]
- Zerhouni, K.; Abbouni, B.; Kanoun, K.; Larbi Daouadji, K.; Tifrit, A.; Benahmed, M.; Chaouche, T.M. Isolation and identification of low density polythene-degrading bacteria from soil of North West of Algeria. South Asian J. Exp. Biol. 2019, 8, 76–82. [Google Scholar] [CrossRef]
- Puglisi, E.; Romaniello, F.; Galletti, S.; Boccaleri, E.; Frache, A.; Cocconcelli, P.S. Selective bacterial colonization processes on polyethylene waste samples in an abandoned landfill site. Sci. Rep. 2019, 9, 14138. [Google Scholar] [CrossRef] [PubMed]
- Saikia, S.S.; Borah, B.K.; Baruah, G.; Rokozeno; Deka, M.K. Characterization of the gut microbes of greater wax moth (Galleria mellonella Linnaeus) shows presence of potential polymer degraders. Folia Microbiol. 2022, 67, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Kag, S.; Kumar, P.; Kataria, R. Potato Peel Waste as an Economic Feedstock for PHA Production by Bacillus circulans. Appl. Biochem. Biotechnol. Appl. Biochem. Biotechnol. 2023, 1–15. [Google Scholar] [CrossRef]
- Phukon, P.; Saikia, J.P.; Konwar, B.K. Bio-plastic (P-3HB-co-3HV) from Bacillus circulans (MTCC 8167) and its biodegradation. Colloids Surf B Biointerfaces 2012, 92, 30–34. [Google Scholar] [CrossRef]
- Nakkabi, A.; Noureddine, E.; Moulay, S.; Ibnsouda, K.S.; Mohammed, F.F. Biodegradation of Poly (Ethylene Terephthalate) by Bacillus subtilis. Int. J. Recent. Adv. Multidiscip. Res. 2015, 2, 1060–1062. [Google Scholar]
- Benavides Fernández, C.D.; Guzmán Castillo, M.P.; Quijano Pérez, S.A.; Carvajal Rodríguez, L.V. Microbial degradation of polyethylene terephthalate: A systematic review. SN Appl. Sci. 2022, 4, 263. [Google Scholar] [CrossRef]
- Li, W.; Jia, M.-X.; Deng, J.; Wang, J.-H.; Lin, Q.-L.; Liu, C.; Wang, S.-S.; Tang, J.-X.; Zeng, X.-X.; Ma, L.; et al. Isolation, genetic identification and degradation characteristics of COD-degrading bacterial strain in slaughter wastewater. Saudi J. Biol. Sci. 2018, 25, 1800–1805. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, S.; Dang, G.; Jia, R.; Chen, S.; Deng, X.; Liu, G.; Beckers, Y.; Cai, H. Screening and characterization of Bacillus velezensis LB-Y-1 toward selection as a potential probiotic for poultry with multi-enzyme production property. Front. Microbiol. 2023, 14, 1143265. [Google Scholar] [CrossRef] [PubMed]
- Gui, Z.; Liu, G.; Liu, X.; Cai, R.; Liu, R.; Sun, C. A Deep-Sea Bacterium Is Capable of Degrading Polyurethane. Microbiol. Spectr. 2023, 11, e0007323. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Chen, C.; Shen, Q.; Rosen, B.P.; Zhao, F.-J. Genetically Engineering Bacillus subtilis with a Heat-Resistant Arsenite Methyltransferase for Bioremediation of Arsenic-Contaminated Organic Waste. Appl. Environ. Microbiol. 2015, 81, 6718–6724. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Zimmermann, W. Microbial enzymes for the recycling of recalcitrant petroleum-based plastics: How far are we? Microb. Biotechnol. 2017, 10, 1308–1322. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Seong, H.J.; Jang, Y.-S. Degradation of low density polyethylene by Bacillus species. Appl. Biol. Chem. 2022, 65, 84. [Google Scholar] [CrossRef]
- Nakkabi, A.; Sadiki, M.; Fahim, M.; Ittobane, N.; IbnsoudaKoraichi, S.; Barkai, H.; El Abed, S. Biodegradation of Poly(ester urethane)s by Bacillus subtilis. Int. J. Environ. Res. 2015, 9, 157–162. [Google Scholar]
- Mukherjee, S.; Roy Chowdhuri, U.; Kundu, P.P. Bio-degradation of polyethylene waste by simultaneous use of two bacteria: Bacillus licheniformis for production of bio-surfactant and Lysinibacillus fusiformis for bio-degradation. RSC Adv. 2016, 6, 2982–2992. [Google Scholar] [CrossRef]
- Arena, M.; Abbate, C.; Fukushima, K.; Gennari, M. Degradation of poly (lactic acid) and nanocomposites by Bacillus licheniformis. Environ. Sci. Pollut. Res. 2011, 18, 865–870. [Google Scholar] [CrossRef]
- Jain, K.; Bhunia, H.; Reddy, M.S. Degradation of polypropylene-poly-L-lactide blends by Bacillus isolates: A microcosm and field evaluation. Bioremediation J. 2022, 26, 64–75. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, T.; Zhang, W.; Lin, J.; Wang, Z.; Lyu, S.; Tong, H. Biodegradation of polylactic acid by a mesophilic bacteria Bacillus safensis. Chemosphere 2023, 318, 137991. [Google Scholar] [CrossRef]
- Abdullah, A.; Waqas, M.; Haris, M.; Asim, N.; Islam, H.U.; Khan, A.; Khattak, H.; Ali, S. Biodegradable Potential of Bacillus amyloliquefaciens and Bacillus safensis Using Low Density Polyethylene Thermoplastic (LDPE) Substrate. Eur. J. Environ. Public Health 2021, 5, em0069. [Google Scholar] [CrossRef] [PubMed]
- Oulidi, O.; Nakkabi, A.; Bouymajane, A.; Elaraaj, I.; Filali, F.R.; Fahim, M.; El Moualij, N. Biodegradation of polyamide 6 by Lysinibacillus sp, Alcaligene faecalis and Enterococcus faecalis. Clean. Chem. Eng. 2022, 3, 100054. [Google Scholar] [CrossRef]
- Hchaichi, I.; Bandini, F.; Spini, G.; Banni, M.; Cocconcelli, P.S.; Puglisi, E. Enterococcus faecalis and Vibrio harveyi colonize low-density polyethylene and biodegradable plastics under marine conditions. FEMS Microbiol. Lett. 2020, 367, fnaa125. [Google Scholar] [CrossRef] [PubMed]
- Nur, C.; Guven, O. Poly--hydroxybutyrate (PHB) production from domestic wastewater using Enterobacter aerogenes 12Bi strain. Afr. J. Microbiol. Res. 2011, 5, 690–702. [Google Scholar]
- Bhuwal, A.K.; Singh, G.; Aggarwal, N.K.; Goyal, V.; Yadav, A. Isolation and Screening of Polyhydroxyalkanoates Producing Bacteria from Pulp, Paper, and Cardboard Industry Wastes. Int. J. Biomater. 2013, 2013, 752821. [Google Scholar] [CrossRef]
- Obradors, N.; Aguilar, J. Efficient biodegradation of high-molecular-weight polyethylene glycols by pure cultures of Pseudomonas stutzeri. Appl. Environ. Microbiol. 1991, 57, 2383–2388. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, T.; Zhen, Y.; Li, Q.; Liang, Q.; Qi, Q. Screening and genome analysis of a Pseudomonas stutzeri that degrades PET monomer terephthalate. Acta Microbiol. Sin. 2022, 62, 200–212. [Google Scholar]
- Hooda, S.; Annu; Mondal, P. Insights into the degradation of high-density polyethylene microplastics using microbial strains: Effect of process parameters, degradation kinetics and modeling. Waste Manag. 2023, 164, 143–153. [Google Scholar] [CrossRef]
- Uefuji, M.; Kasuya, K.; Doi, Y. Enzymatic degradation of poly[(R)-3-hydroxybutyrate]: Secretion and properties of PHB depolymerase from Pseudomonas stutzeri. Polym. Degrad. Stab. 1997, 58, 275–281. [Google Scholar] [CrossRef]
- Drzyzga, O.; Prieto, A. Plastic waste management, a matter for the ‘community’. Microb. Biotechnol. 2019, 12, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Fields, R.D.; Rodriguez, F.; Finn, R.K. Microbial degradation of polyesters: Polycaprolactone degraded by P. pullulans. J. Appl. Polym. Sci. 1974, 18, 3571–3579. [Google Scholar] [CrossRef]
- Shah, Z.; Krumholz, L.; Aktas, D.F.; Hasan, F.; Khattak, M.; Shah, A.A. Degradation of polyester polyurethane by a newly isolated soil bacterium, Bacillus subtilis strain MZA-75. Biodegradation 2013, 24, 865–877. [Google Scholar] [CrossRef] [PubMed]
- Shah, Z.; Hasan, F.; Krumholz, L.; Aktas, D.F.; Shah, A.A. Degradation of polyester polyurethane by newly isolated Pseudomonas aeruginosa strain MZA-85 and analysis of degradation products by GC–MS. Int. Biodeterior. Biodegradation 2013, 77, 114–122. [Google Scholar] [CrossRef]
- Sriyapai, P.; Chansiri, K.; Sriyapai, T. Isolation and Characterization of Polyester-Based Plastics-Degrading Bacteria from Compost Soils. Microbiology 2018, 87, 290–300. [Google Scholar] [CrossRef]
- Liu, G.; Zhong, H.; Yang, X.; Liu, Y.; Shao, B.; Liu, Z. Advances in applications of rhamnolipids biosurfactant in environmental remediation: A review. Biotechnol. Bioeng. 2018, 115, 796–814. [Google Scholar] [CrossRef]
- López-Prieto, A.; Rodríguez-López, L.; Rincón-Fontán, M.; Cruz, J.M.; Moldes, A.B. Characterization of extracellular and cell bound biosurfactants produced by Aneurinibacillus aneurinilyticus isolated from commercial corn steep liquor. Microbiol. Res. 2021, 242, 126614. [Google Scholar] [CrossRef]
- Hajfarajollah, H.; Mokhtarani, B.; Tohidi, A.; Bazsefidpar, S.; Akbari Noghabi, K. Overproduction of lipopeptide biosurfactant by Aneurinibacillus thermoaerophilus HAK01 in various fed-batch modes under thermophilic conditions. RSC Adv. 2019, 9, 30419–30427. [Google Scholar] [CrossRef]
- Manikkasundaram, V.; Baskaran, A.; Kaari, M.; Angamuthu, V.; Venugopal, G.; Manikkam, R. Production and characterization of glycolipid biosurfactant from Streptomyces enissocaesilis HRB1 and its evaluation for biomedical and bioremediation applications. J. Surfactants Deterg. 2023, 26, 491–503. [Google Scholar] [CrossRef]
- Zambry, N.S.; Rusly, N.S.; Awang, M.S.; Md Noh, N.A.; Yahya AR, M. Production of lipopeptide biosurfactant in batch and fed-batch Streptomyces sp. PBD-410L cultures growing on palm oil. Bioprocess. Biosyst. Eng. 2021, 44, 1577–1592. [Google Scholar] [CrossRef]
- Mgbechidinma, C.L.; Akan, O.D.; Zhang, C.; Huang, M.; Linus, N.; Zhu, H.; Wakil, S.M. Integration of green economy concepts for sustainable biosurfactant production—A review. Bioresour. Technol. 2022, 364, 128021. [Google Scholar] [CrossRef]
- Bugnicourt, E.; Cinelli, P.; Lazzeri, A.; Alvarez, V. Polyhydroxyalkanoate (PHA): Review of synthesis, characteristics, processing and potential applications in packaging. Express Polym. Lett. 2014, 8, 791–808. [Google Scholar] [CrossRef]
- Akiyama, M.; Tsuge, T.; Doi, Y. Environmental life cycle comparison of polyhydroxyalkanoates produced from renewable carbon resources by bacterial fermentation. Polym. Degrad. Stab. 2003, 80, 183–194. [Google Scholar] [CrossRef]
- Kingsly, J.S.; Chathalingath, N.; Parthiban, S.A.; Sivakumar, D.; Sambugan, S.; Senniyappan, V.; Sekar, D.V.; Jasmine, A.H.; Gunasekar, G. Utilization of sugarcane molasses as the main carbon source for the production of polyhydroxyalkanoates from Enterobacter cloacae. Energy Nexus 2022, 6, 100071. [Google Scholar] [CrossRef]
- Obruca, S.; Benesova, P.; Marsalek, L.; Marova, I. Use of Lignocellulosic Materials for PHA Production. Chem. Biochem. Eng. Q. 2015, 29, 135–144. [Google Scholar] [CrossRef]
- Hassan, M.A.; Yee, L.-N.; Yee, P.L.; Ariffin, H.; Raha, A.R.; Shirai, Y.; Sudesh, K. Sustainable production of polyhydroxyalkanoates from renewable oil-palm biomass. Biomass Bioenergy 2013, 50, 1–9. [Google Scholar] [CrossRef]
- Asyraf, M.; Ishak, M.; Syamsir, A.; Nurazzi, N.; Sabaruddin, F.; Shazleen, S.; Norrrahim, M.; Rafidah, M.; Ilyas, R.; Rashid, M.Z.A.; et al. Mechanical properties of oil palm fibre-reinforced polymer composites: A review. J. Mater. Res. Technol. 2022, 17, 33–65. [Google Scholar] [CrossRef]
- Chanasit, W.; Hodgson, B.; Sudesh, K.; Umsakul, K. Efficient production of polyhydroxyalkanoates (PHAs) from Pseudomonas mendocina PSU using a biodiesel liquid waste (BLW) as the sole carbon source. Biosci. Biotechnol. Biochem. 2016, 80, 1440–1450. [Google Scholar] [CrossRef]
- Van Thuoc, D.; My, D.N.; Loan, T.T.; Sudesh, K. Utilization of waste fish oil and glycerol as carbon sources for polyhydroxyalkanoate production by Salinivibrio sp. M318. Int. J. Biol. Macromol. 2019, 141, 885–892. [Google Scholar] [CrossRef]
- Esmael, M.E.; Ibrahim, M.I.; Aldhumri, S.A.; Bayoumi, R.A.; Matsuo, K.; Khattab, A.M. Lipid-membranes interaction, structural assessment, and sustainable production of polyhydroxyalkanoate by Priestia filamentosa AZU-A6 from sugarcane molasses. Int. J. Biol. Macromol. 2023, 242, 124721. [Google Scholar] [CrossRef]
- Jung, H.J.; Kim, S.H.; Shin, N.; Oh, S.-J.; Hwang, J.H.; Kim, H.J.; Kim, Y.-H.; Bhatia, S.K.; Jeon, J.-M.; Yoon, J.-J.; et al. Polyhydroxybutyrate (PHB) production from sugar cane molasses and tap water without sterilization using novel strain, Priestia sp. YH4. Int. J. Biol. Macromol. 2023, 250, 126152. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Chinnadurai, G.S.; Perumal, P. Polyhydroxybutyrate by Streptomyces sp.: Production and characterization. Int. J. Biol. Macromol. 2017, 104, 1165–1171. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Fox, B.G.; Takasuka, T.E. Consolidated bioprocessing of plant biomass to polyhydroxyalkanoate by co-culture of Streptomyces sp. SirexAA-E and Priestia megaterium. Bioresour. Technol. 2023, 376, 128934. [Google Scholar] [CrossRef]
- Koller, M. Switching from petro-plastics to microbial polyhydroxyalkanoates (PHA): The biotechnological escape route of choice out of the plastic predicament? Eurobiotech J. 2019, 3, 32–44. [Google Scholar] [CrossRef]
- Kenny, S.T.; Runic, J.N.; Kaminsky, W.; Woods, T.; Babu, R.P.; Keely, C.M.; Blau, W.; O’connor, K.E. Up-Cycling of PET (Polyethylene Terephthalate) to the Biodegradable Plastic PHA (Polyhydroxyalkanoate). Environ. Sci. Technol. 2008, 42, 7696–7701. [Google Scholar] [CrossRef]
- Urtuvia, V.; Villegas, P.; González, M.; Seeger, M. Bacterial production of the biodegradable plastics polyhydroxyalkanoates. Int. J. Biol. Macromol. 2014, 70, 208–213. [Google Scholar] [CrossRef]
- Spasic, J.; Mandic, M.; Djokic, L.; Nikodinovic-Runic, J. Streptomyces spp. in the biocatalysis toolbox. Appl. Microbiol. Biotechnol. 2018, 102, 3513–3536. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).