
Assessment of Lipopeptide Mixtures Produced by Bacillus subtilis as Biocontrol Products against Apple Scab (Venturia inaequalis)

 , , , ,
, , , , 

Abstract
:1. Introduction
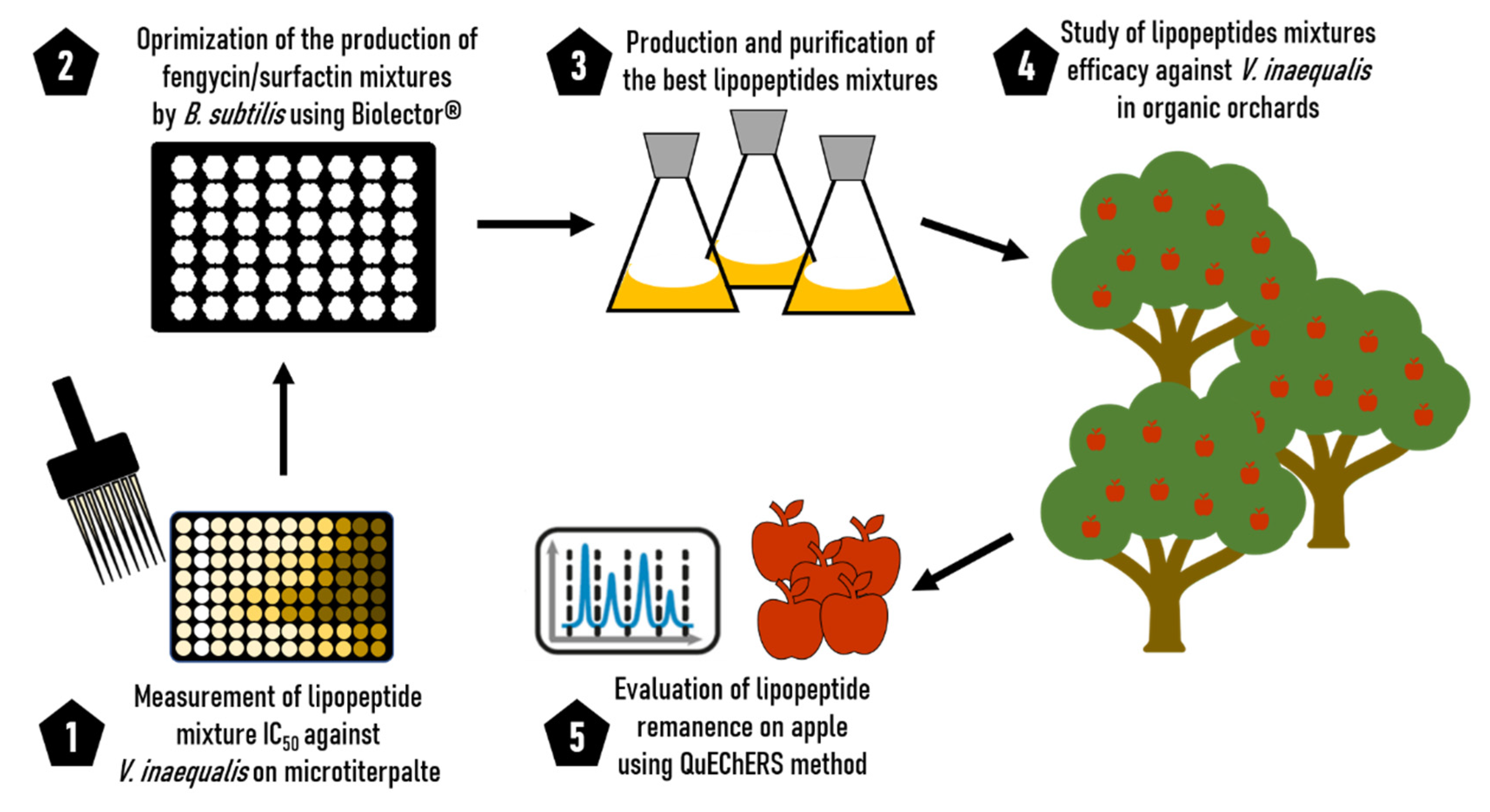
2. Materials and Methods
2.1. Lipopeptide Production and Purification for In Vitro Antifungal Assays
2.2. In Vitro Experiments against V. inaequalis
2.3. Medium Optimization for the Production of a Mix Fengycin/Surfactin by B. subtilis ATCC 21332
2.4. Quantification of Lipopeptides Using RP-UPLC
2.5. Lipopeptide Production for Orchard Trials
2.6. Protection of Apple Trees against Scab
2.7. Detection of Lipopeptide on Apple Using QuEChERS Method and Study of the Persistence of Lipopeptides
2.8. Quantification of the Lipopeptides by RP-UPLC and Mass Spectrometry (RP-UPLC-MS)
2.9. Statistical Analysis
3. Results
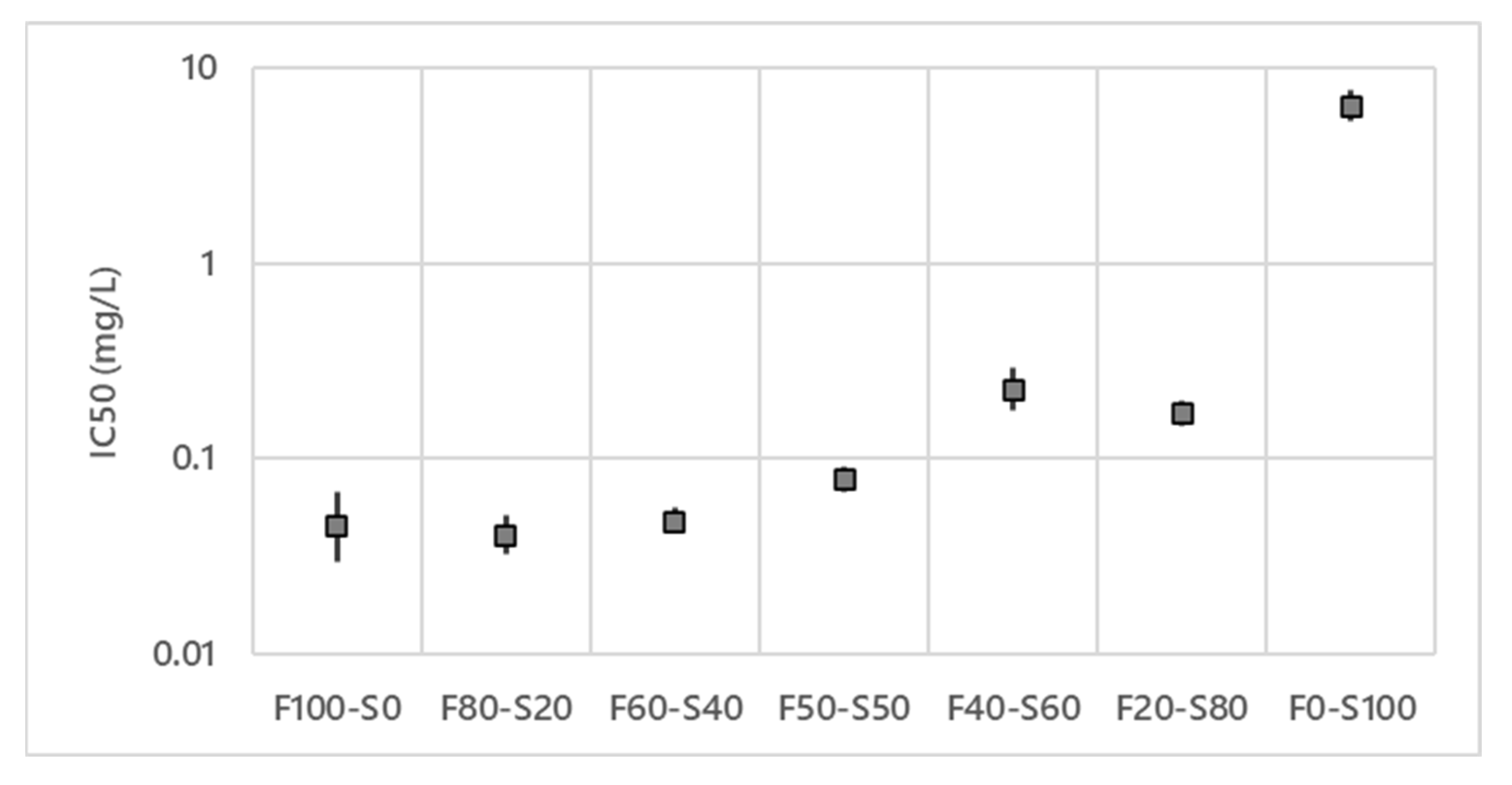
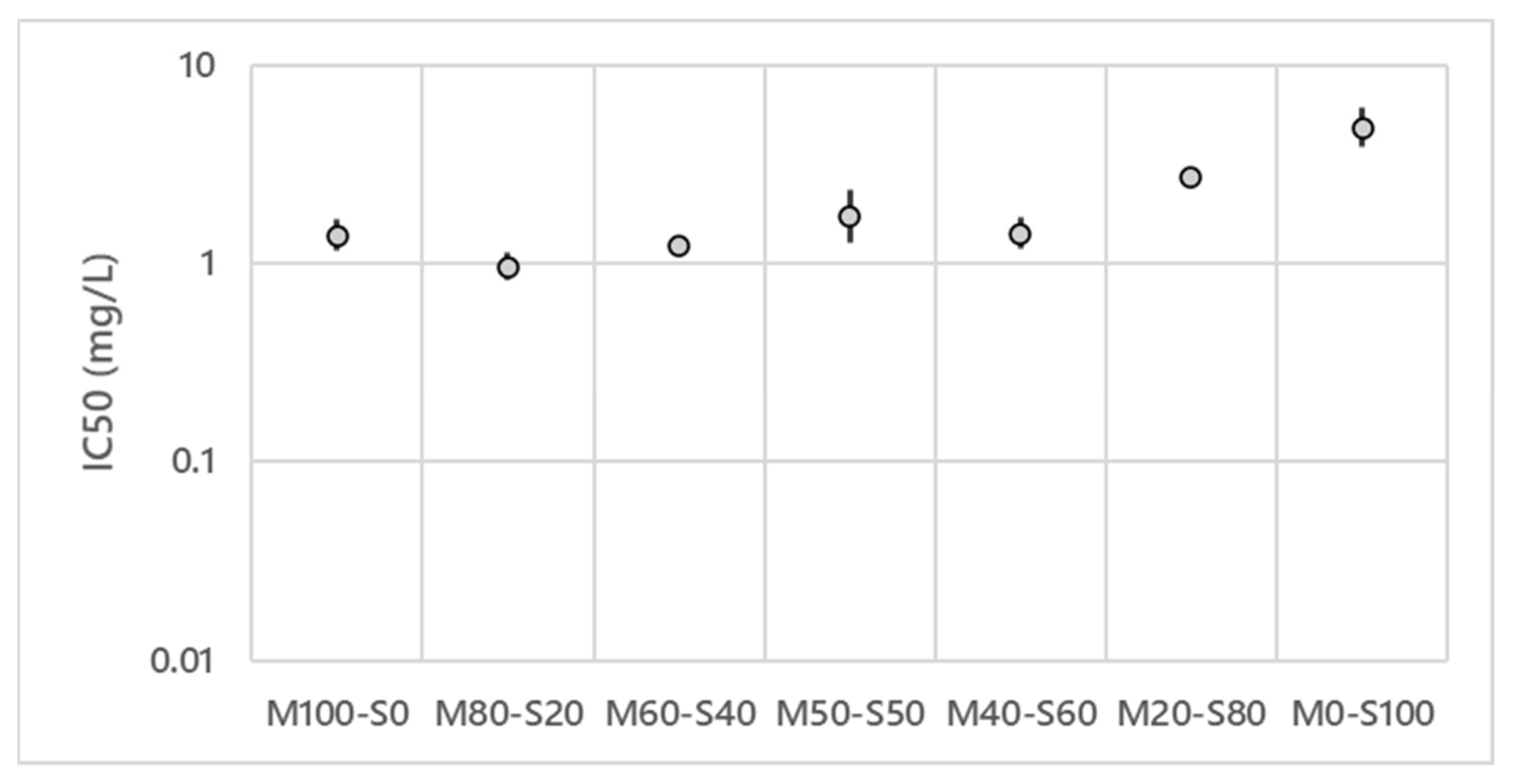
3.1. Antifungal Activities of the Mixture of Lipopeptides against V. inaequalis


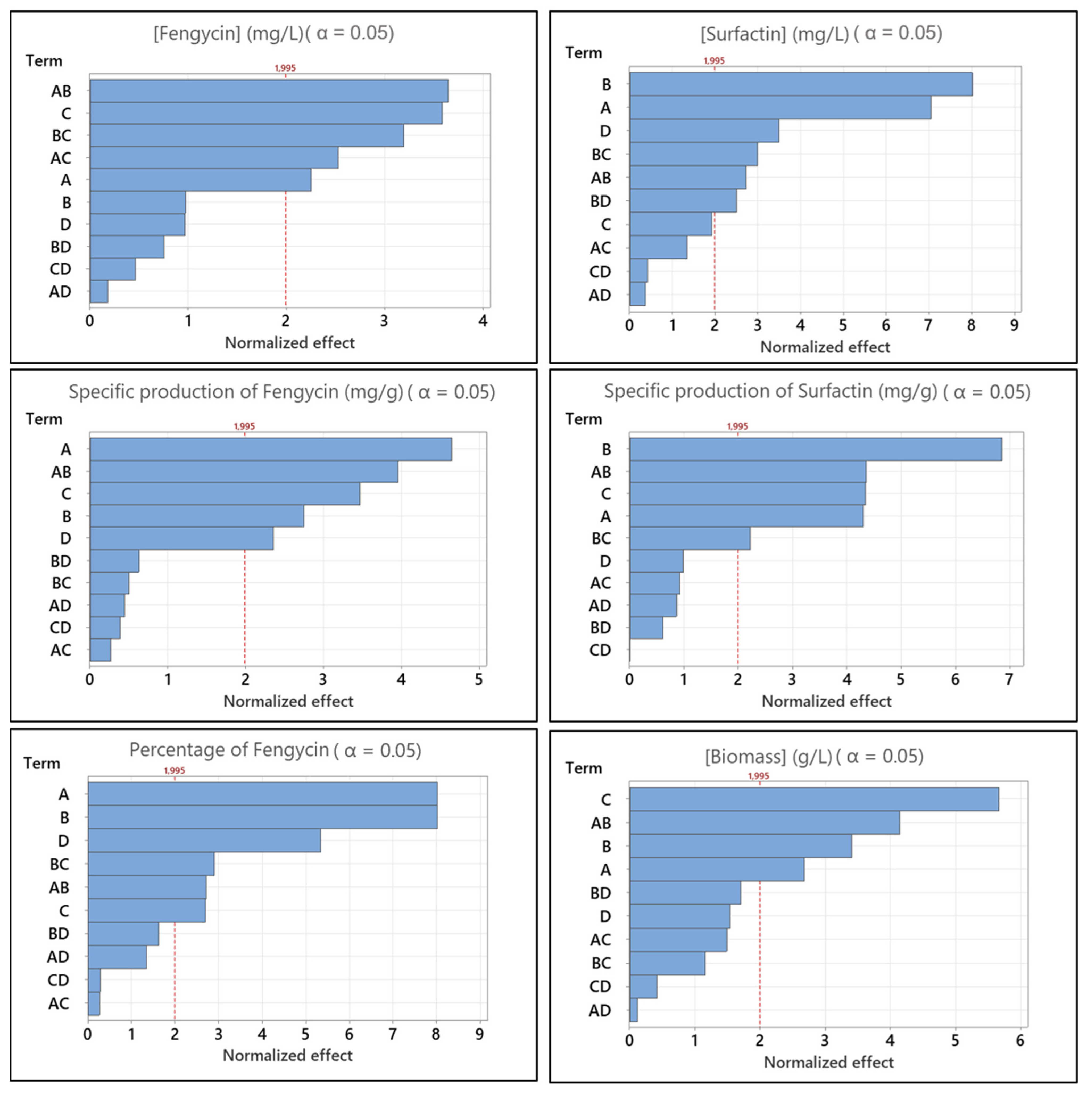
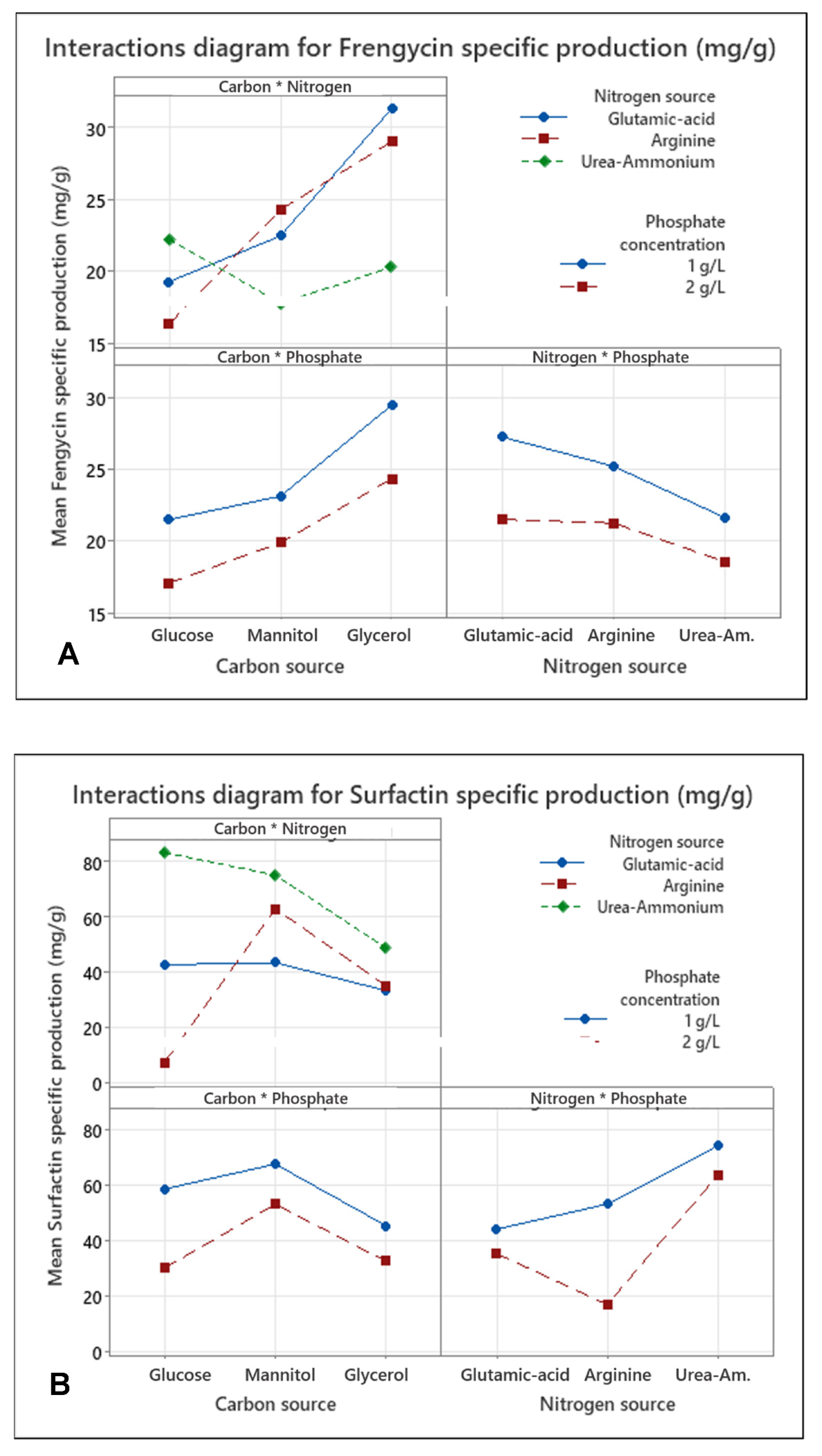
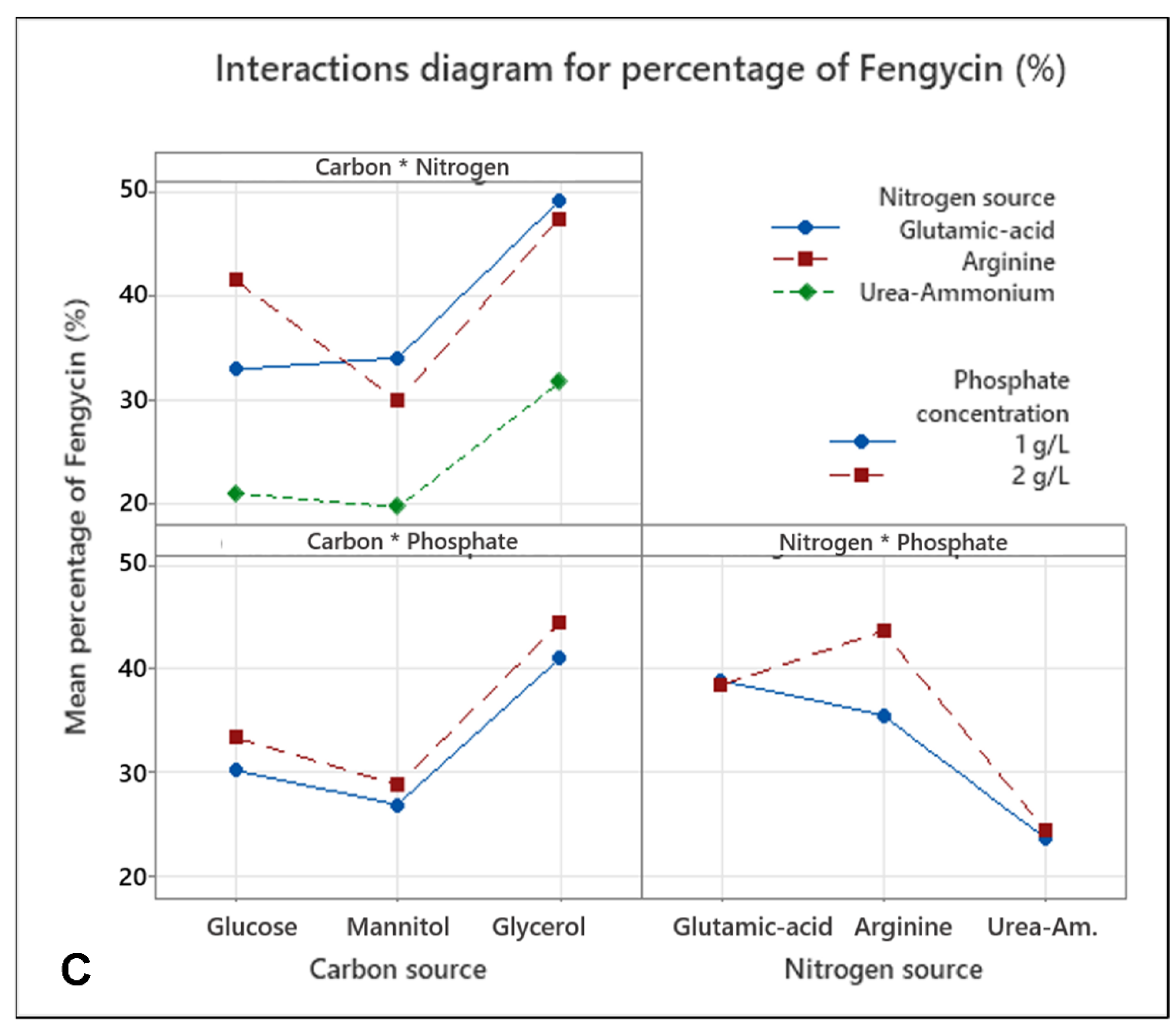
3.2. Medium Optimization Using DoE for the Production of Fengycin/surfactin Mixture by B. subtilis ATCC 21332
3.3. Protection of Apple Tree against Scab
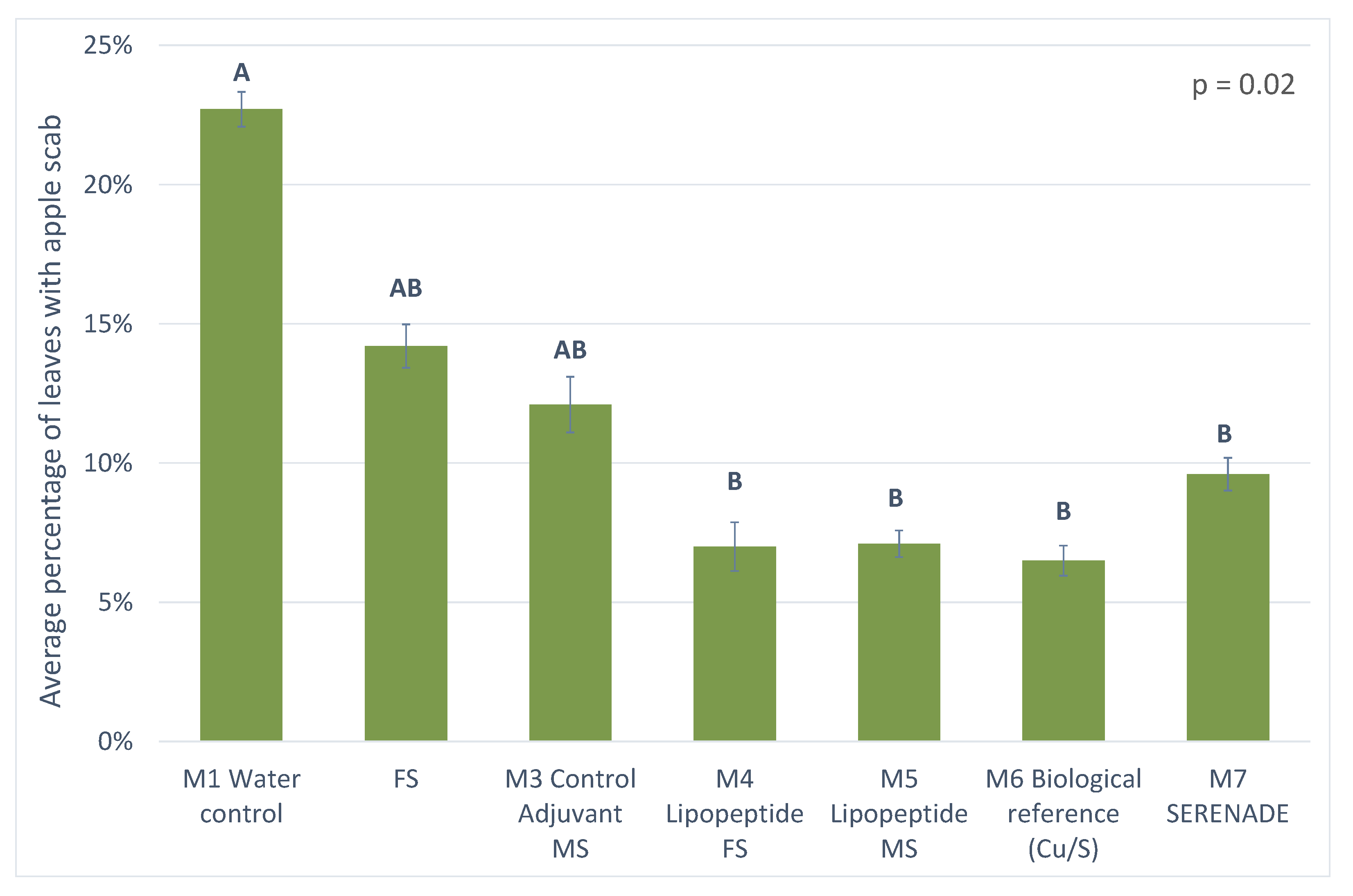
3.3.1. Results from 2018 Trial
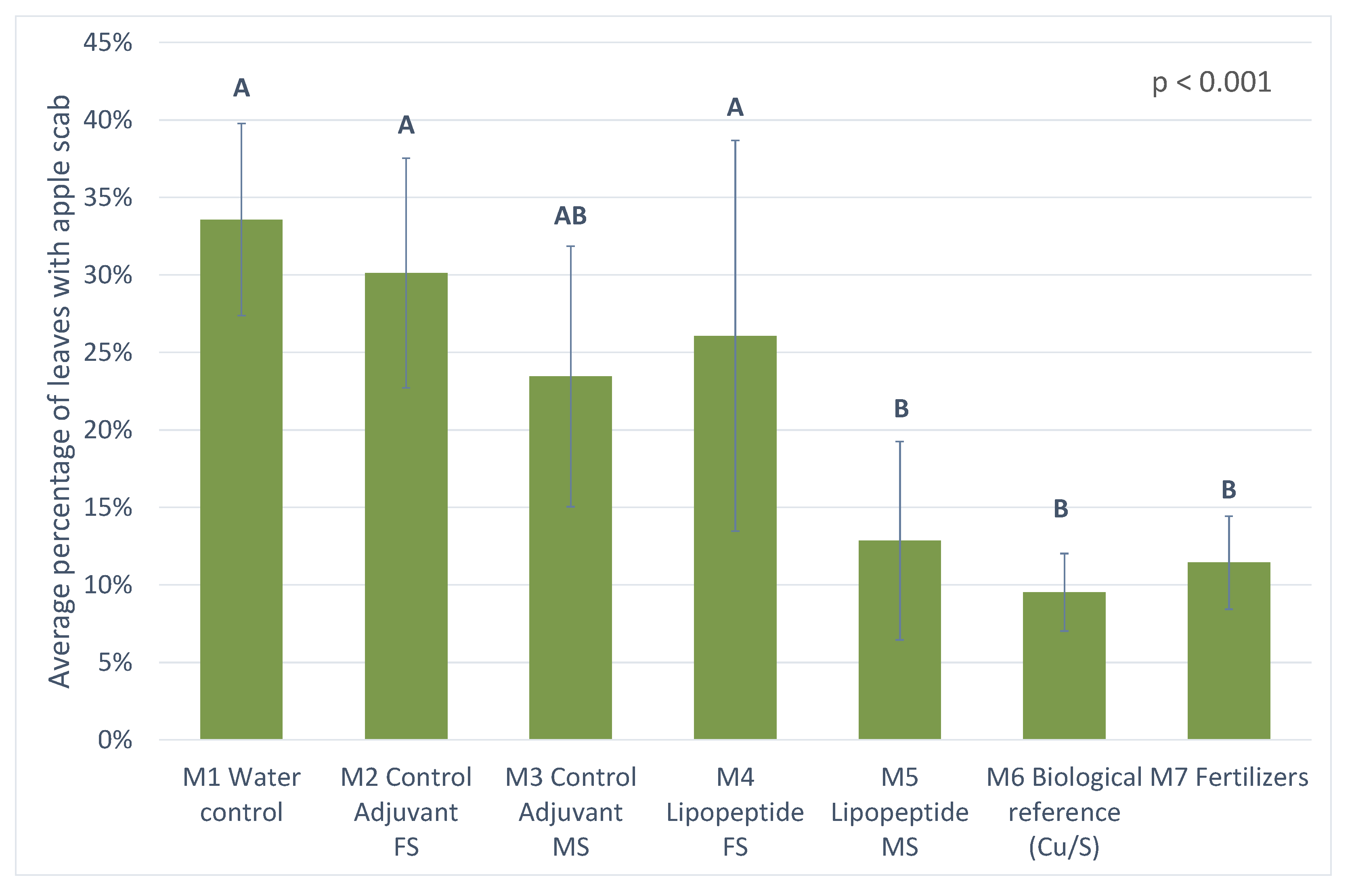
3.3.2. Results from 2019 Trial
3.4. Study of the Persistence of Molecules after Spraying on Fruit Trees
4. Discussion
4.1. In Vitro Antifungal Activities of the Lipopeptides Mixtures
4.2. Production of the Fengycin/Surfactin Mixture and Its Optimization by Design of Experiment
4.3. Protection against Apple Scab
4.4. Persistence of Lipopeptides on Apple Fruits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- MacHardy, W.E. Apple Scab: Biology, Epidemiology, and Management; APS Press: St. Paul, MN, USA, 1996. [Google Scholar]
- Holb, I.J.; Heijne, B.; Withagen, J.C.M.; Gáll, J.M.; Jeger, M.J. Analysis of Summer Epidemic Progress of Apple Scab at Different Apple Production Systems in the Netherlands and Hungary. Phytopathology 2005, 95, 1001–1020. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.K.; Mesarich, C.H.; Bus, V.G.M.; Beresford, R.M.; Plummer, K.M.; Templeton, M.D. Venturia inaequalis: The Causal Agent of Apple Scab: Venturia inaequalis. Mol. Plant Pathol. 2011, 12, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Chatzidimopoulos, M.; Lioliopoulou, F.; Sotiropoulos, T.; Vellios, E. Efficient Control of Apple Scab with Targeted Spray Applications. Agronomy 2020, 10, 217. [Google Scholar] [CrossRef]
- Cordero-Limon, L.; Shaw, M.W.; Passey, T.A.; Robinson, J.D.; Xu, X. Cross-resistance between myclobutanil and tebuconazole and the genetic basis of tebuconazole resistance in Venturia inaequalis. Pest Manag. Sci. 2020, 77, 844–850. [Google Scholar] [CrossRef]
- Shirane, N.; Takenaka, H.; Ueda, K.; Hashimoto, Y.; Katoh, K.; Ishii, H. Sterol analysis of DMI-resistant and -sensitive strains of Venturia inaequalis. Phytochemistry 1996, 41, 1301–1308. [Google Scholar] [CrossRef]
- Gao, L.; Berrie, A.; Yang, J.; Xu, X. Within- and between-Orchard Variability in the Sensitivity of Venturia inaequalis to Myclobutanil, a DMI Fungicide, in the UK. Pest Manag. Sci. 2009, 65, 1241–1249. [Google Scholar] [CrossRef]
- Li, X.; Li, H.; Yu, Z.; Gao, L.; Yang, J. Investigating the sensitivity of Venturia inaequalis isolates to difenoconazole and pyraclostrobin in apple orchards in China. Eur. J. Plant Pathol. 2021, 161, 207–217. [Google Scholar] [CrossRef]
- Villani, S.M.; Biggs, A.R.; Cooley, D.R.; Raes, J.J.; Cox, K.D. Prevalence of Myclobutanil Resistance and Difenoconazole Insensitivity in Populations of Venturia inaequalis. Plant Dis. 2015, 99, 1526–1536. [Google Scholar] [CrossRef]
- Xu, X.-M.; Gao, L.-Q.; Yang, J.-R. Are Insensitivities of Venturia inaequalis to Myclobutanil and Fenbuconazole Correlated? Crop Prot. 2010, 29, 183–189. [Google Scholar] [CrossRef]
- Yaegashi, H.; Hirayama, K.; Akahira, T.; Ito, T. Point mutation in CYP51A1 of Venturia inaequalis is associated with low sensitivity to sterol demethylation inhibitors. J. Gen. Plant Pathol. 2020, 86, 245–249. [Google Scholar] [CrossRef]
- European Food Safety Conclusion on the Peer Review of the Pesticide Risk Assessment of Confirmatory Data Submitted for the Active Substance Copper (I), Copper (II) Variants Namely Copper Hydroxide, Copper Oxychloride, Tribasic Copper Sulfate, Copper (I) Oxide, Bordeaux Mixture. EFSA J. 2013, 11, 3235.
- Coutte, F.; Niehren, J.; Dhali, D.; John, M.; Versari, C.; Jacques, P. Modeling Leucine’s Metabolic Pathway and Knockout Prediction Improving the Production of Surfactin, a Biosurfactant from Bacillus subtilis. Biotechnol. J. 2015, 10, 1216–1234. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Yin, Y.; Wen, J. Increasing fengycin production by strengthening the fatty acid synthesis pathway and optimizing fermentation conditions. Biochem. Eng. J. 2021, 177, 108235. [Google Scholar] [CrossRef]
- Jiang, J.; Han, M.; Fu, S.; Du, J.; Wang, S.; Zhang, H.; Li, W. Enhanced Production of Iturin A-2 Generated from Bacillus velezensis T701 and the Antitumor Activity of Iturin A-2 against Human Gastric Carcinoma Cells. Int. J. Pept. Res. Ther. 2021, 28, 27. [Google Scholar] [CrossRef]
- Zhao, X.; Han, Y.; Tan, X.-Q.; Wang, J.; Zhou, Z.-J. Optimization of antifungal lipopeptide production from Bacillus sp. BH072 by response surface methodology. J. Microbiol. 2014, 52, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Mizumoto, S.; Shoda, M. Medium optimization of antifungal lipopeptide, iturin A, production by Bacillus subtilis in solid-state fermentation by response surface methodology. Appl. Microbiol. Biotechnol. 2007, 76, 101–108. [Google Scholar] [CrossRef]
- Akpa, E.; Jacques, P.; Wathelet, B.; Paquot, M.; Fuchs, R.; Budzikiewicz, H.; Thonart, P. Influence of Culture Conditions on Lipopeptide Production by Bacillus subtilis. Appl. Biochem. Biotechnol. 2001, 91, 551–561. [Google Scholar] [CrossRef]
- Huang, X.; Lu, Z.; Zhao, H.; Bie, X.; Lü, F.; Yang, S. Antiviral Activity of Antimicrobial Lipopeptide from Bacillus subtilis fmbj Against Pseudorabies Virus, Porcine Parvovirus, Newcastle Disease Virus and Infectious Bursal Disease Virus in Vitro. Int. J. Pept. Res. Ther. 2006, 12, 373–377. [Google Scholar] [CrossRef]
- Chen, M.; Wang, J.; Zhu, Y.; Liu, B.; Yang, W.; Ruan, C. Antibacterial activity against Ralstonia solanacearum of the lipopeptides secreted from the Bacillus amyloliquefaciens strain FJAT-2349. J. Appl. Microbiol. 2019, 126, 1519–1529. [Google Scholar] [CrossRef]
- Zhao, H.; Shao, D.; Jiang, C.; Shi, J.; Li, Q.; Huang, Q.; Rajoka, M.S.R.; Yang, H.; Jin, M. Biological activity of lipopeptides from Bacillus. Appl. Microbiol. Biotechnol. 2017, 101, 5951–5960. [Google Scholar] [CrossRef]
- Moyne, A.-L.; Shelby, R.; Cleveland, T.E.; Tuzun, S. Bacillomycin D: An iturin with antifungal activity against Aspergillus flavus. J. Appl. Microbiol. 2001, 90, 622–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desmyttere, H.; Deweer, C.; Muchembled, J.; Sahmer, K.; Jacquin, J.; Coutte, F.; Jacques, P. Antifungal Activities of Bacillus subtilis Lipopeptides to Two Venturia inaequalis Strains Possessing Different Tebuconazole Sensitivity. Front. Microbiol. 2019, 10, 2327. [Google Scholar] [CrossRef] [PubMed]
- Jacques, P. Surfactin and Other Lipopeptides from Bacillus Spp. In Biosurfactants; Soberón-Chávez, G., Ed.; Microbiology Monographs; Springer: Berlin/Heidelberg, Germany, 2011; Volume 20, pp. 57–91. [Google Scholar]
- Krishnan, N.; Velramar, B.; Velu, R.K. Investigation of antifungal activity of surfactin against mycotoxigenic phytopathogenic fungus Fusarium moniliforme and its impact in seed germination and mycotoxicosis. Pestic. Biochem. Physiol. 2019, 155, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Mnif, I.; Ghribi, G. Lipopeptide Surfactants: Production, Recovery and Pore Forming Capacity. Peptides 2015, 71, 100–112. [Google Scholar]
- Liu, J.; Hagberg, I.; Novitsky, L.; Hadj-Moussa, H.; Avis, T.J. Interaction of antimicrobial cyclic lipopeptides from Bacillus subtilis influences their effect on spore germination and membrane permeability in fungal plant pathogens. Fungal Biol. 2014, 118, 855–861. [Google Scholar] [CrossRef]
- Deravel, J.; Lemière, S.; Coutte, F.; Krier, F.; Van Hese, N.; Béchet, M.; Sourdeau, N.; Höfte, M.; Leprêtre, A.; Jacques, P. Mycosubtilin and Surfactin Are Efficient, Low Ecotoxicity Molecules for the Biocontrol of Lettuce Downy Mildew. Appl. Microbiol. Biotechnol. 2014, 98, 6255–6264. [Google Scholar] [CrossRef]
- Ben Ayed, H.; Nasri, R.; Jemil, N.; Ben Amor, I.; Gargouri, J.; Hmidet, N.; Nasri, M. Acute and sub-chronic oral toxicity profiles of lipopeptides from Bacillus mojavensis A21 and evaluation of their in vitro anticoagulant activity. Chem. Interactions 2015, 236, 1–6. [Google Scholar] [CrossRef]
- Mnif, I.; Ghribi, D. Review lipopeptides biosurfactants: Mean classes and new insights for industrial, biomedical, and environmental applications. Biopolymers 2015, 104, 129–147. [Google Scholar] [CrossRef]
- Cawoy, H.; Mariutto, M.; Henry, G.; Fisher, C.; Vasilyeva, N.; Thonart, P.; Dommes, J.; Ongena, M. Plant Defense Stimulation by Natural Isolates of Bacillus Depends on Efficient Surfactin Production. Mol. Plant-Microbe Interact. 2014, 27, 87–100. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Deleu, M.; Paquot, M.; Nylander, T. Effect of Fengycin, a Lipopeptide Produced by Bacillus subtilis, on Model Biomembranes. Biophys. J. 2008, 94, 2667–2679. [Google Scholar] [CrossRef]
- Falardeau, J.; Wise, C.; Novitsky, L.; Avis, T.J. Ecological and Mechanistic Insights Into the Direct and Indirect Antimicrobial Properties of Bacillus subtilis Lipopeptides on Plant Pathogens. J. Chem. Ecol. 2013, 39, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Mahata, D.; Paul, D.; Korpole, S.; Franco, O.L.; Mandal, S.M. Purification, biochemical characterization and self-assembled structure of a fengycin-like antifungal peptide from Bacillus thuringiensis strain SM1. Front. Microbiol. 2013, 4, 332. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Han, J.; Liu, H.; Qu, X.; Lu, Z.; Bie, X. Plipastatin and surfactin coproduction by Bacillus subtilis pB2-L and their effects on microorganisms. Antonie Van Leeuwenhoek 2017, 110, 1007–1018. [Google Scholar] [CrossRef]
- Fan, H.; Ru, J.; Zhang, Y.; Wang, Q.; Li, Y. Fengycin produced by Bacillus subtilis 9407 plays a major role in the biocontrol of apple ring rot disease. Microbiol. Res. 2017, 199, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Xie, J.; Qi, Y.; Qin, Q.; Jin, C.; Wang, B.; Fang, W. A Thermotolerant Marine Bacillus amyloliquefaciens S185 Producing Iturin A5 for Antifungal Activity against Fusarium oxysporum f. Sp. Cubense. Mar. Drugs 2021, 19, 516. [Google Scholar] [CrossRef] [PubMed]
- Farzaneh, M.; Shi, Z.-Q.; Ahmadzadeh, M.; Hu, L.-B.; Ghassempour, A. Inhibition of the Aspergillus flavus Growth and Aflatoxin B1 Contamination on Pistachio Nut by Fengycin and Surfactin-Producing Bacillus subtilis UTBSP1. Plant Pathol. J. 2016, 32, 209–215. [Google Scholar] [CrossRef]
- Mejri, S.; Siah, A.; Coutte, F.; Magnin-Robert, M.; Randoux, B.; Tisserant, B.; Krier, F.; Jacques, P.; Reignault, P.; Halama, P. Biocontrol of the wheat pathogen Zymoseptoria tritici using cyclic lipopeptides from Bacillus subtilis. Environ. Sci. Pollut. Res. 2017, 25, 29822–29833. [Google Scholar] [CrossRef]
- Toral, L.; Rodríguez, M.; Béjar, V.; Sampedro, I. Antifungal Activity of Lipopeptides From Bacillus XT1 CECT 8661 Against Botrytis cinerea. Front. Microbiol. 2018, 9, 1315. [Google Scholar] [CrossRef]
- Fickers, P.; Leclère, V.; Guez, J.-S.; Béchet, M.; Coucheney, F.; Joris, B.; Jacques, P. Temperature dependence of mycosubtilin homologue production in Bacillus subtilis ATCC6633. Res. Microbiol. 2008, 159, 449–457. [Google Scholar] [CrossRef]
- Fahim, S.; Dimitrov, K.; Gancel, F.; Vauchel, P.; Jacques, P.; Nikov, I. Impact of energy supply and oxygen transfer on selective lipopeptide production by Bacillus subtilis BBG21. Bioresour. Technol. 2012, 126, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Coutte, F.; Lecouturier, D.; Yahia, S.A.; Leclère, V.; Béchet, M.; Jacques, P.; Dhulster, P. Production of surfactin and fengycin by Bacillus subtilis in a bubbleless membrane bioreactor. Appl. Microbiol. Biotechnol. 2010, 87, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Yaseen, Y.; Gancel, F.; Béchet, M.; Drider, D.; Jacques, P. Study of the correlation between fengycin promoter expression and its production by Bacillus subtilis under different culture conditions and the impact on surfactin production. Arch. Microbiol. 2017, 199, 1371–1382. [Google Scholar] [CrossRef] [PubMed]
- Ghribi, D.; Ellouze-Chaabouni, S. Enhancement of Bacillus subtilis Lipopeptide Biosurfactants Production through Optimization of Medium Composition and Adequate Control of Aeration. Biotechnol. Res. Int. 2011, 2011, 1–6. [Google Scholar] [CrossRef]
- Etchegaray, A.; Coutte, F.; Chataigné, G.; Béchet, M.; dos Santos, R.H.Z.; Leclère, V.; Jacques, P. Production of Bacillus amyloliquefaciens OG and Its Metabolites in Renewable Media: Valorisation for Biodiesel Production and p-Xylene Decontamination. Can. J. Microbiol. 2017, 63, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Jacques, P.; Hbid, C.; Destain, J.; Razafindralambo, H.; Paquot, M.; De Pauw, E.; Thonart, P. Optimization of Biosurfactant Lipopeptide Production from Bacillus Subtilis S499 by Plackett-Burman Design. In Twentieth Symposium on Biotechnology for Fuels and Chemicals: Presented as Volumes 77–79 of Applied Biochemistry and Biotechnology; Davison, B.H., Finkelstein, M., Eds.; Applied Biochemistry and Biotechnology; Humana Press: Totowa, NJ, USA, 1999; pp. 223–233. [Google Scholar]
- Motta Dos Santos, L.F.; Coutte, F.; Ravallec, R.; Dhulster, P.; Tournier-Couturier, L.; Jacques, P. An Improvement of Surfactin Production by Bacillus subtilis BBG131 Using Design of Experiments in Microbioreactors and Continuous Process in Bubbleless Membrane Bioreactor. Bioresour. Technol. 2016, 218, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.-H.; Lai, C.-C.; Chang, J.-S. Using Taguchi experimental design methods to optimize trace element composition for enhanced surfactin production by Bacillus subtilis ATCC 21332. Process Biochem. 2007, 42, 40–45. [Google Scholar] [CrossRef]
- Vassaux, A.; Rannou, M.; Peers, S.; Daboudet, T.; Jacques, P.; Coutte, F. Impact of the Purification Process on the Spray-Drying Performances of the Three Families of Lipopeptide Biosurfactant Produced by Bacillus subtilis. Front. Bioeng. Biotechnol. 2021, 9, 815337. [Google Scholar] [CrossRef] [PubMed]
- Muchembled, J.; Deweer, C.; Sahmer, K.; Halama, P. Antifungal activity of essential oils on two Venturia inaequalis strains with different sensitivities to tebuconazole. Environ. Sci. Pollut. Res. 2017, 25, 29921–29928. [Google Scholar] [CrossRef]
- Kensy, F.; Zang, E.; Faulhammer, C.; Tan, R.-K.; Büchs, J. Validation of a high-throughput fermentation system based on online monitoring of biomass and fluorescence in continuously shaken microtiter plates. Microb. Cell Factories 2009, 8, 31. [Google Scholar] [CrossRef]
- Funke, M.; Diederichs, S.; Kensy, F.; Müller, C.; Büchs, J. The baffled microtiter plate: Increased oxygen transfer and improved online monitoring in small scale fermentations. Biotechnol. Bioeng. 2009, 103, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Tiryaki, O. Validation of QuEChERS method for the determination of some pesticide residues in two apple varieties. J. Environ. Sci. Health Part B 2016, 51, 722–729. [Google Scholar] [CrossRef]
- Romero, D.; De Vicente, A.; Rakotoaly, R.H.; Dufour, S.E.; Veening, J.-W.; Arrebola, E.; Cazorla, F.M.; Kuipers, O.P.; Paquot, M.; Pérez-García, A. The Iturin and Fengycin Families of Lipopeptides Are Key Factors in Antagonism of Bacillus subtilis Toward Podosphaera fusca. Mol. Plant Microbe Interact. 2007, 20, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, M.; Castillo, F.; Alcal, E.; Morales, G.; Valdes, R.; Reyes, F. Biological Effectiveness of Bacillus Spp. and Trichoderma Spp. on Apple Scab (Venturia inaequalis) in Vitro and under Field Conditions. Eur. J. Phys. Agric. Sci. 2018, 6, 11. [Google Scholar]
- Kucheryava, N.; Fiss, M.; Auling, G.; Kroppenstedt, R.M. Isolation and Characterization of Epiphytic Bacteria from the Phyllosphere of Apple, Antagonistic in vitro to Venturia inaequalis, the Causal Agent of Apple Scab. Syst. Appl. Microbiol. 1999, 22, 472–478. [Google Scholar] [CrossRef]
- Köhl, J.J.; Molhoek, W.W.M.L.; Haas, B.B.H.G.-D.; De Geijn, H.H.M.G.-V. Selection and orchard testing of antagonists suppressing conidial production by the apple scab pathogen Venturia inaequalis. Eur. J. Plant Pathol. 2008, 123, 401–414. [Google Scholar] [CrossRef]
- Köhl, J.; Scheer, C.; Holb, I.J.; Masny, S.; Molhoek, W. Toward an Integrated Use of Biological Control by Cladosporium cladosporioides H39 in Apple Scab (Venturia inaequalis) Management. Plant Dis. 2015, 99, 535–543. [Google Scholar] [CrossRef]
- Saxena, A.k.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.j. Bacillus Species in Soil as a Natural Resource for Plant Health and Nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef]
- Kensy, F.; Engelbrecht, C.; Büchs, J. Scale-up from microtiter plate to laboratory fermenter: Evaluation by online monitoring techniques of growth and protein expression in Escherichia coli and Hansenula polymorpha fermentations. Microb. Cell Factories 2009, 8, 68. [Google Scholar] [CrossRef]
- Janek, T.; Gudiña, E.; Połomska, X.; Biniarz, P.; Jama, D.; Rodrigues, L.; Rymowicz, W.; Lazar, Z. Sustainable Surfactin Production by Bacillus subtilis Using Crude Glycerol from Different Wastes. Molecules 2021, 26, 3488. [Google Scholar] [CrossRef]
- Sousa, M.; Melo, V.; Rodrigues, S.; Sant’Ana, H.B.; Gonçalves, L.R.B. Screening of biosurfactant-producing Bacillus strains using glycerol from the biodiesel synthesis as main carbon source. Bioprocess Biosyst. Eng. 2012, 35, 897–906. [Google Scholar] [CrossRef] [PubMed]
- de Faria, A.F.; Stéfani, D.; Vaz, B.G.; Silva, Í.S.; Garcia, J.S.; Eberlin, M.N.; Grossman, M.J.; Alves, O.L.; Durrant, L.R. Purification and Structural Characterization of Fengycin Homologues Produced by Bacillus subtilis LSFM-05 Grown on Raw Glycerol. J. Ind. Microbiol. Biotechnol. 2011, 38, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Denoirjean, T.; Doury, G.; Poli, P.; Coutte, F.; Ameline, A. Effects of Bacillus lipopeptides on the survival and behavior of the rosy apple aphid Dysaphis plantaginea. Ecotoxicol. Environ. Saf. 2021, 226, 112840. [Google Scholar] [CrossRef] [PubMed]
- Denoirjean, T.; Ameline, A.; Couty, A.; Dubois, F.; Coutte, F.; Doury, G. Effects of surfactins, Bacillus lipopeptides, on the behavior of an aphid and host selection by its parasitoid. Pest Manag. Sci. 2021, 78, 929–937. [Google Scholar] [CrossRef]
- Toure, Y.; Ongena, M.; Jacques, P.; Guiro, A.; Thonart, P. Role of lipopeptides produced by Bacillus subtilis GA1 in the reduction of grey mould disease caused by Botrytis cinerea on apple. J. Appl. Microbiol. 2004, 96, 1151–1160. [Google Scholar] [CrossRef]
- Kourmentza, K.; Gromada, X.; Michael, N.; Degraeve, C.; Vanier, G.; Ravallec, R.; Coutte, F.; Karatzas, K.A.; Jauregi, P. Antimicrobial Activity of Lipopeptide Biosurfactants Against Foodborne Pathogen and Food Spoilage Microorganisms and Their Cytotoxicity. Front. Microbiol. 2021, 11, 561060. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipopeptide(s) | Ratios (%) | Code | Purity (%) |
|---|---|---|---|
| Fengycin | 100–0 | F100 | 98 |
| Fengycin–Surfactin | 80–20 | F80–S20 | 98–90 |
| 60–40 | F60–S40 | ||
| 50–50 | F50–S50 | ||
| 40–60 | F40–S60 | ||
| 20–80 | F20–S80 | ||
| Surfactin | 0–100 | S100 | 90 |
| Mycosubtilin | 100–0 | M100 | 75 |
| Mycosubtilin–Surfactin | 80–20 | M80–S20 | 75–90 |
| 60–40 | M60–S40 | ||
| 50–50 | M50–S50 | ||
| 40–60 | M40–S60 | ||
| 20–80 | M20–S80 | ||
| Surfactin | 0–100 | S100 | 90 |
| Modalities | Pre-Flowering | Post-Flowering | ||||
|---|---|---|---|---|---|---|
| Substances | Dose | Applications | Substances | Dose | Applications | |
| M1 Water control | Water | T1 to T5 | Water | T6 to T11 | ||
| M2 Control Adjuvant Lipopeptides 1 (Fengycin/Surfactin) | Copper | 1.5 kg/ha | T1 to T2 | Adjuvant | 2 L/ha | |
| M3 Control Adjuvant Lipopeptides 2 (Mycosubtilin/Surfactin) | Adjuvant | |||||
| M4 Lipopeptides 1 (Fengycin/Surfactin) | Sulphur | 7.5 kg/ha | T3 to T4 | 55–45% | 250 mg/L | |
| M5 Lipopeptides 2 (Mycosubtilin/Surfactin) | 80–20% | 500 mg/L | ||||
| M6 Biological reference (Cu/S) | Sulphur + Copper | 5 kg/ha + 1 kg/ha | T5 | Sulphur + Copper | 5 kg/ha + 1 kg/ha | |
| M7 SERENADE | B. subtilis str. QST 713 | 2 kg/ha | ||||
| Modalities | Substances | Dose | Applications |
|---|---|---|---|
| M1 Water control | Water | - | T1 to T12 |
| M2 Control Adjuvant Lipopeptides 1 (Fengycin/Surfactin) | Adjuvant | 2 L/ha | T1 to T12 |
| M3 Control Adjuvant Lipopeptides 2 (Mycosubtilin/Surfactin) | Adjuvant | - | T1 to T12 |
| M4 Lipopeptides 1 (Fengycin/Surfactin) | 55–45% | 250 mg/L | T1 to T12 |
| M5 Lipopeptides 2 (Mycosubtilin/Surfactin) | 80–20% | 500 mg/L | T1 to T12 |
| M6 Biological reference (Cu/S) | Copper | 1.5 kg/ha | T1 |
| Sulphur | 7.5 kg/ha | T2 and T4 | |
| Sulphur + Copper | 5 kg/ha + 1 kg/ha | T5 to T12 | |
| M7 Fertilizers | Copper | 1.5 kg/ha | T1 |
| Fertilizer 1 | 2% to 4% | T2 to T4 | |
| Fertilizer 2 | 8% | T5 to T12 |
| Modalities | Fengycin | Surfactin | Mycosubtilin | |
|---|---|---|---|---|
| Control | Skin | |||
| Flesh | ||||
| Fengycin–Surfactin | Skin | |||
| Flesh | ||||
| Mycosubtilin–Surfactin | Skin | |||
| Flesh |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leconte, A.; Tournant, L.; Muchembled, J.; Paucellier, J.; Héquet, A.; Deracinois, B.; Deweer, C.; Krier, F.; Deleu, M.; Oste, S.; et al. Assessment of Lipopeptide Mixtures Produced by Bacillus subtilis as Biocontrol Products against Apple Scab (Venturia inaequalis). Microorganisms 2022, 10, 1810. https://doi.org/10.3390/microorganisms10091810
Leconte A, Tournant L, Muchembled J, Paucellier J, Héquet A, Deracinois B, Deweer C, Krier F, Deleu M, Oste S, et al. Assessment of Lipopeptide Mixtures Produced by Bacillus subtilis as Biocontrol Products against Apple Scab (Venturia inaequalis). Microorganisms. 2022; 10(9):1810. https://doi.org/10.3390/microorganisms10091810
Chicago/Turabian StyleLeconte, Aline, Ludovic Tournant, Jérôme Muchembled, Jonathan Paucellier, Arnaud Héquet, Barbara Deracinois, Caroline Deweer, François Krier, Magali Deleu, Sandrine Oste, and et al. 2022. "Assessment of Lipopeptide Mixtures Produced by Bacillus subtilis as Biocontrol Products against Apple Scab (Venturia inaequalis)" Microorganisms 10, no. 9: 1810. https://doi.org/10.3390/microorganisms10091810
APA StyleLeconte, A., Tournant, L., Muchembled, J., Paucellier, J., Héquet, A., Deracinois, B., Deweer, C., Krier, F., Deleu, M., Oste, S., Jacques, P., & Coutte, F. (2022). Assessment of Lipopeptide Mixtures Produced by Bacillus subtilis as Biocontrol Products against Apple Scab (Venturia inaequalis). Microorganisms, 10(9), 1810. https://doi.org/10.3390/microorganisms10091810