
Optimization of the Fermentation Conditions of Metarhizium robertsii and Its Biological Control of Wolfberry Root Rot Disease


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strain, Culture Medium, and Wolfberry Plant
2.2. Determination of the Antagonistic Effect of HYC-7 Strain on F. solani
2.3. Screening of Optimum Carbon and Nitrogen Sources for Fermentation Medium
2.4. Single-Factor Test of the Effect of Different Fermentation Conditions on the Inhibition Rate of HYC-7 Fermentation Broth
2.5. Response Surface Optimization Test
2.6. Determination of Colony Diameter, Sporulation, Spore Germination Rate, and Germ Tube Length
2.7. Determination of Antioxidant Enzymes Activity
2.8. Determination of Content of Malondialdehyde, Soluble Protein, and Soluble Sugar
2.9. Determination of Decay Incidence and Disease Index of Wolfberry
2.10. Data Analysis
3. Results
3.1. HYC-7 Strain on F. solani
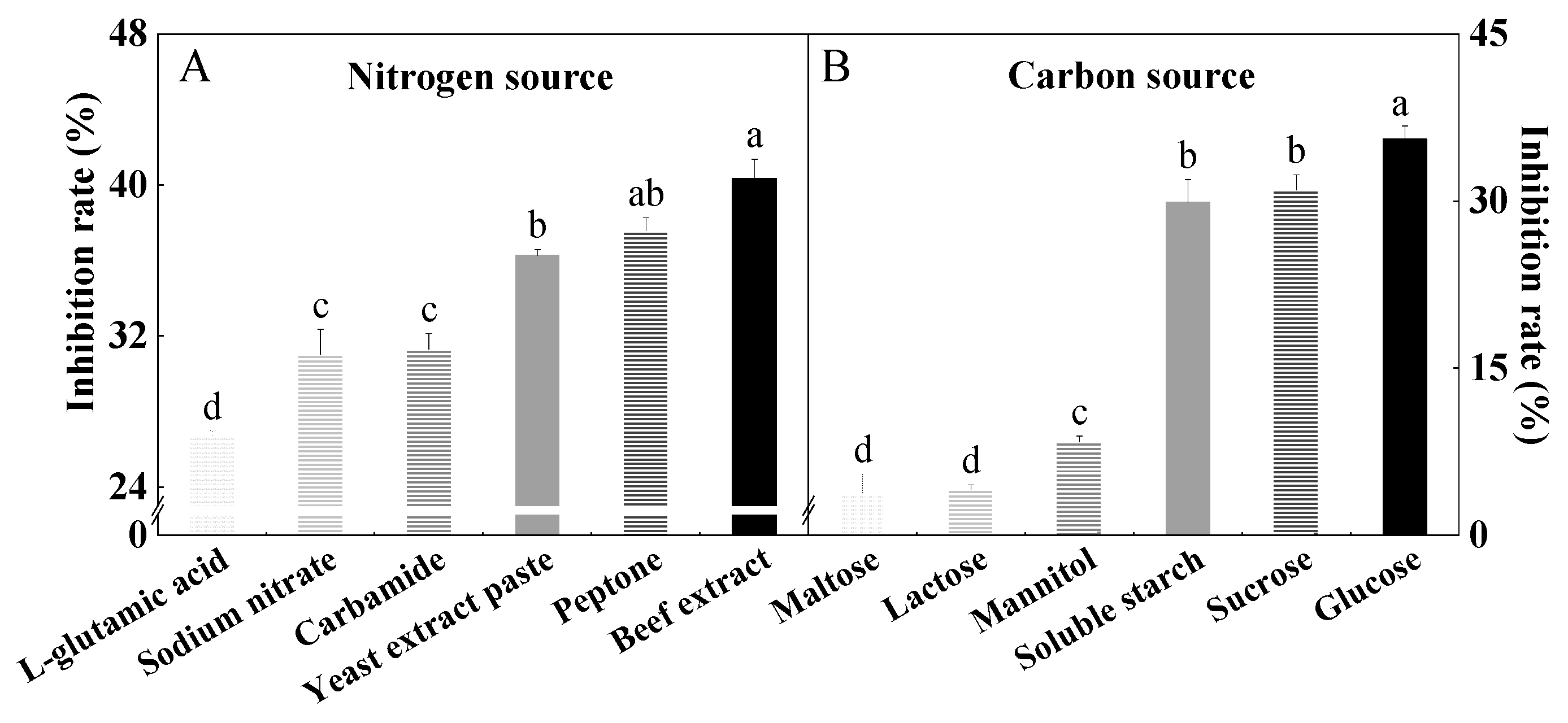
3.2. Screening of Optimum Nitrogen and Carbon Sources for Fermentation Medium
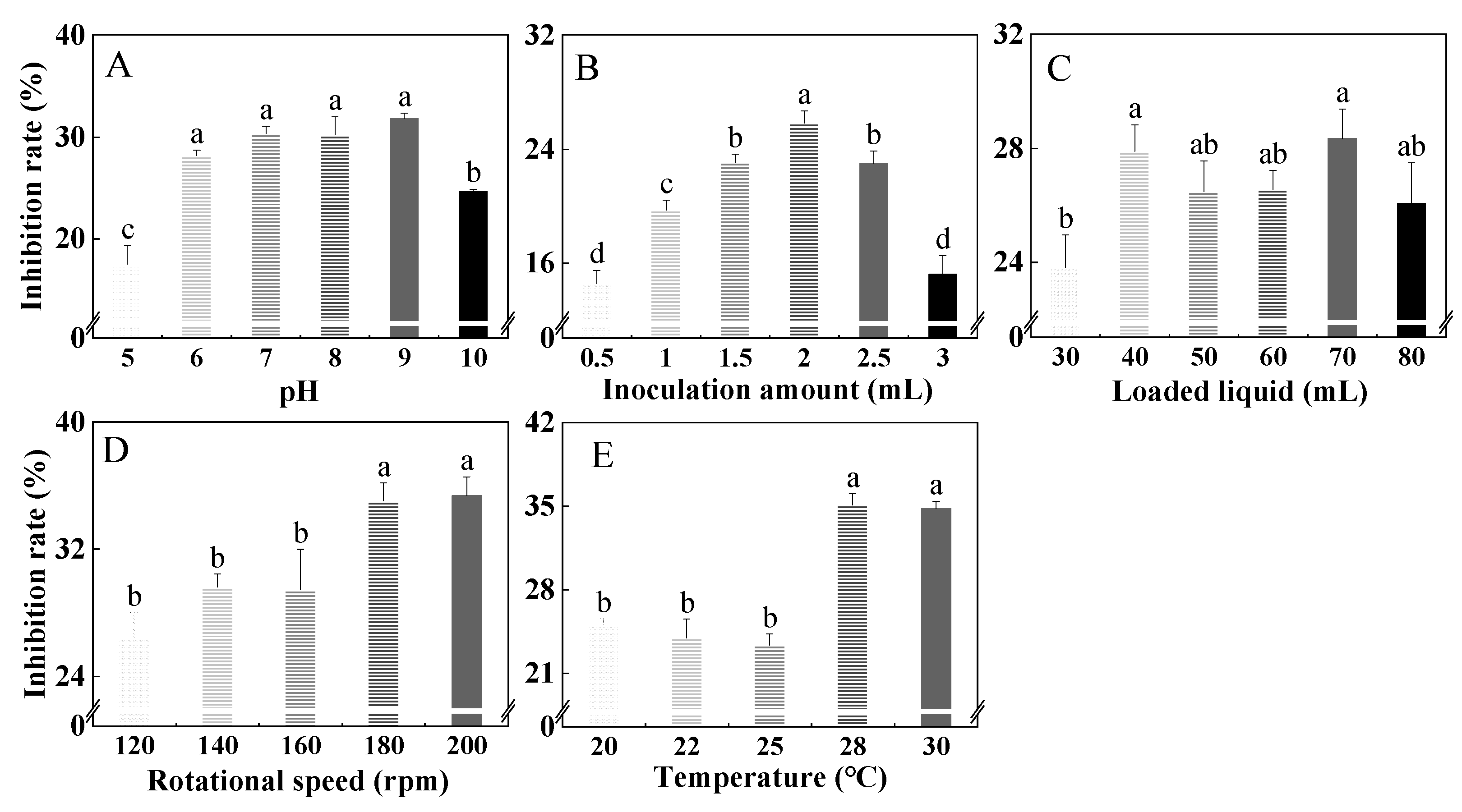
3.3. Single-Factor Test
3.4. Response Surface Test Optimization Results
3.4.1. Regression Equation Fitting and Analysis of Variance
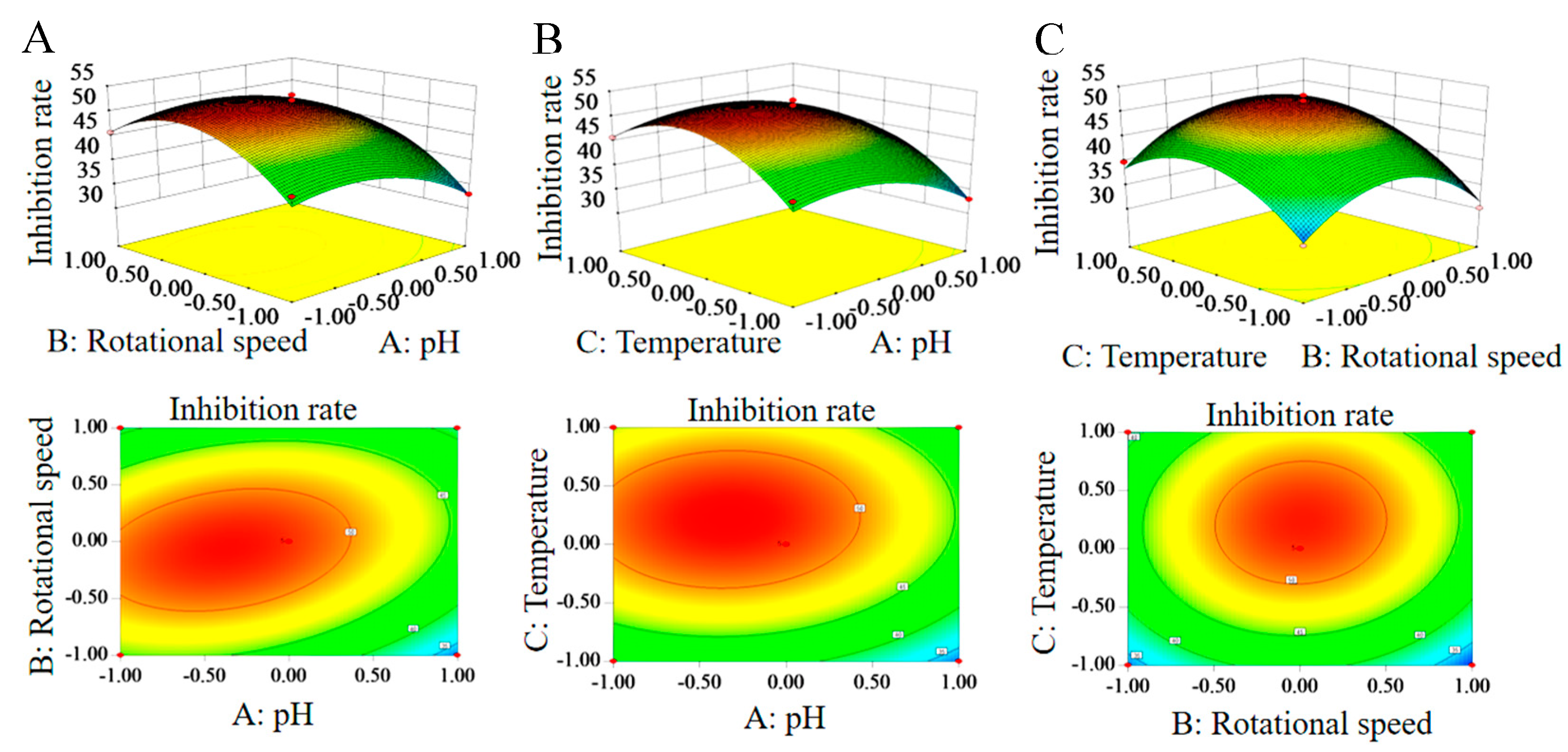
3.4.2. Response Surface Analysis of Interaction of Various Factors
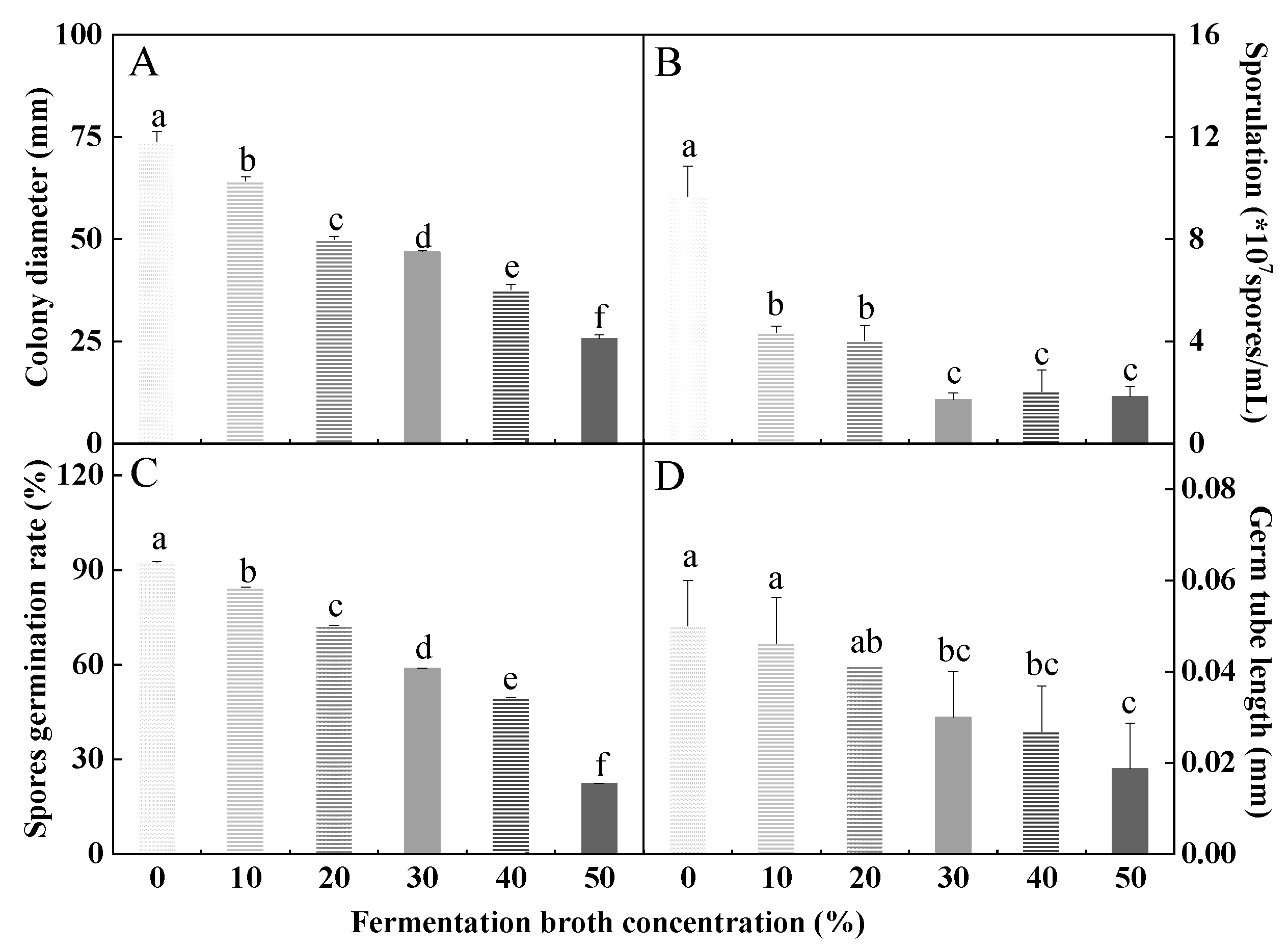
3.5. Effects of HYC-7 Fermentation Broth on the Growth and Development of F. solani
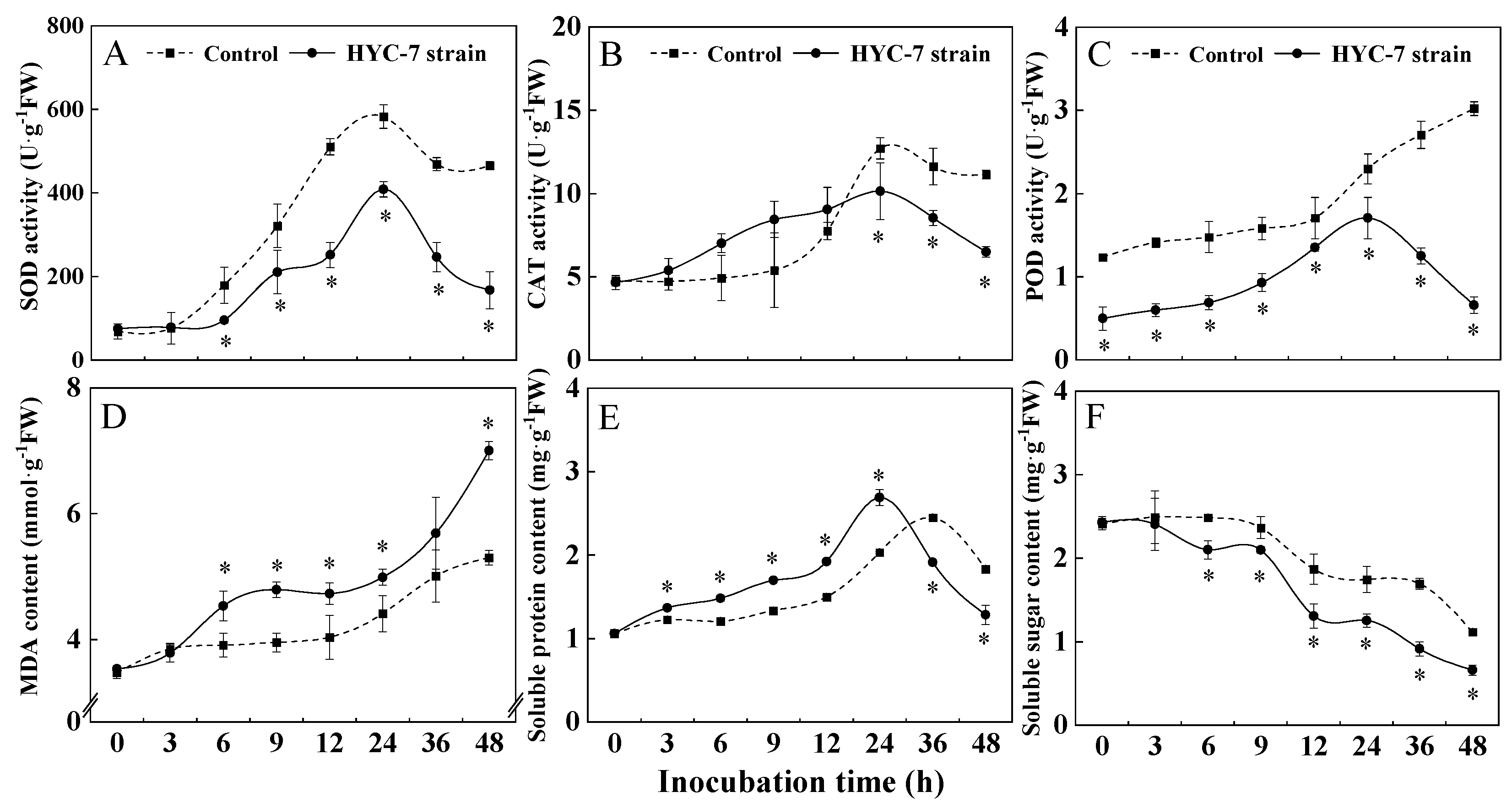
3.6. Effects of HYC-7 Strain Fermentation Broth on the Activities of SOD, CAT, and POD and the Contents of MDA, Soluble Protein, and Soluble Sugar
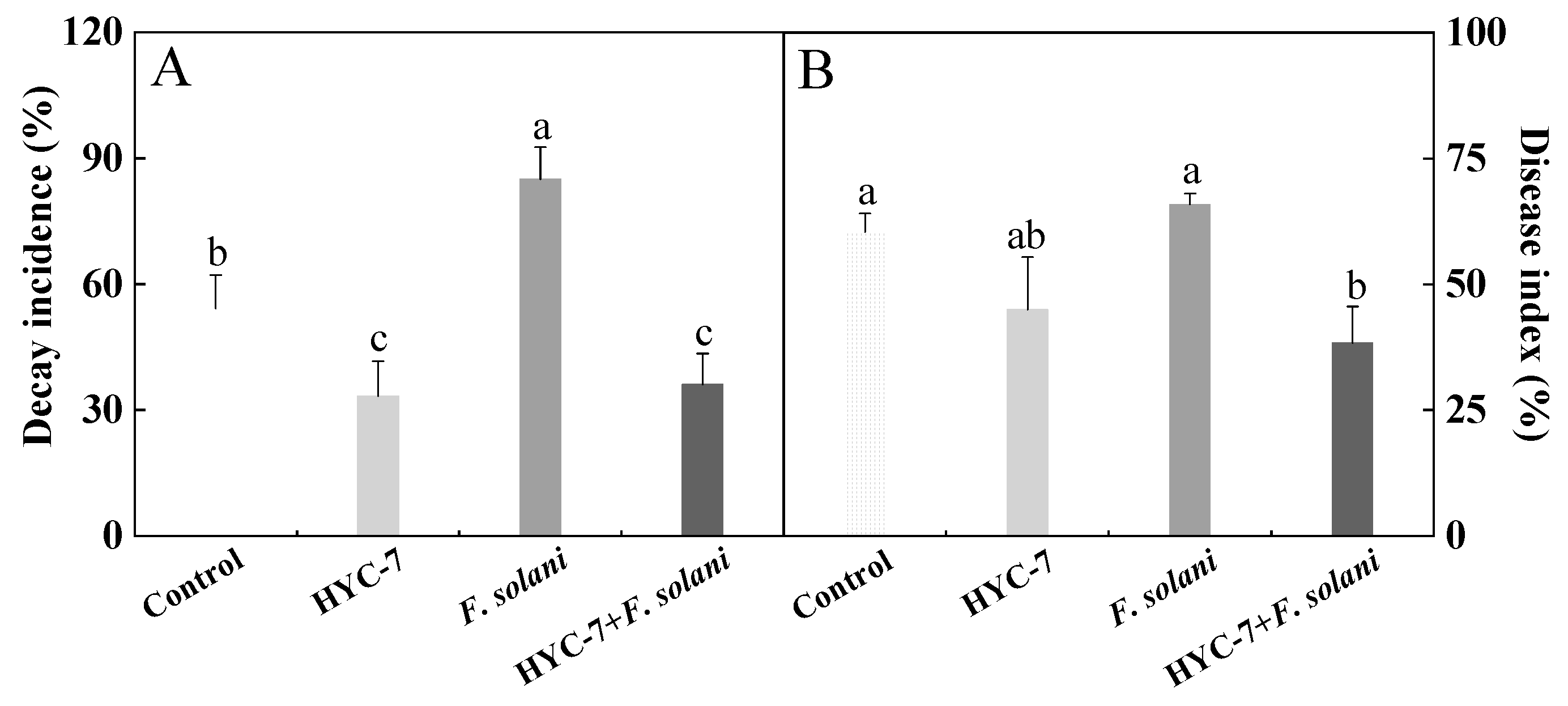
3.7. Effect of HYC-7 Fermentation Broth on the Decay Incidence and Disease Index of Wolfberry Root Rot
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, D.; Xia, T.; Dang, S.; Fan, G.; Wang, Z. Investigation of Chinese wolfberry (Lycium spp.) germplasm by restriction site-associated DNA sequencing (RAD-seq). Biochem. Genet. 2018, 56, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wei, Y.; Wang, Y.; Gao, F.; Chen, Z. Lycium barbarum, a traditional Chinese herb and promising anti-aging agent. Aging Dis. 2017, 8, 778–791. [Google Scholar] [CrossRef] [PubMed]
- Uwaremwe, C.; Yue, L.; Liu, Y.; Tian, Y.; Zhao, X.; Wang, Y.; Xie, Z.K.; Zhang, Y.B.; Cui, Z.T.; Wang, R.Y. Molecular identification and pathogenicity of Fusarium and Alternaria species associated with root rot disease of wolfberry in Gansu and Ningxia provinces, China. Plant Pathol. 2021, 70, 397–406. [Google Scholar] [CrossRef]
- Rojo, F.G.; Reynoso, M.M.; Ferez, M.; Sofía, N.; Chulze, S.N.; Torres, A.M. Biological control by Trichoderma species of Fusarium solani causing peanut brown root rot under field conditions. Crop Prot. 2007, 26, 549–555. [Google Scholar] [CrossRef]
- Al-Ani, R.A.; Adhab, M.A.; Mahdi, M.H.; Abood, H.M. Rhizobium japonicum as a biocontrol agent of soybean root rot disease caused by Fusarium solani and Macrophomina phaseolina. Plant Protect Sci. 2012, 48, 149–155. [Google Scholar] [CrossRef]
- Toghueo, R.; Eke, P.; Zabalgogeazcoab, Í.Z.; Aldana, B.D.; Nana, L.W.; Boyom, F.F. Biocontrol and growth enhancement potential of two endophytic Trichoderma spp. from Terminalia catappa against the causative agent of common bean root rot (Fusarium solani). Biol. Control 2016, 96, 8–20. [Google Scholar] [CrossRef]
- Sasan, R.K.; Bidochka, M.J. Antagonism of the endophytic insect pathogenic fungus Metarhizium robertsii against the bean plant pathogen Fusarium solani f. sp. phaseoli. Can. J. Plant Pathol. 2013, 35, 288–293. [Google Scholar] [CrossRef]
- Zhang, X.Y.; He, J.; Hou, C.X.; Zhang, S.H. Screening and identification of antagonistic strains of wolfberry root rot. Acta Agric. Zhejiangensis 2020, 32, 858–865. [Google Scholar]
- Sasan, R.K.; Bidochka, M.J. The insect-pathogenic fungus Metarhizium robertsii (Clavicipitaceae) is also an endophyte that stimulates plant root development. Am. J. Bot. 2012, 99, 101–107. [Google Scholar] [CrossRef]
- Velavan, V.; Rangeshwaran, R.; Sivakumar, G.; Sasidharan, T.O.; Sundararaj, R.; Kandan, A. Occurrence of Metarhizium spp. isolated from forest samples in south india and their potential in biological control of banana stem weevil Odoiporus longicollis oliver. Egypt J. Biol. Pest Control 2021, 31, 131. [Google Scholar] [CrossRef]
- Plantey, R.L.; Papura, D.; Couture, C.; Thiéry, D.; Lucero, G.S. Characterization of entomopathogenic fungi from vineyards in argentina with potential as biological control agents against the European grapevine moth Lobesia botrana. BioControl 2019, 64, 501–511. [Google Scholar] [CrossRef]
- Heller, J.; Tudzynski, P. Reactive oxygen species in phytopathogenic fungi: Signaling, development, and disease. Annu. Rev. Phytopathol. 2011, 49, 369–390. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Feng, J.T.; Zhang, Q.; Zhang, X. Optimization of fermentation condition for antibiotic production by Xenorhabdus nematophila with response surface methodology. J. Appl. Microbiol. 2008, 104, 735–744. [Google Scholar] [CrossRef]
- Wanicki, S.A.; Mascarin, G.M.; Moreno, S.G.; Eilenberg, J.; Júnior, I.D. Growth kinetic and nitrogen source optimization for liquid culture fermentation of Metarhizium robertsii blastospores and bioefficacy against the corn leafhopper Dalbulus maidis. World J. Microbiol. Biot. 2020, 36, 71. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; He, Q.; Yun, J.; Huang, L. Optimization of nutrient and fermentation parameters for antifungal activity by Streptomyces lavendulae xjy and its biocontrol efficacies against Fulvia fulva and Botryosphaeria dothidea. J. Phytopathol. 2016, 164, 155–165. [Google Scholar] [CrossRef]
- Li, M.L.; Zheng, T.T.; Chen, Y.Q.; Sui, Y.; Ding, R.H.; Hou, L.W.; Zheng, F.L.; Zhu, C.Y. The antagonistic mechanisms of Streptomyces sioyaensis on the growth and metabolism of poplar canker pathogen Valsa sordida. Biol. Control 2020, 151, 104392. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, Y.D.; Li, Y.C.; Bi, Y.; Mao, R.Y.; Yang, Y.Y.; Jiang, Q.Q.; Prusky, D. Cellular responses required for oxidative stress tolerance of the necrotrophic fungus Alternaria alternata, causal agent of pear black spot. Microorganisms 2022, 10, 621. [Google Scholar] [CrossRef]
- Wang, B.; Li, Z.C.; Han, Z.H.; Xue, S.L.; Bi, Y.; Prusky, D. Effects of nitric oxide treatment on lignin biosynthesis and texture properties at wound sites of muskmelons. Food Chem. 2021, 362, 130193. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dai, T.T.; Xu, Z.Y.; Zhou, X.; Li, B.; Mao, S.F. The inhibitory effect of the plant alkaloid camptothecin on the rice sheath blight pathogen Rhizoctonia solani. Int. J. Agric. Biol. 2017, 19, 558–562. [Google Scholar] [CrossRef]
- Dai, Y.; Wang, Y.H.; Li, M.; Zhu, M.L.; Wen, T.Y.; Wu, X.Q. Medium optimization to analyze the protein composition of Bacillus pumilus HR10 antagonizing Sphaeropsis sapinea. AMB Express 2022, 12, 61. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.L.; Chen, J.W.; Tzeng, Y.M. Production of cyclodepsipeptides destruxin A and B from Metarhizium anisopliae. Biotechnol. Progr. 2000, 16, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Han, Y.; Tan, X.Q.; Wang, J.; Zhou, Z.J. Optimization of antifungal lipopeptide production from Bacillus sp. BH072 by response surface methodology. J. Microbiol. 2014, 52, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Sa, R.B.; He, S.; Han, D.D.; Liu, M.J.; Yu, Y.X.; Shang, R.G.; Song, M.M. Isolation and identification of a new biocontrol bacteria against Salvia miltiorrhiza root rot and optimization of culture conditions for antifungal substance production using response surface methodology. BMC Microbiol. 2022, 22, 231. [Google Scholar] [CrossRef] [PubMed]
- Donzelli, B.; Krasnoff, S.B. Molecular genetics of secondary chemistry in Metarhizium fungi. Adv. Genet. 2016, 94, 365–436. [Google Scholar]
- Yadav, R.N.; Mahtab Rashid, M.; Zaidi, N.W.; Kumar, R.; Singh, H.B. Secondary Metabolites of Metarhizium spp. and Verticillium spp. and Their Agricultural Applications. Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms; Singh, H., Keswani, C., Reddy, M., Sansinenea, E., García-Estrada, C., Eds.; Springer: Singapore, 2019. [Google Scholar]
- Xu, Y.J.; Luo, F.; Li, B.; Shang, Y.F.; Wang, C.H. Metabolic conservation and diversification of Metarhizium species correlate with fungal host-specificity. Front. Microbiol. 2016, 7, 2020. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Chen, Y.; Li, B.Q.; Chen, T.; Tian, S.P. Reactive oxygen species: A generalist in regulating development and pathogenicity of phytopathogenic fungi. Comput. Struct. Biotec. 2020, 18, 3344–3349. [Google Scholar] [CrossRef]
- Marschall, R.; Tudzynski, P. Reactive oxygen species in development and infection processes. Semin. Cell Dev. Biol. 2016, 57, 138–146. [Google Scholar] [CrossRef]
- Fang, S.W.; Li, C.F.; Shih, D.Y.C. Antifungal activity of chitosan and its preservative effect on low-sugar Candied Kumquat. J. Food Prot. 1994, 57, 136–140. [Google Scholar] [CrossRef]
- Dai, Y.; Wu, X.Q.; Wang, Y.H.; Zhu, M.L. Biocontrol potential of Bacillus pumilus HR10 against Sphaeropsis shoot blight disease of pine. Biol. Control 2021, 152, 104458. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Zhang, X.; Wang, Q.; Li, N.; Ding, D.; Wang, B. Optimization of the Fermentation Conditions of Metarhizium robertsii and Its Biological Control of Wolfberry Root Rot Disease. Microorganisms 2023, 11, 2380. https://doi.org/10.3390/microorganisms11102380
He J, Zhang X, Wang Q, Li N, Ding D, Wang B. Optimization of the Fermentation Conditions of Metarhizium robertsii and Its Biological Control of Wolfberry Root Rot Disease. Microorganisms. 2023; 11(10):2380. https://doi.org/10.3390/microorganisms11102380
Chicago/Turabian StyleHe, Jing, Xiaoyan Zhang, Qinghua Wang, Nan Li, Dedong Ding, and Bin Wang. 2023. "Optimization of the Fermentation Conditions of Metarhizium robertsii and Its Biological Control of Wolfberry Root Rot Disease" Microorganisms 11, no. 10: 2380. https://doi.org/10.3390/microorganisms11102380
APA StyleHe, J., Zhang, X., Wang, Q., Li, N., Ding, D., & Wang, B. (2023). Optimization of the Fermentation Conditions of Metarhizium robertsii and Its Biological Control of Wolfberry Root Rot Disease. Microorganisms, 11(10), 2380. https://doi.org/10.3390/microorganisms11102380