
Thermal Inactivation of Hepatitis E Virus in Pork Products Estimated with a Semiquantitative Infectivity Assay
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Stock Preparation
2.2. HEV Extraction from Pork Matrices and Naturally Infected Pig Livers
2.3. Food Processing Conditions
2.4. Cell Culture Method
2.5. Immunofluorescence Detection Assay
2.6. Reverse Transcription Polymerase Chain Reaction
2.7. Bayesian MPN Model
2.7.1. Bayesian MPN Model for Medium, Dried Sausage and Liver Homogenate
2.7.2. Bayesian MPN Model for Naturally Infected Pig Livers
2.7.3. Region of Practical Equivalence (ROPE)
- The HDI was contained within the ROPE: there was no meaningful difference;
- The HDI was completely outside the ROPE: there was a meaningful difference;
- The HDI overlapped the ROPE: we withheld judgment.
3. Results
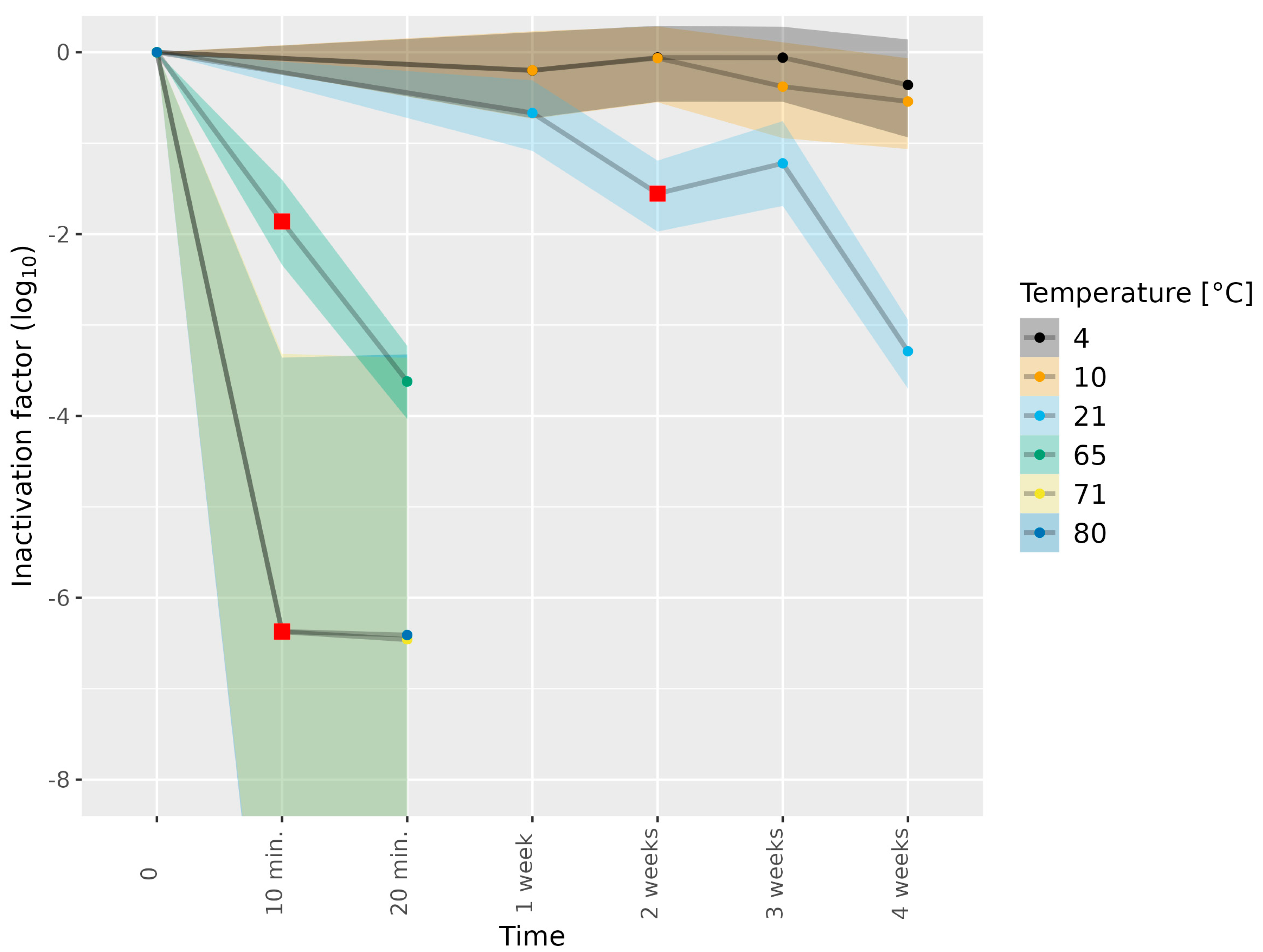
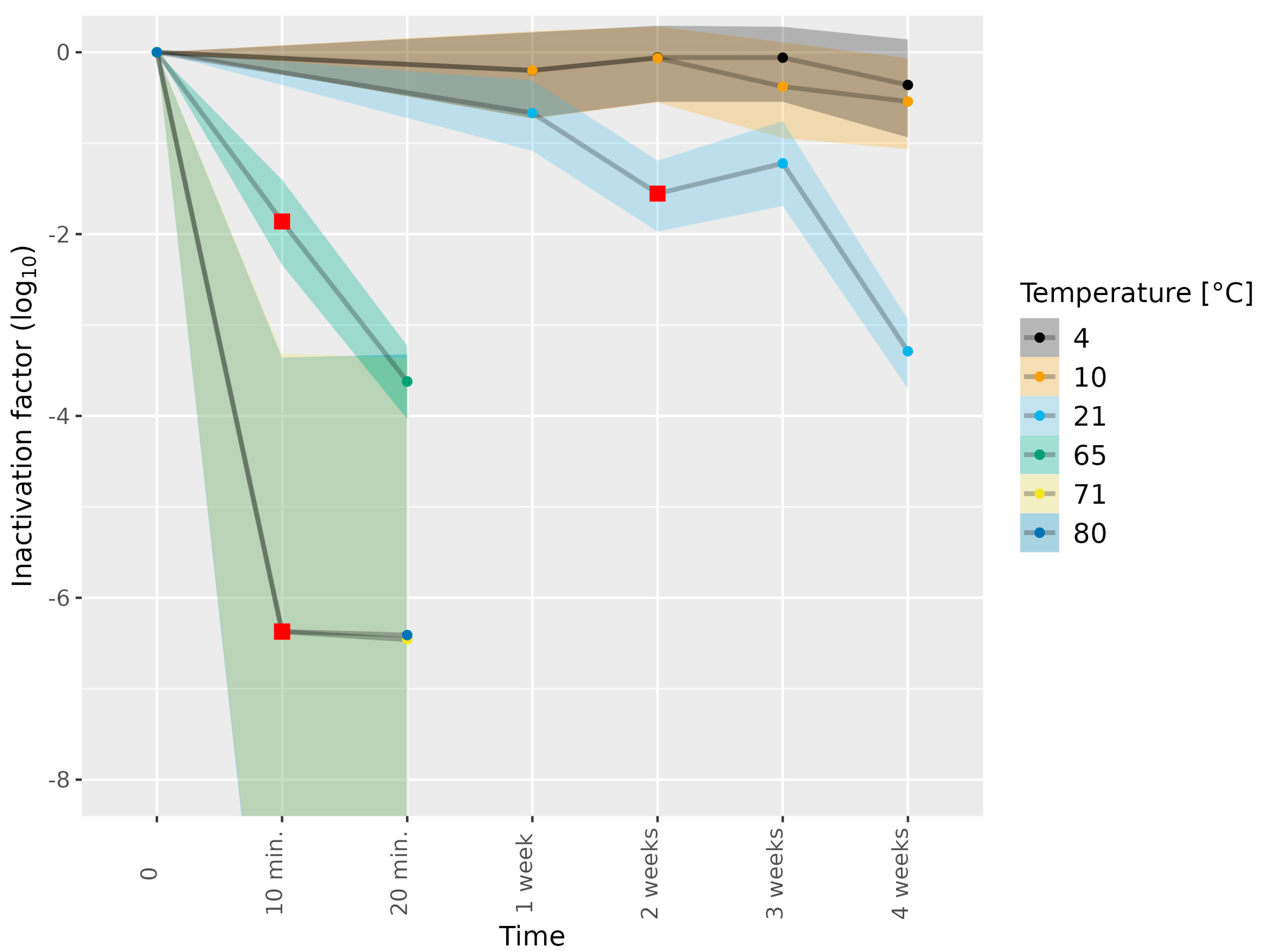
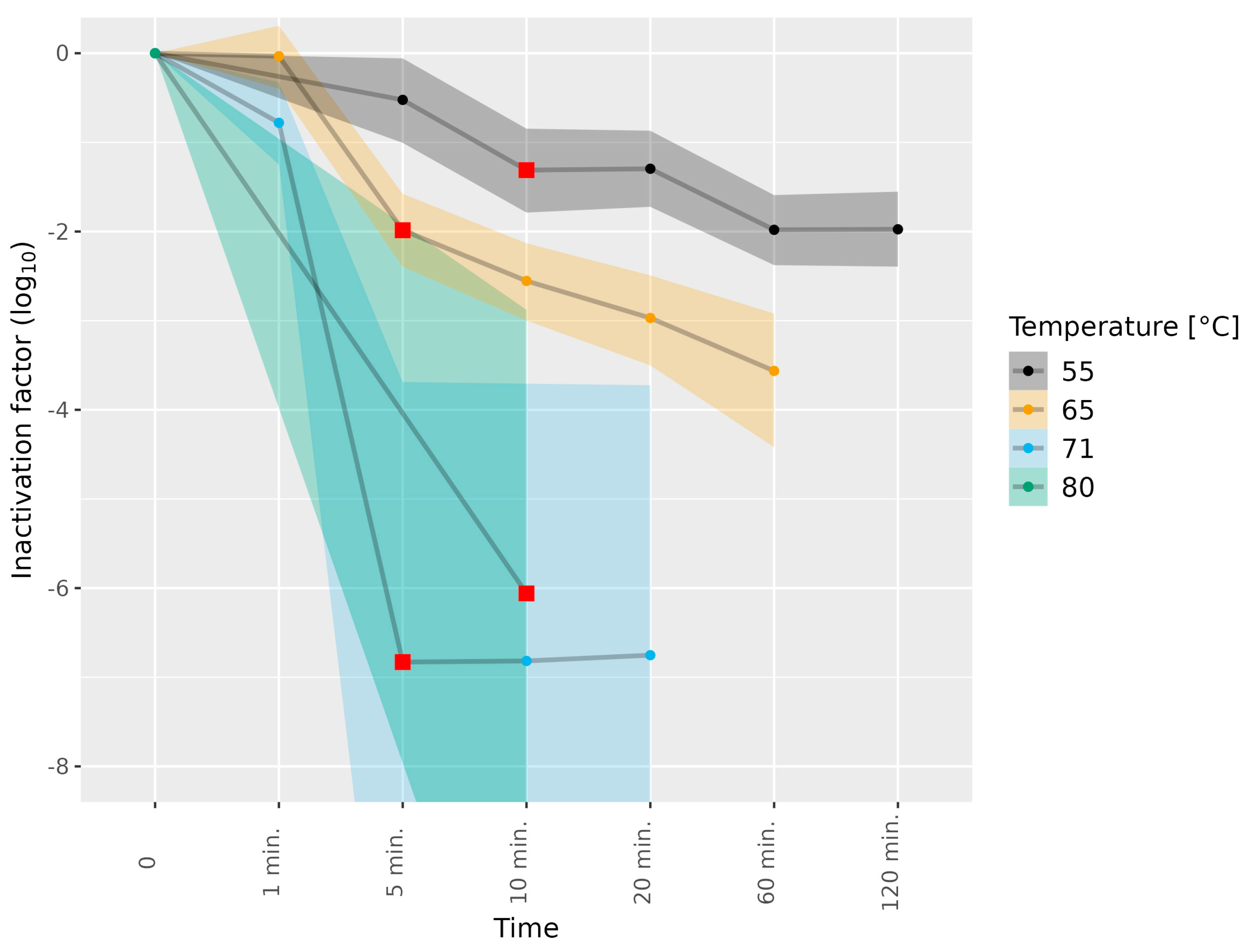
3.1. Determining Infectious HEV in Cell Culture Medium Subjected to Thermal Food-Processing Steps
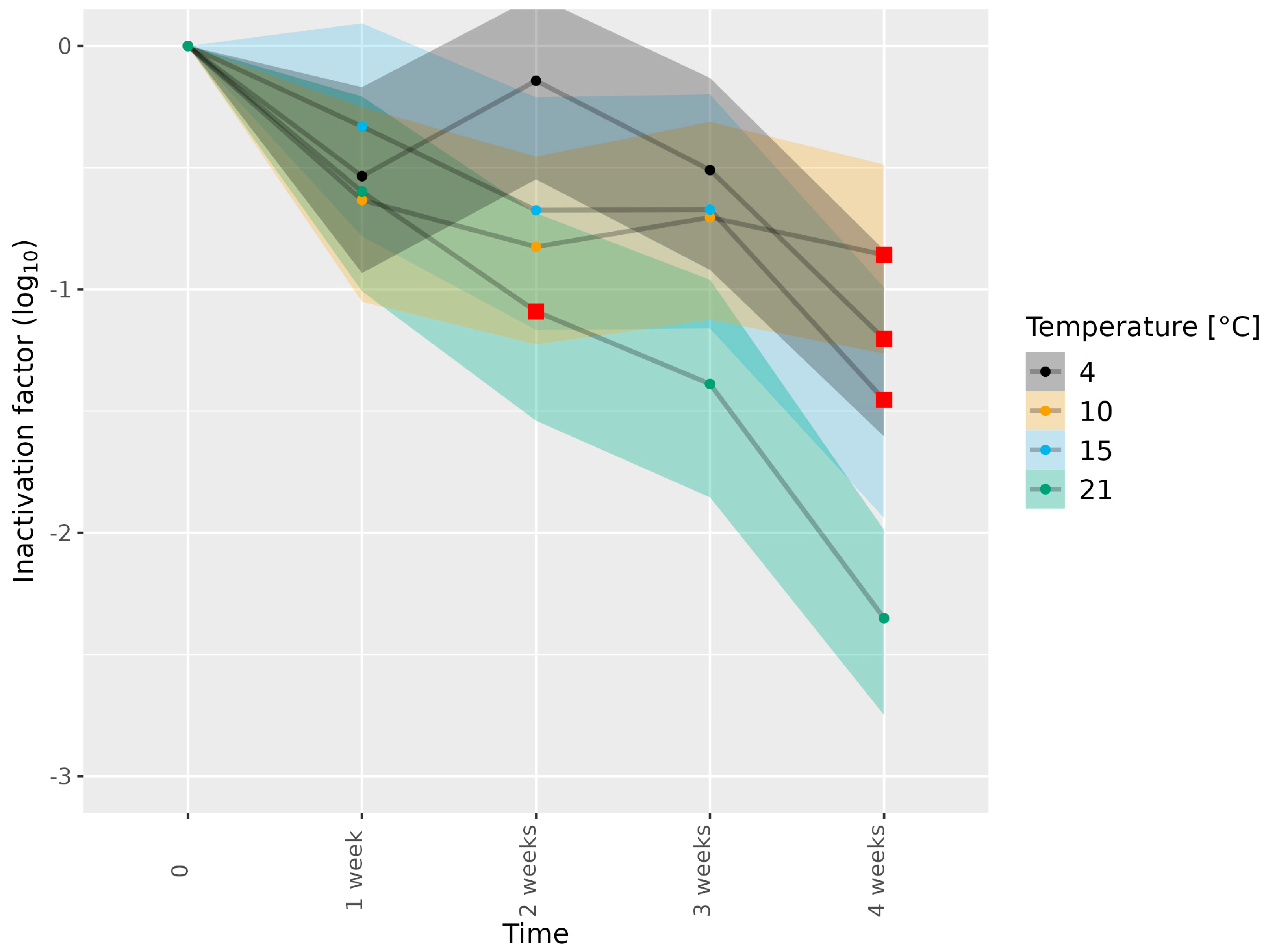
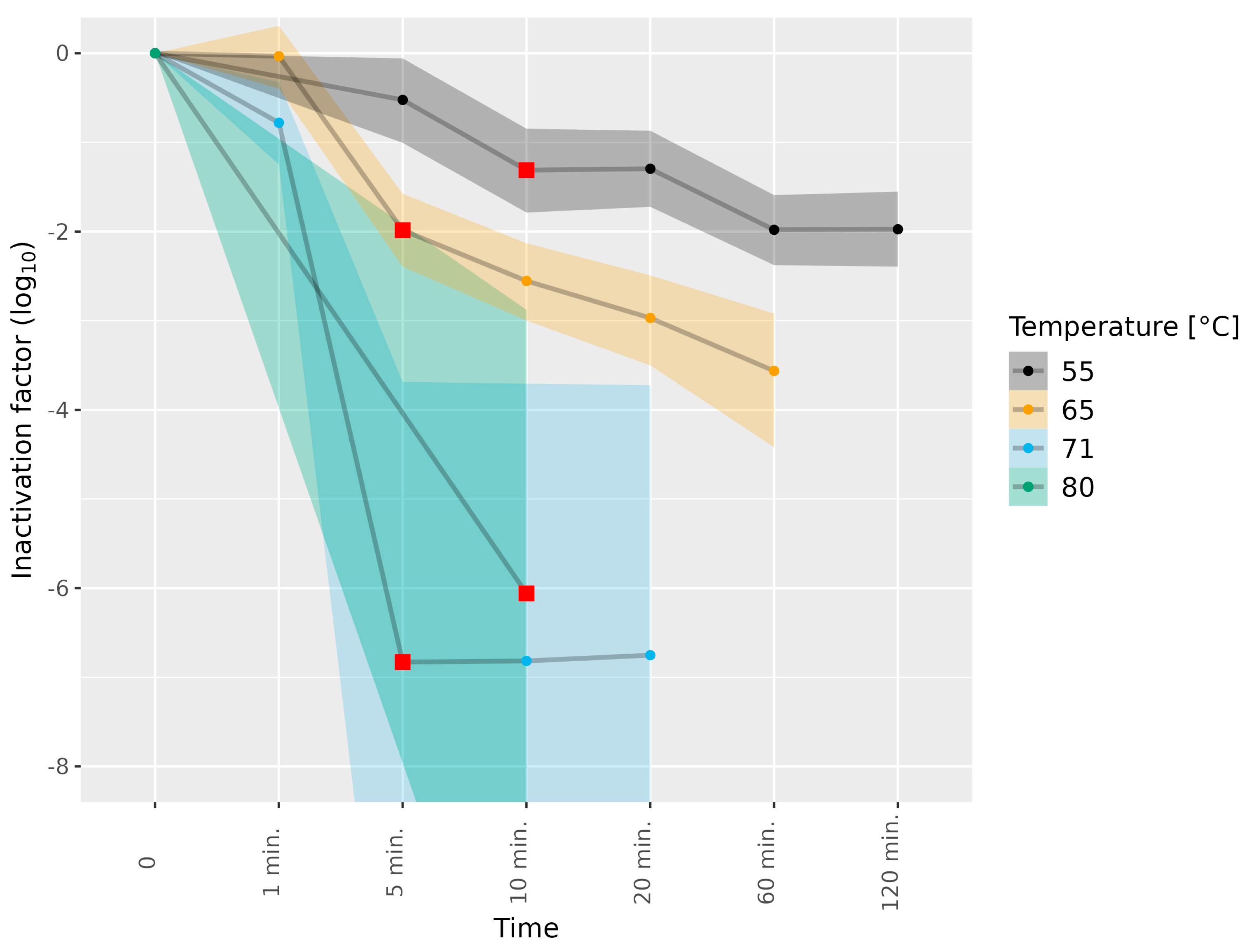
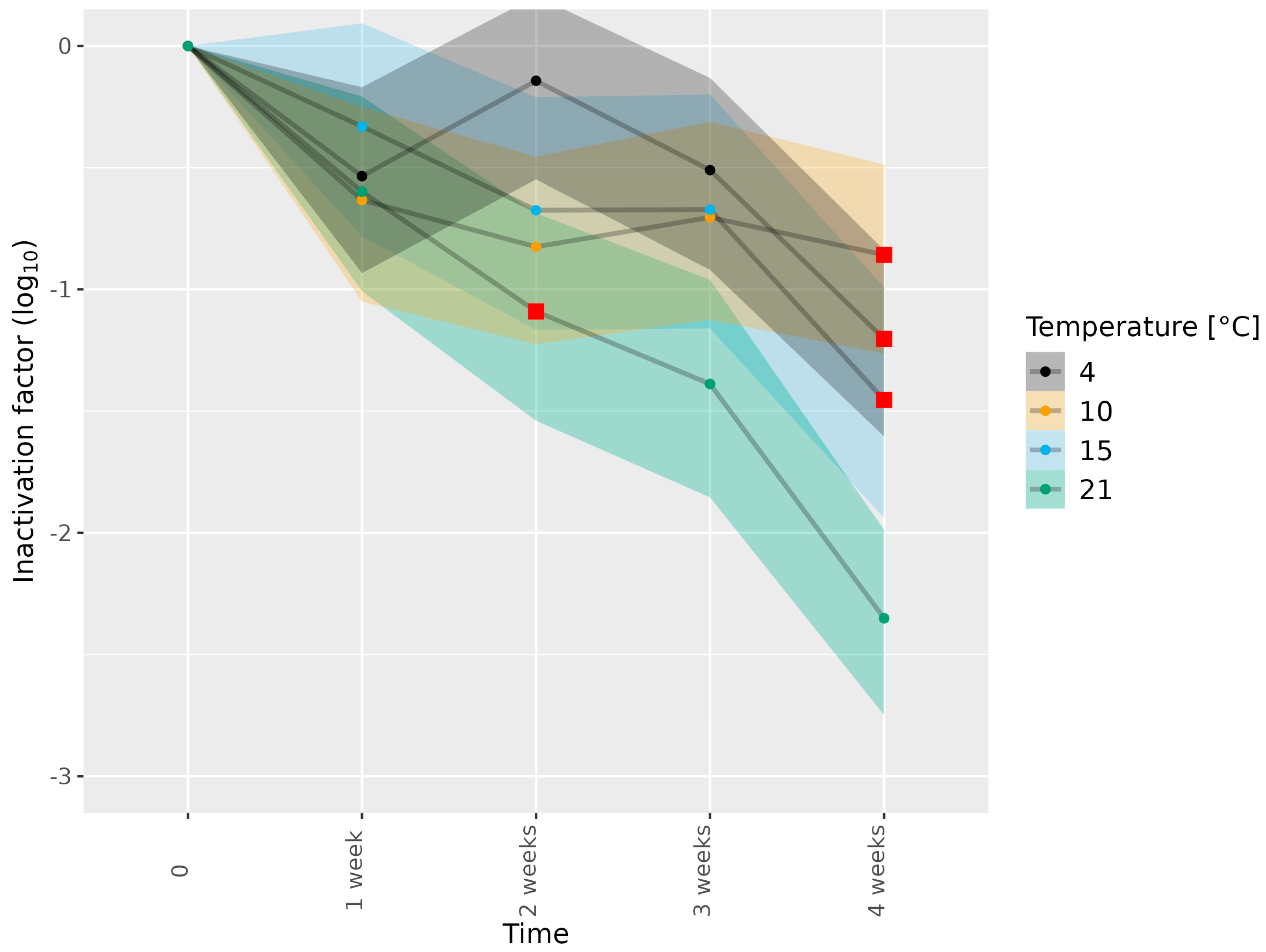
3.2. Determining Infectious HEV in Pork Matrices Subjected to Thermal Food-Processing Steps
3.3. Detection of Infectious HEV in Naturally Infected Pig Livers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hazards, E.P.O.B.; Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernandez Escamez, P.S.; Herman, L.; Koutsoumanis, K.; Lindqvist, R.; et al. Public health risks associated with hepatitis E virus (HEV) as a food-borne pathogen. EFSA J. 2017, 15, e04886. [Google Scholar] [CrossRef]
- Koot, H.; Hogema, B.M.; Koot, M.; Molier, M.; Zaaijer, H.L. Frequent hepatitis E in the Netherlands without traveling or immunosuppression. J. Clin. Virol. 2015, 62, 38–40. [Google Scholar] [CrossRef] [PubMed]
- Adlhoch, C.; Avellon, A.; Baylis, S.A.; Ciccaglione, A.R.; Couturier, E.; de Sousa, R.; Epstein, J.; Ethelberg, S.; Faber, M.; Feher, A.; et al. Hepatitis E virus: Assessment of the epidemiological situation in humans in Europe, 2014/15. J. Clin. Virol. 2016, 82, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Aspinall, E.J.; Couturier, E.; Faber, M.; Said, B.; Ijaz, S.; Tavoschi, L.; Takkinen, J.; Adlhoch, C.; The Country, E. Hepatitis E virus infection in Europe: Surveillance and descriptive epidemiology of confirmed cases, 2005 to 2015. Euro. Surveill. 2017, 22, 30561. [Google Scholar] [CrossRef] [PubMed]
- Hogema, B.M.; Molier, M.; Sjerps, M.; de Waal, M.; van Swieten, P.; van de Laar, T.; Molenaar-de Backer, M.; Zaaijer, H.L. Incidence and duration of hepatitis E virus infection in Dutch blood donors. Transfusion 2016, 56, 722–728. [Google Scholar] [CrossRef]
- Rutjes, S.A.; Bouwknegt, M.; van der Giessen, J.W.; de Roda Husman, A.M.; Reusken, C.B. Seroprevalence of hepatitis E virus in pigs from different farming systems in The Netherlands. J. Food Prot. 2014, 77, 640–642. [Google Scholar] [CrossRef]
- Rutjes, S.A.; Lodder, W.J.; Lodder-Verschoor, F.; van den Berg, H.H.; Vennema, H.; Duizer, E.; Koopmans, M.; de Roda Husman, A.M. Sources of hepatitis E virus genotype 3 in The Netherlands. Emerg. Infect. Dis. 2009, 15, 381–387. [Google Scholar] [CrossRef]
- van der Poel, W.H.M.; Verschoor, F.; van der Heide, R.; Inmaculada Herrera, M.; Vivo, A.; Kooreman, M.; de Roda Husman, A.M. Hepatitis E Virus Sequences in Swine Related to Sequences in Humans, the Netherlands. Emerg. Infect. Dis. 2001, 7, 970–976. [Google Scholar] [CrossRef]
- Ianiro, G.; Pavoni, E.; Aprea, G.; Romantini, R.; Alborali, G.L.; D’Angelantonio, D.; Garofolo, G.; Scattolini, S.; De Sabato, L.; Magistrali, C.F.; et al. Cross-sectional study of hepatitis E virus (HEV) circulation in Italian pig farms. Front Vet. Sci. 2023, 10, 1136225. [Google Scholar] [CrossRef]
- Lapa, D.; Capobianchi, M.R.; Garbuglia, A.R. Epidemiology of Hepatitis E Virus in European Countries. Int. J. Mol. Sci. 2015, 16, 25711–25743. [Google Scholar] [CrossRef]
- Mooij, S.H.; Hogema, B.M.; Tulen, A.D.; van Pelt, W.; Franz, E.; Zaaijer, H.L.; Molier, M.; Hofhuis, A. Risk factors for hepatitis E virus seropositivity in Dutch blood donors. BMC Infect. Dis. 2018, 18, 173. [Google Scholar] [CrossRef] [PubMed]
- van Gageldonk-Lafeber, A.B.; van der Hoek, W.; Borlee, F.; Heederik, D.J.; Mooi, S.H.; Maassen, C.B.; Yzermans, C.J.; Rockx, B.; Smit, L.A.; Reimerink, J.H. Hepatitis E virus seroprevalence among the general population in a livestock-dense area in the Netherlands: A cross-sectional population-based serological survey. BMC Infect. Dis. 2017, 17, 21. [Google Scholar] [CrossRef] [PubMed]
- Tulen, A.D.; Vennema, H.; van Pelt, W.; Franz, E.; Hofhuis, A. A case-control study into risk factors for acute hepatitis E in the Netherlands, 2015–2017. J. Infect. 2019, 78, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Said, B.; Usdin, M.; Warburton, F.; Ijaz, S.; Tedder, R.S.; Morgan, D. Pork products associated with human infection caused by an emerging phylotype of hepatitis E virus in England and Wales. Epidemiol. Infect. 2017, 145, 2417–2423. [Google Scholar] [CrossRef] [PubMed]
- Boxman, I.L.A.; Jansen, C.C.C.; Hagele, G.; Zwartkruis-Nahuis, A.; Tijsma, A.S.L.; Vennema, H. Monitoring of pork liver and meat products on the Dutch market for the presence of HEV RNA. Int. J. Food Microbiol. 2019, 296, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Boxman, I.L.A.; Jansen, C.C.C.; Zwartkruis-Nahuis, A.J.T.; Hagele, G.; Sosef, N.P.; Dirks, R.A.M. Detection and quantification of hepatitis E virus RNA in ready to eat raw pork sausages in the Netherlands. Int. J. Food Microbiol. 2020, 333, 108791. [Google Scholar] [CrossRef]
- Boxman, I.L.A.; Verhoef, L.; Dop, P.Y.; Vennema, H.; Dirks, R.A.M.; Opsteegh, M. High prevalence of acute hepatitis E virus infection in pigs in Dutch slaughterhouses. Int. J. Food Microbiol. 2022, 379, 109830. [Google Scholar] [CrossRef]
- Bouwknegt, M.; Lodder-Verschoor, F.; van der Poel, W.; Rutjes, S.; de Roda Husman, A.M. Hepatitis E Virus RNA in Commercial Porcine Livers in The Netherlands. J. Food Prot. 2007, 70, 2889–2895. [Google Scholar] [CrossRef]
- Colson, P.; Borentain, P.; Queyriaux, B.; Kaba, M.; Moal, V.; Gallian, P.; Heyries, L.; Raoult, D.; Gerolami, R. Pig liver sausage as a source of hepatitis E virus transmission to humans. J. Infect. Dis. 2010, 202, 825–834. [Google Scholar] [CrossRef]
- Evers, E.G.; Pielaat, A.; Smid, J.H.; van Duijkeren, E.; Vennemann, F.B.; Wijnands, L.M.; Chardon, J.E. Comparative Exposure Assessment of ESBL-Producing Escherichia coli through Meat Consumption. PLoS ONE 2017, 12, e0169589. [Google Scholar] [CrossRef]
- Ferigolo, L.P.; de Oliveira Elias, S.; Carmo da Silva, D.; Lopes, S.M.; Geimba, M.P.; Tondo, E.C. Escherichia coli inactivation on tenderloin beef medallions processed by sous vide treatment. Int. J. Gastron. Food Sci. 2021, 25, 100366. [Google Scholar] [CrossRef]
- Baldwin, D.E. Sous vide cooking: A review. Int. J. Gastron. Food Sci. 2012, 1, 15–30. [Google Scholar] [CrossRef]
- Feagins, A.R.; Opriessnig, T.; Guenette, D.K.; Halbur, P.G.; Meng, X.J. Inactivation of infectious hepatitis E virus present in commercial pig livers sold in local grocery stores in the United States. Int. J. Food Microbiol. 2008, 123, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Imagawa, T.; Sugiyama, R.; Shiota, T.; Li, T.C.; Yoshizaki, S.; Wakita, T.; Ishii, K. Evaluation of Heating Conditions for Inactivation of Hepatitis E Virus Genotypes 3 and 4. J. Food Prot. 2018, 81, 947–952. [Google Scholar] [CrossRef] [PubMed]
- Barnaud, E.; Rogee, S.; Garry, P.; Rose, N.; Pavio, N. Thermal inactivation of infectious hepatitis E virus in experimentally contaminated food. Appl. Environ. Microbiol. 2012, 78, 5153–5159. [Google Scholar] [CrossRef]
- Johne, R.; Trojnar, E.; Filter, M.; Hofmann, J. Thermal Stability of Hepatitis E Virus as Estimated by a Cell Culture Method. Appl. Environ. Microbiol. 2016, 82, 4225–4231. [Google Scholar] [CrossRef]
- Schielke, A.; Filter, M.; Appel, B.; Johne, R. Thermal stability of hepatitis E virus assessed by a molecular biological approach. Virol. J. 2011, 8, 487. [Google Scholar] [CrossRef]
- Schemmerer, M.; Johne, R.; Erl, M.; Jilg, W.; Wenzel, J.J. Isolation of Subtype 3c, 3e and 3f-Like Hepatitis E Virus Strains Stably Replicating to High Viral Loads in an Optimized Cell Culture System. Viruses 2019, 11, 483. [Google Scholar] [CrossRef]
- Schemmerer, M.; Apelt, S.; Trojnar, E.; Ulrich, R.G.; Wenzel, J.J.; Johne, R. Enhanced Replication of Hepatitis E Virus Strain 47832c in an A549-Derived Subclonal Cell Line. Viruses 2016, 8, 267. [Google Scholar] [CrossRef]
- Kubickova, B.; Schenk, J.A.; Ramm, F.; Markuškienė, K.; Reetz, J.; Dremsek, P.; Tamosiunas, P.L.; Cepulyte, L.; Trinh, H.A.; Scholz, J.; et al. A broadly cross-reactive monoclonal antibody against hepatitis E virus capsid antigen. Appl. Microbiol Biotechnol. 2021, 105, 4957–4973. [Google Scholar] [CrossRef]
- Scheltinga, S.A.; Templeton, K.E.; Beersma, M.F.; Claas, E.C. Diagnosis of human metapneumovirus and rhinovirus in patients with respiratory tract infections by an internally controlled multiplex real-time RNA PCR. J. Clin. Virol. 2005, 33, 306–311. [Google Scholar] [CrossRef]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef]
- Boxman, I.L.A.; Jansen, C.C.C.; Hagele, G.; Zwartkruis-Nahuis, A.; Cremer, J.; Vennema, H.; Tijsma, A.S.L. Porcine blood used as ingredient in meat productions may serve as a vehicle for hepatitis E virus transmission. Int. J. Food Microbiol. 2017, 257, 225–231. [Google Scholar] [CrossRef]
- Stan Development Team. Stan Modeling Language Users Guide and Reference Manual, Version 2.30. Stan-Dev.Github.Io. Stan Development Team: St. Louis, MO, USA, 2022. Available online: https://mc-stan.org/(accessed on 10 July 2023).
- R Core Team. R Foundation for Statistical Computing, Vienna, Austria. 2023. Available online: https://www.R-project.org (accessed on 10 July 2023).
- Gilks, W.R.; Richardson, S.; Spiegelhalter, D.J. Markov Chain Monte Carlo in Practice; Chapman & Hall: London, UK, 1995. [Google Scholar]
- Kruschke, J.K. Rejecting or Accepting Parameter Values in Bayesian Estimation. Adv. Methods Pract. Psychol. Sci. 2018, 1, 270–280. [Google Scholar] [CrossRef]
- Okamoto, H. Culture systems for hepatitis E virus. J. Gastroenterol. 2013, 48, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Li, X.; Feng, Z. Role of Envelopment in the HEV Life Cycle. Viruses 2016, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Mc, L.M.; Belford, G.; Harlow, J.; Nasheri, N. Examining the Effect of Organic Acids on Inactivation of Hepatitis E Virus. J. Food Prot. 2022, 85, 1690–1695. [Google Scholar] [CrossRef]
- Bouquet, P.; Alexandre, V.; De Lamballerie, M.; Ley, D.; Lesage, J.; Goffard, A.; Cocquerel, L. Effect of High Hydrostatic Pressure Processing and Holder Pasteurization of Human Milk on Inactivation of Human Coronavirus 229E and Hepatitis E Virus. Viruses 2023, 15, 1571. [Google Scholar] [CrossRef] [PubMed]
- Lopez Borela, V.; de Alencar, E.R.; Mendonca, M.A.; Han, H.; Raposo, A.; Ariza-Montes, A.; Araya-Castillo, L.; Zandonadi, R.P. Influence of Different Cooking Methods on Fillet Steak Physicochemical Characteristics. Int. J. Environ. Res. Public Health 2022, 19, 606. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Matrix | HEV-3 Subtype | Temperature (°C) | Duration | Number of Biological Replicates |
|---|---|---|---|---|
| No matrix (medium) | 3c | 21 | 1, 4 weeks | 1 |
| No matrix (medium) | 3e | 4, 10 | 1, 2, 3, 4 weeks | 1 |
| No matrix (medium) | 3e | 21 | 2, 3 weeks | 1 |
| No matrix (medium) | 3e | 21 | 1, 4 weeks | 2 |
| No matrix (medium) | 3c | 65 | 10, 20 min | 1 |
| No matrix (medium) | 3e | 65 | 10, 20 min | 1 |
| No matrix (medium) | 3e | 71 | 10, 20 min | 1 |
| No matrix (medium) | 3e | 80 | 10, 20 min | 1 |
| Dried sausage | 3c | 4, 10, 15, 21 | 1, 2, 3, 4 weeks | 1 |
| Dried sausage | 3e | 4, 10, 21 | 1, 2, 3, 4 weeks | 2 |
| Dried sausage | 3e | 15 | 1, 2, 3, 4 weeks | 1 |
| Liver homogenate | 3c | 55 | 5, 10, 20, 60, 120 min | 1 |
| Liver homogenate | 3e | 55 | 5, 10, 20, 60, 120 min | 1 |
| Liver homogenate | 3c | 65 | 1, 5, 10, 20, 60 min | 1 |
| Liver homogenate | 3e | 65 | 1, 5, 10, 20, 60 min | 1 |
| Liver homogenate | 3c | 71 | 1, 5, 10, 20 min | 1 |
| Liver homogenate | 3e | 71 | 1, 5 min | 1 |
| Liver homogenate | 3e | 71 | 10, 20 min | 2 |
| Liver homogenate | 3e | 80 | 10 min | 1 |
| Pig Liver ID | Dilution with Positive IF Results | Log10 Viral Particles/g (95% C.I.) | Cq Values (RIVM) | Cq Values (WFSR) |
|---|---|---|---|---|
| L-01 | 5× (8)/10× (3)/100× (1) | 2.7 [2.42, 2.96] | 31.38 | 33.75 |
| L-02 | n.d. | <0.66 | n.d. | 35.62 |
| L-03 | n.d. | <0.66 | 31.24 | 32.17 |
| L-04 | n.d. | <0.66 | 31.09 | 26.53 |
| L-05 | 100× (8) | >2.96 | 25.68 | 26.53 |
| L-06 | 100× (8) | >2.96 | 24.58 | 24.17 |
| L-07 | 100× (8) | >2.96 | 25.52 | 27.24 |
| L-08 | n.d. | <0.66 | 35.71 | 35.72 |
| L-09 | n.d. | <0.66 | 32.88 | 33.25 |
| L-10 | n.d. | <0.66 | 27.17 | 26.42 |
| L-11 | 5× (0)/10× (2)/100× (0) | 1.5 [0.66, 2.09] | 31.18 | 29.55 |
| L-12 | 5× (8)/10× (8)/100× (7) | >2.96 | 27.44 | 22.15 |
| L-13 | n.d. | <0.66 | n.d. | n.d. |
| L-14 | n.d. | <0.66 | 34.66 | 34.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stunnenberg, M.; Huizen, S.C.v.; Swart, A.; Lodder, W.J.; Boxman, I.L.A.; Rutjes, S.A. Thermal Inactivation of Hepatitis E Virus in Pork Products Estimated with a Semiquantitative Infectivity Assay. Microorganisms 2023, 11, 2451. https://doi.org/10.3390/microorganisms11102451
Stunnenberg M, Huizen SCv, Swart A, Lodder WJ, Boxman ILA, Rutjes SA. Thermal Inactivation of Hepatitis E Virus in Pork Products Estimated with a Semiquantitative Infectivity Assay. Microorganisms. 2023; 11(10):2451. https://doi.org/10.3390/microorganisms11102451
Chicago/Turabian StyleStunnenberg, Melissa, Suzanne C. van Huizen, Arno Swart, Willemijn J. Lodder, Ingeborg L. A. Boxman, and Saskia A. Rutjes. 2023. "Thermal Inactivation of Hepatitis E Virus in Pork Products Estimated with a Semiquantitative Infectivity Assay" Microorganisms 11, no. 10: 2451. https://doi.org/10.3390/microorganisms11102451
APA StyleStunnenberg, M., Huizen, S. C. v., Swart, A., Lodder, W. J., Boxman, I. L. A., & Rutjes, S. A. (2023). Thermal Inactivation of Hepatitis E Virus in Pork Products Estimated with a Semiquantitative Infectivity Assay. Microorganisms, 11(10), 2451. https://doi.org/10.3390/microorganisms11102451