
Characterization of Bacillus velezensis UTB96, Demonstrating Improved Lipopeptide Production Compared to the Strain B. velezensis FZB42

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial and Fungal Strains
2.2. Primers and DNA Sequencing
2.3. LC-MS/MS Analyses of Lipopeptides
2.4. Media and Cultivation Procedure
2.4.1. Shake Flask Cultivations
2.4.2. Bioreactor Cultivations
2.5. Lipopeptide Extraction and Quantitative Analysis
2.6. Evaluation of Antifungal Activity
2.7. Data Analysis
3. Results
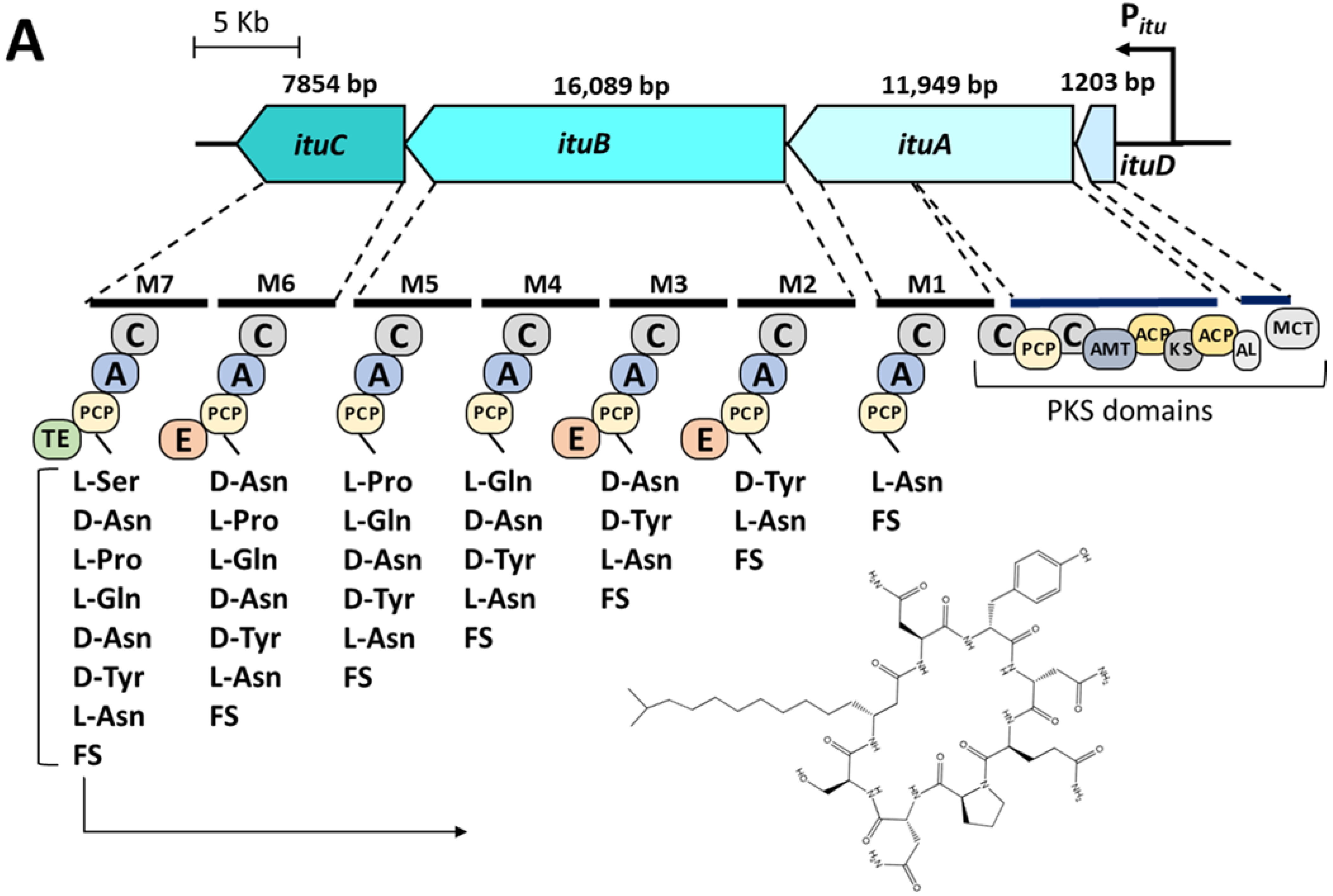
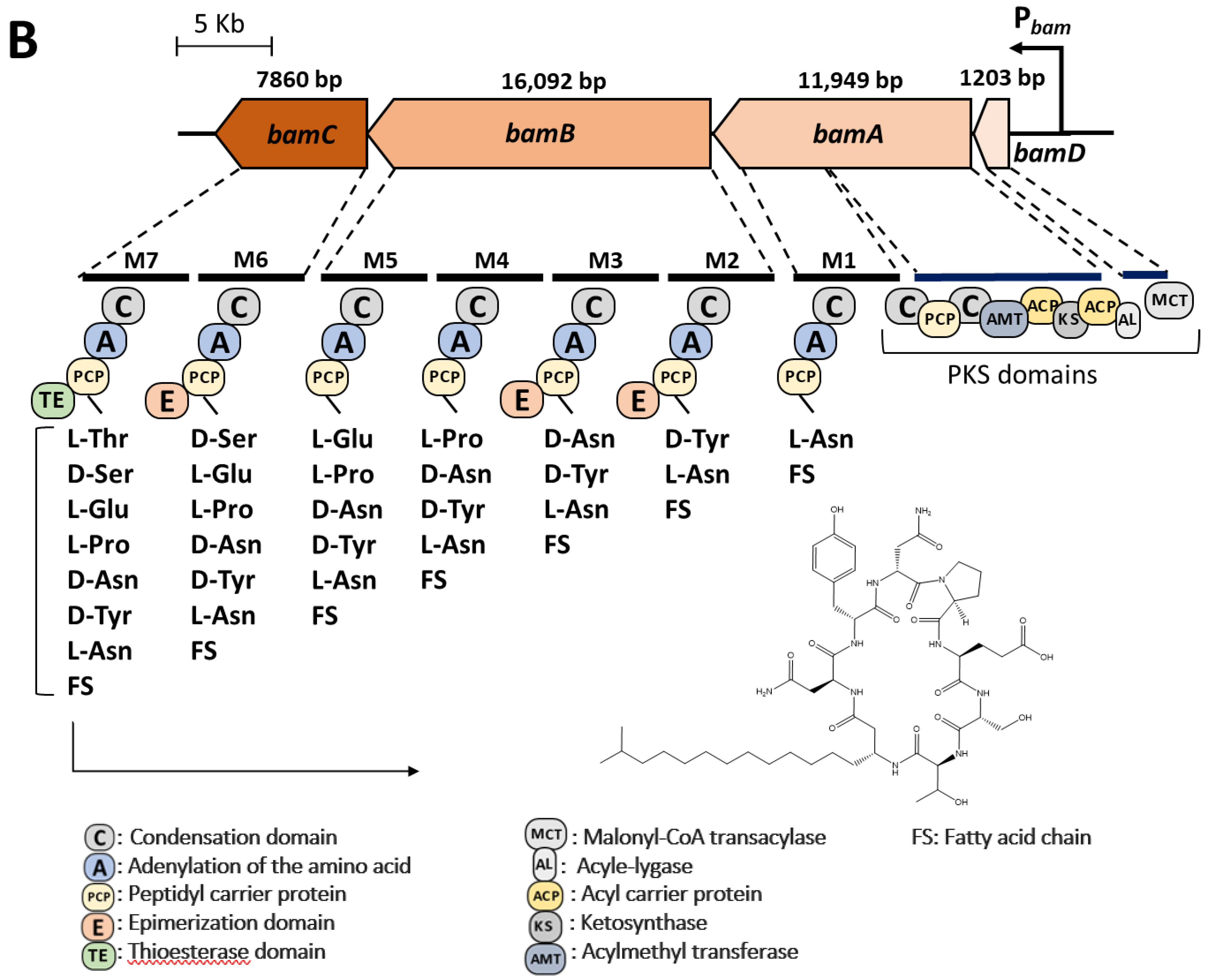
3.1. Bioinformatic Analyses of Lipopeptide Biosynthesis
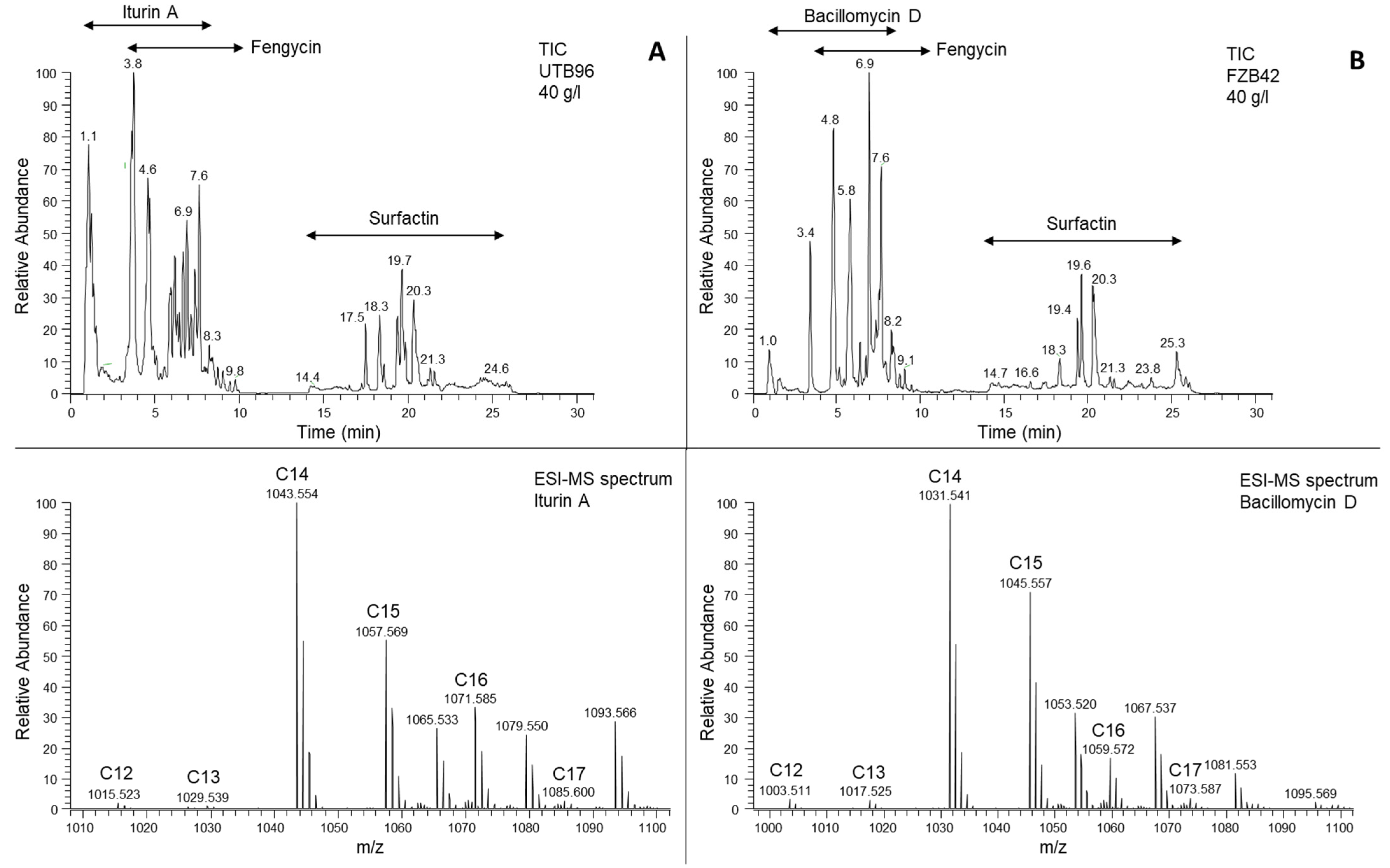
3.2. Comparative Structure-Based Iturin A and Bacillomycin D Analysis by Mass Spectrometry
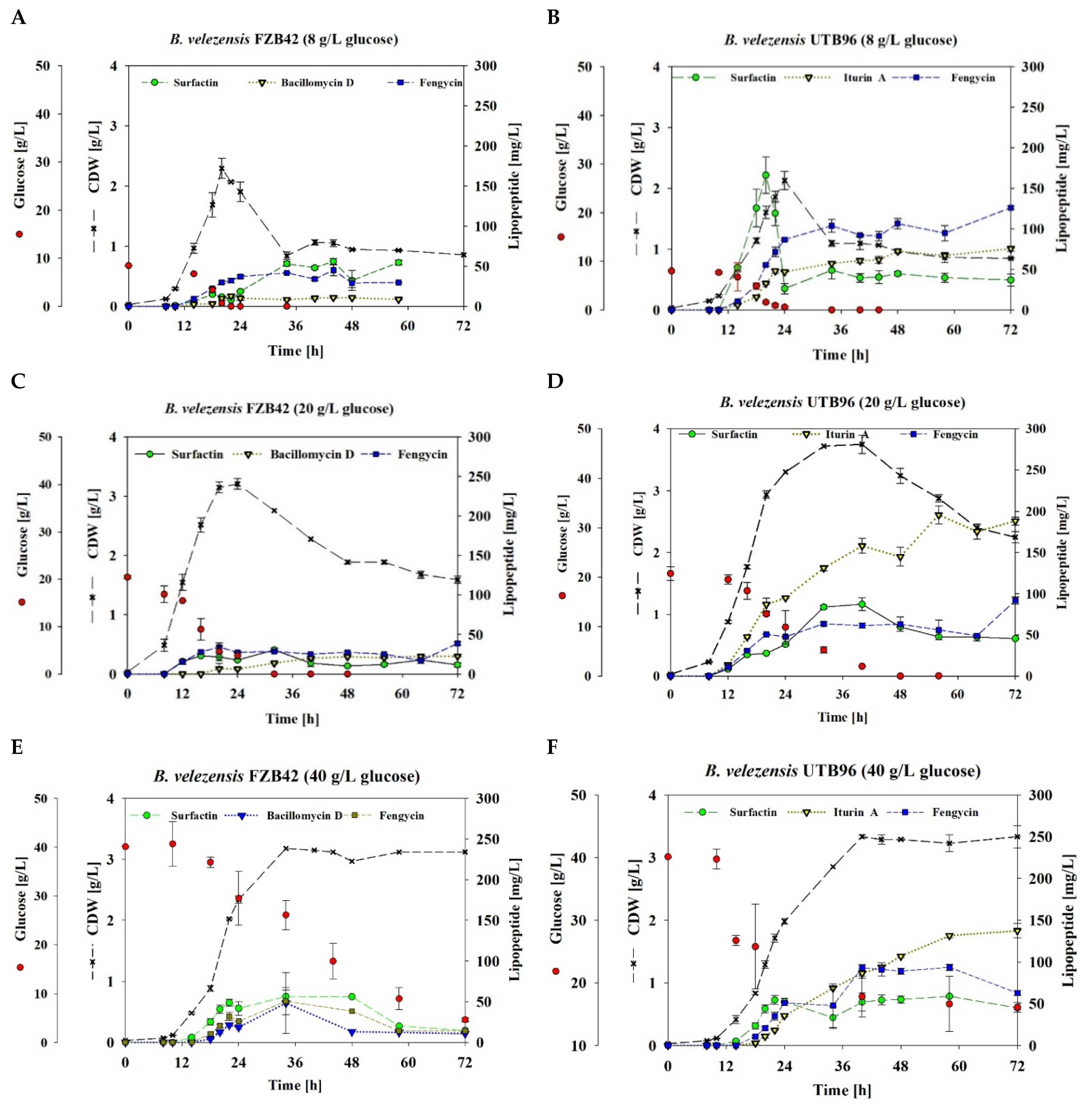
3.3. Lipopeptide Production of B. velezensis UTB96 and FZB42 under Varying Substrate Availability
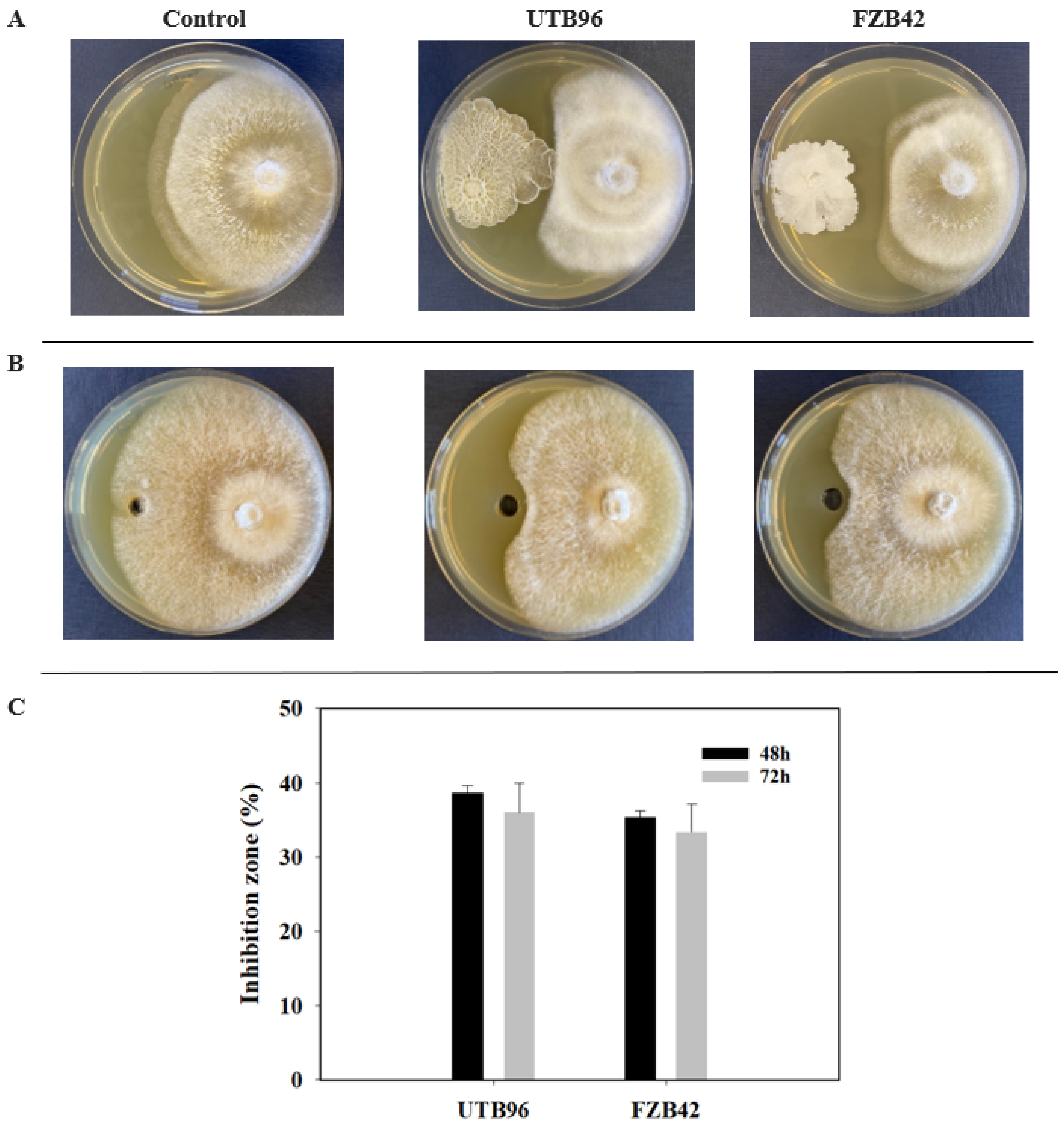
3.4. Antifungal Activity
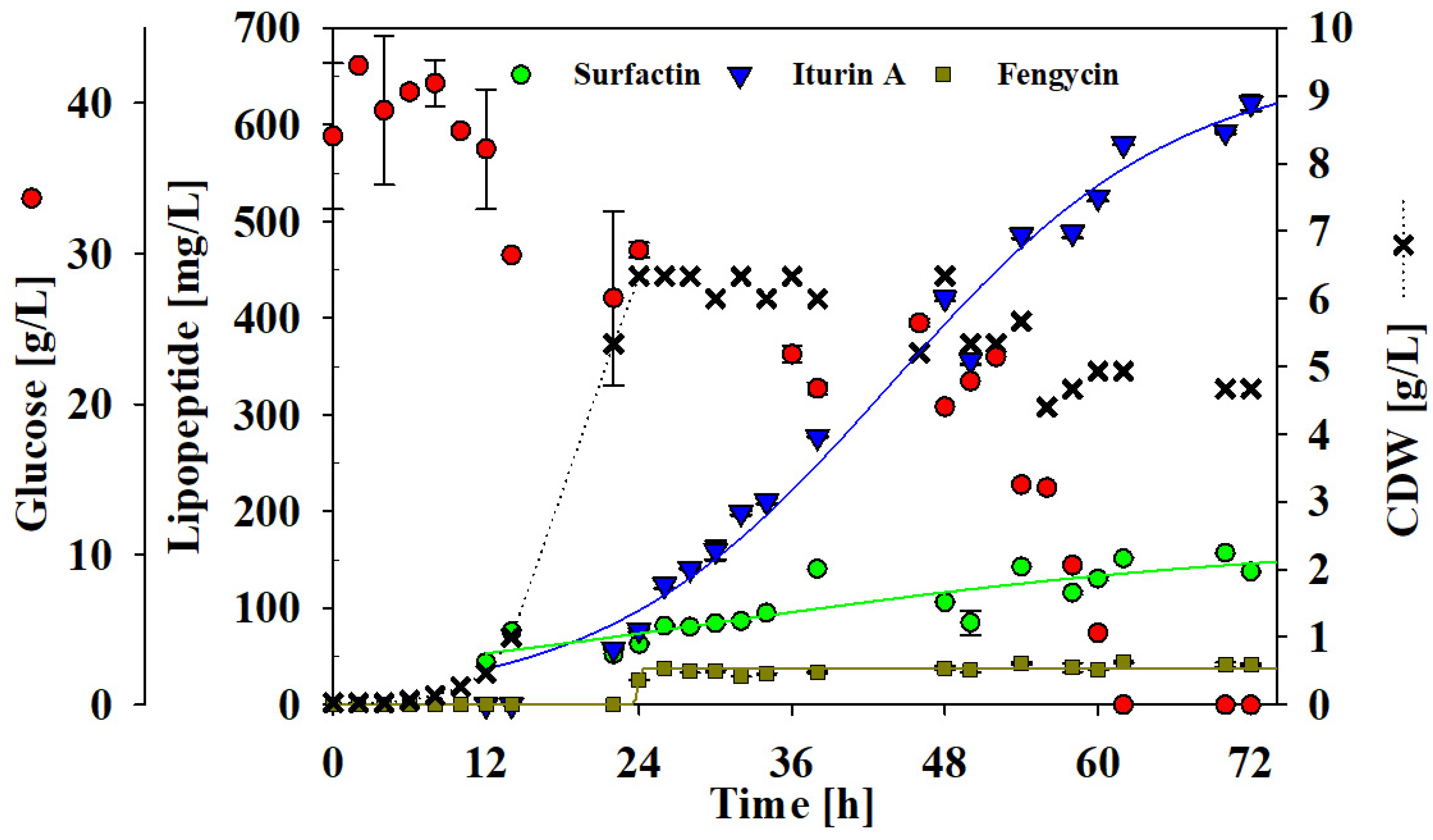
3.5. Batch Bioreactor Cultivation
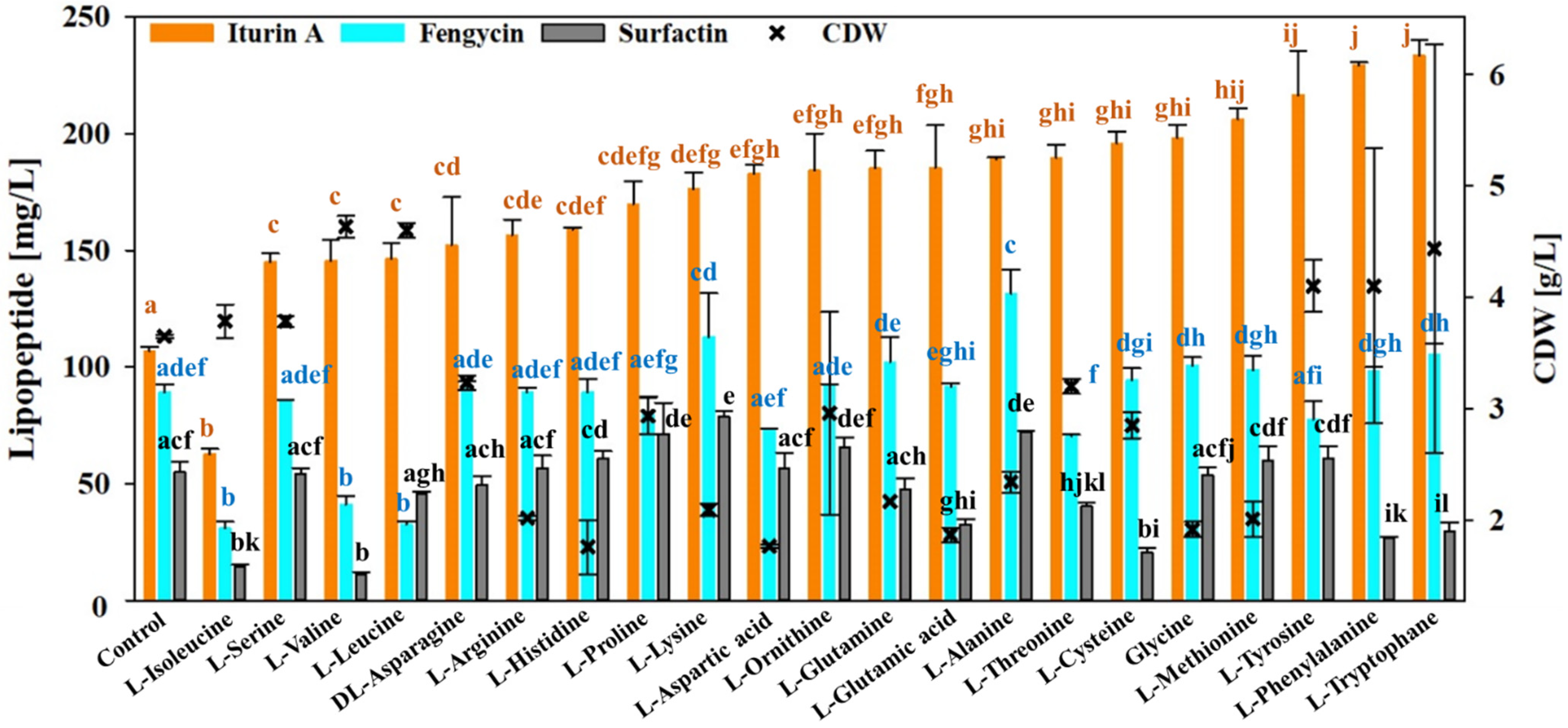
3.6. Effect of Amino Acid Availability on the Lipopeptide Production of B. velezensis UTB96
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rabbee, M.F.; Ali, M.S.; Choi, J.; Hwang, B.S.; Jeong, S.C.; Baek, K.-h. Bacillus velezensis: A valuable member of bioactive molecules within plant microbiomes. Molecules 2019, 24, 1046. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Tang, X.; Yang, R.; Zhang, H.; Li, F.; Tao, F.; Li, F.; Wang, Z. Characteristics and application of a novel species of Bacillus: Bacillus velezensis. ACS Chem. Biol. 2018, 13, 500–505. [Google Scholar] [CrossRef]
- Wu, G.; Liu, Y.; Xu, Y.; Zhang, G.; Shen, Q.; Zhang, R. Exploring elicitors of the beneficial rhizobacterium Bacillus amyloliquefaciens SQR9 to induce plant systemic resistance and their interactions with plant signaling pathways. Mol. Plant-Microbe Interact. 2018, 31, 560–567. [Google Scholar] [CrossRef]
- Borriss, R.; Wu, H.; Gao, X. Secondary metabolites of the plant growth promoting model rhizobacterium Bacillus velezensis FZB42 are involved in direct suppression of plant pathogens and in stimulation of plant-induced systemic resistance. In Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms; Springer: Berlin/Heidelberg, Germany, 2019; pp. 147–168. [Google Scholar]
- Wu, L.; Wu, H.-J.; Qiao, J.; Gao, X.; Borriss, R. Novel routes for improving biocontrol activity of Bacillus based bioinoculants. Front. Microbiol. 2015, 6, 1395. [Google Scholar] [CrossRef]
- Chen, L.; Wang, X.; Ma, Q.; Bian, L.; Liu, X.; Xu, Y.; Zhang, H.; Shao, J.; Liu, Y. Bacillus velezensis CLA178-induced systemic resistance of Rosa multiflora against crown gall disease. Front. Microbiol. 2020, 11, 587667. [Google Scholar] [CrossRef] [PubMed]
- Cawoy, H.; Debois, D.; Franzil, L.; de Pauw, E.; Thonart, P.; Ongena, M. Lipopeptides as main ingredients for inhibition of fungal phytopathogens by Bacillus subtilis/amyloliquefaciens. Microb. Biotechnol. 2015, 8, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef]
- Théatre, A.; Cano-Prieto, C.; Bartolini, M.; Laurin, Y.; Deleu, M.; Niehren, J.; Fida, T.; Gerbinet, S.; Alanjary, M.; Medema, M.H.; et al. The surfactin-like lipopeptides from Bacillus spp.: Natural biodiversity and synthetic biology for a broader application range. Front. Bioeng. Biotechnol. 2021, 9, 623701. [Google Scholar] [CrossRef]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the antimicrobial compounds produced by members of the Bacillus subtilis group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Finking, R.; Marahiel, M.A. Biosynthesis of nonribosomal peptides. Annu. Rev. Microbiol. 2004, 58, 453. [Google Scholar] [CrossRef] [PubMed]
- Geissler, M.; Heravi, K.M.; Henkel, M.; Hausmann, R. Lipopeptide biosurfactants from Bacillus species. In Biobased Surfactants; Elsevier: Amsterdam, The Netherlands, 2019; pp. 205–240. [Google Scholar]
- Dunlap, C.A.; Bowman, M.J.; Rooney, A.P. Iturinic lipopeptide diversity in the Bacillus subtilis species group–important antifungals for plant disease biocontrol applications. Front. Microbiol. 2019, 10, 1794. [Google Scholar] [CrossRef] [PubMed]
- Maget-Dana, R.; Peypoux, F. Iturins, a special class of pore-forming lipopeptides: Biological and physicochemical properties. Toxicology 1994, 87, 151–174. [Google Scholar] [CrossRef]
- Cochrane, S.A.; Vederas, J.C. Lipopeptides from Bacillus and Paenibacillus spp.: A gold mine of antibiotic candidates. Med. Res. Rev. 2016, 36, 4–31. [Google Scholar] [CrossRef] [PubMed]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Dussert, E.; Tourret, M.; Dupuis, C.; Noblecourt, A.; Behra-Miellet, J.; Flahaut, C.; Ravallec, R.; Coutte, F. Evaluation of antiradical and antioxidant activities of lipopeptides produced by Bacillus subtilis strains. Front. Microbiol 2022, 13, 914713. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42–a review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef]
- Jiang, J.; Zhang, H.; Zhang, C.; Han, M.; Du, J.; Yang, X.; Li, W. Production, purification and characterization of ‘Iturin A-2’a lipopeptide with antitumor activity from Chinese Sauerkraut Bacterium Bacillus velezensis T701. Int. J. Pept. Res. Ther. 2021, 27, 2135–2147. [Google Scholar] [CrossRef]
- Lilge, L.; Ersig, N.; Hubel, P.; Aschern, M.; Pillai, E.; Klausmann, P.; Pfannstiel, J.; Henkel, M.; Morabbi Heravi, K.; Hausmann, R. Surfactin shows relatively low antimicrobial activity against Bacillus subtilis and other bacterial model organisms in the absence of synergistic metabolites. Microorganisms 2022, 10, 779. [Google Scholar] [CrossRef]
- Zhao, H.; Shao, D.; Jiang, C.; Shi, J.; Li, Q.; Huang, Q.; Rajoka, M.S.R.; Yang, H.; Jin, M. Biological activity of lipopeptides from Bacillus. Appl. Microbiol. Biotechnol. 2017, 101, 5951–5960. [Google Scholar] [CrossRef] [PubMed]
- Desai, J.D.; Banat, I.M. Microbial production of surfactants and their commercial potential. Microbiol. Mol. Biol. Rev. 1997, 61, 47–64. [Google Scholar] [PubMed]
- Koumoutsi, A.; Chen, X.H.; Henne, A.; Liesegang, H.; Hitzeroth, G.; Franke, P.; Vater, J.; Borriss, R. Structural and functional characterization of gene clusters directing nonribosomal synthesis of bioactive cyclic lipopeptides in Bacillus amyloliquefaciens strain FZB42 2004. Am. Soc. Microbiol. 2004, 186, 1084–1096. [Google Scholar]
- Moreno-Velandia, C.A.; Ongena, M.; Kloepper, J.W.; Cotes, A.M. Biosynthesis of cyclic lipopeptides by Bacillus velezensis Bs006 and its antagonistic activity are modulated by the temperature and culture media conditions. Curr. Microbiol. 2021, 78, 3505–3515. [Google Scholar] [CrossRef]
- Kaspar, F.; Neubauer, P.; Gimpel, M. Bioactive secondary metabolites from Bacillus subtilis: A comprehensive review. J. Nat. Prod. 2019, 82, 2038–2053. [Google Scholar] [CrossRef]
- Leconte, A.; Tournant, L.; Muchembled, J.; Paucellier, J.; Héquet, A.; Deracinois, B.; Deweer, C.; Krier, F.; Deleu, M.; Oste, S.; et al. Assessment of Lipopeptide Mixtures Produced by Bacillus subtilis as biocontrol products against apple scab (Venturia inaequalis). Microorganisms 2022, 10, 1810. [Google Scholar] [CrossRef]
- Qian, S.; Sun, J.; Lu, H.; Lu, F.; Bie, X.; Lu, Z. L-glutamine efficiently stimulates biosynthesis of bacillomycin D in Bacillus subtilis fmbJ. Process Biochem. 2017, 58, 224–229. [Google Scholar] [CrossRef]
- Wu, J.-Y.; Liao, J.-H.; Shieh, C.-J.; Hsieh, F.-C.; Liu, Y.-C. Kinetic analysis on precursors for iturin A production from Bacillus amyloliquefaciens BPD1. J. Biosci. Bioeng. 2018, 126, 630–635. [Google Scholar] [CrossRef]
- Yaseen, Y.; Gancel, F.; Béchet, M.; Drider, D.; Jacques, P. Study of the correlation between fengycin promoter expression and its production by Bacillus subtilis under different culture conditions and the impact on surfactin production. Arch. Microbiol. 2017, 199, 1371–1382. [Google Scholar] [CrossRef]
- Besson, F.; Hourdou, M.L. Effect of amino acids on the biosynthesis of β-amino acids, constituents of bacillomycins F. J. Antibiot. 1987, 40, 221–223. [Google Scholar] [CrossRef]
- Vahidinasab, M.; Ahmadzadeh, M.; Henkel, M.; Hausmann, R.; Morabbi Heravi, K. Bacillus velezensis UTB96 is an antifungal soil isolate with a reduced genome size compared to that of Bacillus velezensis FZB42. Microbiol. Resour. Announc. 2019, 8, e00667-19. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, N.; Ahmadzadeh, M.; Mariotte, P.; Jouzani, G.S. Behavior and interactions of the plant growth-promoting bacteria Azospirillum oryzae NBT506 and Bacillus velezensis UTB96 in a co-culture system. World J. Microbiol. Biotechnol. 2022, 38, 101. [Google Scholar] [CrossRef] [PubMed]
- Afsharmanesh, H.; Ahmadzadeh, M. The Iturin lipopeptides as key compounds in antagonism of Bacillus subtilis UTB96 toward Aspergillus flavus. Biol. Control Pests Plant Dis. 2016, 5, 79–95. [Google Scholar]
- Fan, B.; Wang, C.; Song, X.; Ding, X.; Wu, L.; Wu, H.; Gao, X.; Borriss, R. Bacillus velezensis FZB42 in 2018: The gram-positive model strain for plant growth promotion and biocontrol. Front. Microbiol. 2018, 9, 2491. [Google Scholar] [CrossRef] [PubMed]
- Krebs, B.; Höding, B.; Kübart, S.; Workie, M.A.; Junge, H.; Schmiedeknecht, G.; Grosch, R.; Bochow, H.; Hevesi, M. Use of Bacillus subtilis as biocontrol agent. I. Activities and characterization of Bacillus subtilis strains. J. Plant Dis. Prot. 1998, 105, 181–197. [Google Scholar]
- Santos, J.M.; Vrandečić, K.; Ćosić, J.; Duvnjak, T.; Phillips, A.J.L. Resolving the Diaporthe species occurring on soybean in Croatia. Pers. -Mol. Phylogeny Evol. Fungi 2011, 27, 9–19. [Google Scholar] [CrossRef]
- Hosseini, B.; El-Hasan, A.; Link, T.; Voegele, R.T. Analysis of the species spectrum of the Diaporthe/Phomopsis complex in European soybean seeds. Mycol. Prog. 2020, 19, 455–469. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Bóka, B.; Manczinger, L.; Kecskeméti, A.; Chandrasekaran, M.; Kadaikunnan, S.; Alharbi, N.S.; Vágvölgyi, C.; Szekeres, A. Ion trap mass spectrometry of surfactins produced by Bacillus subtilis SZMC 6179J reveals novel fragmentation features of cyclic lipopeptides. Rapid Commun. Mass Spectrom. 2016, 30, 1581–1590. [Google Scholar] [CrossRef]
- Kecskeméti, A.; Bartal, A.; Bóka, B.; Kredics, L.; Manczinger, L.; Shine, K.; Alharby, N.S.; Khaled, J.M.; Varga, M.; Vágvölgyi, C.; et al. High-frequency occurrence of surfactin monomethyl isoforms in the ferment broth of a Bacillus subtilis strain revealed by ion trap mass spectrometry. Molecules 2018, 23, 2224. [Google Scholar] [CrossRef]
- Lin, L.Z.; Zheng, Q.W.; Wei, T.; Zhang, Z.Q.; Zhao, C.F.; Zhong, H.; Xu, Q.Y.; Lin, J.F.; Guo, L.Q. Isolation and characterization of fengycins produced by Bacillus amyloliquefaciens JFL21 and its broad-spectrum antimicrobial potential against multidrug-resistant foodborne pathogens. Front. Microbiol. 2020, 11, 579621. [Google Scholar] [CrossRef] [PubMed]
- Pathak, K.V.; Keharia, H.; Gupta, K.; Thakur, S.S.; Balaram, P. Lipopeptides from the banyan endophyte, Bacillus subtilis K1: Mass spectrometric characterization of a library of fengycins. J. Am. Soc. Mass Spectrom. 2012, 23, 1716–1728. [Google Scholar] [CrossRef] [PubMed]
- Vahidinasab, M.; Lilge, L.; Reinfurt, A.; Pfannstiel, J.; Henkel, M.; Morabbi Heravi, K.; Hausmann, R. Construction and description of a constitutive plipastatin mono-producing Bacillus subtilis. Microb. Cell Factories 2020, 19, 205. [Google Scholar] [CrossRef] [PubMed]
- Willenbacher, J.; Rau, J.-T.; Rogalla, J.; Syldatk, C.; Hausmann, R. Foam-free production of Surfactin via anaerobic fermentation of Bacillus subtilis DSM 10T. Amb Express 2015, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Klausmann, P.; Hennemann, K.; Hoffmann, M.; Treinen, C.; Aschern, M.; Lilge, L.; Morabbi Heravi, K.; Henkel, M.; Hausmann, R. Bacillus subtilis high cell density fermentation using a sporulation-deficient strain for the production of surfactin. Appl. Microbiol. Biotechnol. 2021, 105, 4141–4151. [Google Scholar] [CrossRef]
- Yazgan, A.; Özcengiz, G.; Marahiel, M.A. Tn10 insertional mutations of Bacillus subtilis that block the biosynthesis of bacilysin. Biochim. Et Biophys. Acta (BBA)-Gene Struct. Expr. 2001, 1518, 87–94. [Google Scholar] [CrossRef]
- Geissler, M.; Oellig, C.; Moss, K.; Schwack, W.; Henkel, M.; Hausmann, R. High-performance thin-layer chromatography (HPTLC) for the simultaneous quantification of the cyclic lipopeptides Surfactin, Iturin A and Fengycin in culture samples of Bacillus species. J. Chromatogr. B 2017, 1044, 214–224. [Google Scholar] [CrossRef]
- Johnson, L.F.; Curl, E.A. Methods for research on the ecology of soil-borne plant pathogens. In Methods for Research on the Ecology of Soil-Borne Plant Pathogens; Burgess Publishing Company: Minneapolis, MN, USA, 1972. [Google Scholar]
- Alenezi, F.N.; Slama, H.B.; Bouket, A.C.; Cherif-Silini, H.; Silini, A.; Luptakova, L.; Nowakowska, J.A.; Oszako, T.; Belbahri, L. Bacillus velezensis: A treasure house of bioactive compounds of medicinal, biocontrol and environmental importance. Forests 2021, 12, 1714. [Google Scholar] [CrossRef]
- Li, X.; Munir, S.; Xu, Y.; Wang, Y.; He, Y. Combined mass spectrometry-guided genome mining and virtual screening for acaricidal activity in secondary metabolites of Bacillus velezensis W1. RSC Adv. 2021, 11, 25441–25449. [Google Scholar] [CrossRef]
- Pupin, M.; Flissi, A.; Jacques, P.; Leclère, V. Bioinformatics tools for the discovery of new lipopeptides with biocontrol applications. Eur. J. Plant Pathol. 2018, 152, 993–1001. [Google Scholar] [CrossRef]
- Xiao, J.; Guo, X.; Qiao, X.; Zhang, X.; Chen, X.; Zhang, D. Activity of fengycin and iturin A isolated from Bacillus subtilis Z-14 on Gaeumannomyces graminis var. tritici and soil microbial diversity. Front. Microbiol. 2021, 1602. [Google Scholar] [CrossRef]
- Cao, Y.; Pi, H.; Chandrangsu, P.; Li, Y.; Wang, Y.; Zhou, H.; Xiong, H.; Helmann, J.D.; Cai, Y. Antagonism of two plant-growth promoting Bacillus velezensis isolates against Ralstonia solanacearum and Fusarium oxysporum. Sci. Rep. 2018, 8, 4360. [Google Scholar] [CrossRef]
- Sawant, S.S.; Song, J.; Seo, H.-J. Characterization of Bacillus velezensis RDA1 as a Biological Control Agent against White Root Rot Disease Caused by Rosellinia necatrix. Plants 2022, 11, 2486. [Google Scholar] [CrossRef]
- Anith, K.N.; Nysanth, N.S.; Natarajan, C. Novel and rapid agar plate methods for in vitro assessment of bacterial biocontrol isolates’ antagonism against multiple fungal phytopathogens. Lett. Appl. Microbiol. 2021, 73, 229–236. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Uhl, J.; Grosch, R.; Alquéres, S.; Pittroff, S.; Dietel, K.; Schmitt-Kopplin, P.; Borriss, R.; Hartmann, A. Cyclic lipopeptides of Bacillus amyloliquefaciens subsp. plantarum colonizing the lettuce rhizosphere enhance plant defense responses toward the bottom rot pathogen Rhizoctonia solani. Mol. Plant-Microbe Interact. 2015, 28, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Calvo, H.; Mendiara, I.; Arias, E.; Blanco, D.; Venturini, M.E. The role of iturin A from B. amyloliquefaciens BUZ-14 in the inhibition of the most common postharvest fruit rots. Food Microbiol. 2019, 82, 62–69. [Google Scholar] [CrossRef]
- Kim, Y.S.; Lee, Y.; Cheon, W.; Park, J.; Kwon, H.T.; Balaraju, K.; Kim, J.; Yoon, Y.J.; Jeon, Y. Characterization of Bacillus velezensis AK-0 as a biocontrol agent against apple bitter rot caused by Colletotrichum gloeosporioides. Sci. Rep. 2021, 11, 626. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Jiang, Q.; Zhao, L.; Su, C.; Li, S.; Si, F.; Li, S.; Zhou, C.; Mu, Y.; Xiao, M. Complete genome sequence of Bacillus velezensis S3-1, a potential biological pesticide with plant pathogen inhibiting and plant promoting capabilities. J. Biotechnol. 2017, 259, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Yang, Y.; Yuan, Q.; Shi, G.; Wu, L.; Lou, Z.; Huo, R.; Wu, H.; Borriss, R.; Gao, X. Bacillomycin D produced by Bacillus amyloliquefaciens is involved in the antagonistic interaction with the plant-pathogenic fungus Fusarium graminearum. Appl. Environ. Microbiol. 2017, 83, e01075-17. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.H.; Vater, J.; Piel, J.; Franke, P.; Scholz, R.; Schneider, K.; Koumoutsi, A.; Hitzeroth, G.; Grammel, N.; Strittmatter, A.W.; et al. Structural and functional characterization of three polyketide synthase gene clusters in Bacillus amyloliquefaciens FZB42. J. Bacteriol. 2006, 188, 4024–4036. [Google Scholar] [CrossRef]
- Yokota, K.; Hayakawa, H. Impact of antimicrobial lipopeptides from Bacillus sp. on suppression of Fusarium yellows of tatsoi. Microbes Environ. 2015, ME15062. [Google Scholar]
- Jin, H.; Li, K.; Niu, Y.; Guo, M.; Hu, C.; Chen, S.; Huang, F. Continuous enhancement of iturin A production by Bacillus subtilis with a stepwise two-stage glucose feeding strategy. BMC Biotechnol. 2015, 15, 53. [Google Scholar] [CrossRef] [PubMed]
- Belitsky, B.R. Role of branched-chain amino acid transport in Bacillus subtilis CodY activity. J. Bacteriol. 2015, 197, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Serror, P.; Sonenshein, A.L. CodY is required for nutritional repression of Bacillus subtilis genetic competence. J. Bacteriol. 1996, 178, 5910–5915. [Google Scholar] [CrossRef]
- Sun, J.; Liu, Y.; Lin, F.; Lu, Z.; Lu, Y. CodY, ComA, DegU and Spo0A controlling lipopeptides biosynthesis in Bacillus amyloliquefaciens fmbJ. J. Appl. Microbiol. 2021, 131, 1289–1304. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Similarity with the Reference Strain Bacillus velezensis FZB42 | |||
|---|---|---|---|
| Lipopeptide | Chromosomal Localization (from–to) | Similarity in Nucleotide Sequence | Similarity in Amino Acid Sequence |
| Surfactin | 301,941 nt–366,339 nt | 97.53% | 91.67% |
| Iturin A | 1,762,934 nt–1,800,180 nt | 97.02% | 67.94% |
| Fengycin | 1,823,096 nt–1,860,765 nt | 96.50% | 97.08% |
| Surfactin | Iturin A in UTB96 or Bacillomycin D in FZB42 | Fengycin | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B. velezensis | Initial Glucose [g/L] | YX/S [g/g] | Growth Rate µ [1/h] | YP/S [mg/g] | YP/X [mg/g] | q [mg/g.h] | YP/S [mg/g] | YP/X [mg/g] | q [mg/g.h] | YP/S [mg/g] | YP/X [mg/g] | q [mg/g.h] |
| UTB96 | 8 (SF) | 0.2 ± 0.01 | 0.2 ± 0.0 | 12.9 ± 1.16 | 27.4 ± 1.59 | 2.3 ± 0.13 | 5.8 ± 0.25 | 168.3 ± 8.76 | 1.2 ± 0.06 | 9.6 ± 0.13 | 281.5 ± 6.35 | 2.0 ± 0.04 |
| 20 (SF) | 0.6 ± 0.02 | 0.1 ± 0.0 | 3.6 ± 0.08 | 46.3 ± 5.93 | 0.6 ± 0.07 | 7.0 ± 0.86 | 134.4 ± 10.33 | 1.2 ± 0.09 | 3.4 ± 0.08 | 82.6 ± 1.41 | 0.7 ± 0.01 | |
| 40 (SF) | 0.1 ± 0.01 | 0.1 ± 0.0 | 1.7 ± 0.1 | 62.4 ± 4.08 | 1.4 ± 0.09 | 3.5 ± 0.30 | 81.9 ± 9.51 | 0.6 ± 0.06 | 2.5 ± 0.13 | 55.4 ± 1.57 | 0,7 ± 0.01 | |
| 40 (BR) | 0.6 ± 0.0 | 0.2 ± 0.0 | 8.4 ± 0.2 | 43.8 ± 3.8 | 0.9 ± 0.1 | 16.3 ± 0.8 | 249.4 ± 15.3 | 1.8 ± 0.0 | 1.2 ± 0.0 | 17.4 ±0.3 | 0.1 ± 0.0 | |
| FZB42 | 8 (SF) | 0,2 ± 0.021 | 0.2 ± 0.00 | 5.0 ±0.24 | 120.6 ± 1.75 | 1.8 ± 0.00 | 1.4 ± 0 | 12.0 ± 2.04 | 0.3 ± 0.04 | 3.7 ± 0.59 | 81.9 ± 8.09 | 0.9 ±0.03 |
| 20 (SF) | 0.2 0.01 | 0.2 ± 0.01 | 1.2 ± 0.11 | 21.7 ± 1.68 | 0.3 ± 0.00 | 0.9 ± 0.01 | 35.2 ± 3.11 | 0.2 ± 0.02 | 1.3 ± 0.06 | 54.4 ± 6.23 | 0,3 ± 0.04 | |
| 40 (SF) | 0.1 ± 0.0 | 0.1 ± 0.0 | 2.1 ± 0.04 | 39.6 ± 5.01 | 0.5 ± 0.06 | 0.4 ± 0.00 | 7.5 ± 0.53 | 0.1 ± 0.0 | 1.9 ± 0.4 | 32.1 ± 9.63 | 0.5 ± 0.14 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vahidinasab, M.; Adiek, I.; Hosseini, B.; Akintayo, S.O.; Abrishamchi, B.; Pfannstiel, J.; Henkel, M.; Lilge, L.; Voegele, R.T.; Hausmann, R. Characterization of Bacillus velezensis UTB96, Demonstrating Improved Lipopeptide Production Compared to the Strain B. velezensis FZB42. Microorganisms 2022, 10, 2225. https://doi.org/10.3390/microorganisms10112225
Vahidinasab M, Adiek I, Hosseini B, Akintayo SO, Abrishamchi B, Pfannstiel J, Henkel M, Lilge L, Voegele RT, Hausmann R. Characterization of Bacillus velezensis UTB96, Demonstrating Improved Lipopeptide Production Compared to the Strain B. velezensis FZB42. Microorganisms. 2022; 10(11):2225. https://doi.org/10.3390/microorganisms10112225
Chicago/Turabian StyleVahidinasab, Maliheh, Isabel Adiek, Behnoush Hosseini, Stephen Olusanmi Akintayo, Bahar Abrishamchi, Jens Pfannstiel, Marius Henkel, Lars Lilge, Ralf T. Voegele, and Rudolf Hausmann. 2022. "Characterization of Bacillus velezensis UTB96, Demonstrating Improved Lipopeptide Production Compared to the Strain B. velezensis FZB42" Microorganisms 10, no. 11: 2225. https://doi.org/10.3390/microorganisms10112225
APA StyleVahidinasab, M., Adiek, I., Hosseini, B., Akintayo, S. O., Abrishamchi, B., Pfannstiel, J., Henkel, M., Lilge, L., Voegele, R. T., & Hausmann, R. (2022). Characterization of Bacillus velezensis UTB96, Demonstrating Improved Lipopeptide Production Compared to the Strain B. velezensis FZB42. Microorganisms, 10(11), 2225. https://doi.org/10.3390/microorganisms10112225