
Dynamics of the Queensland Fruit Fly Microbiome through the Transition from Nature to an Established Laboratory Colony



Abstract
:1. Introduction
2. Materials and Methods
2.1. Colony Origins
2.2. Colony Maintenance
2.3. Sample Preparation
2.4. Microbiome Profiling
2.5. Sequence Data Processing
2.6. Microbiome Analysis
3. Results
3.1. 16s rRNA Sequence Reads and OTUs
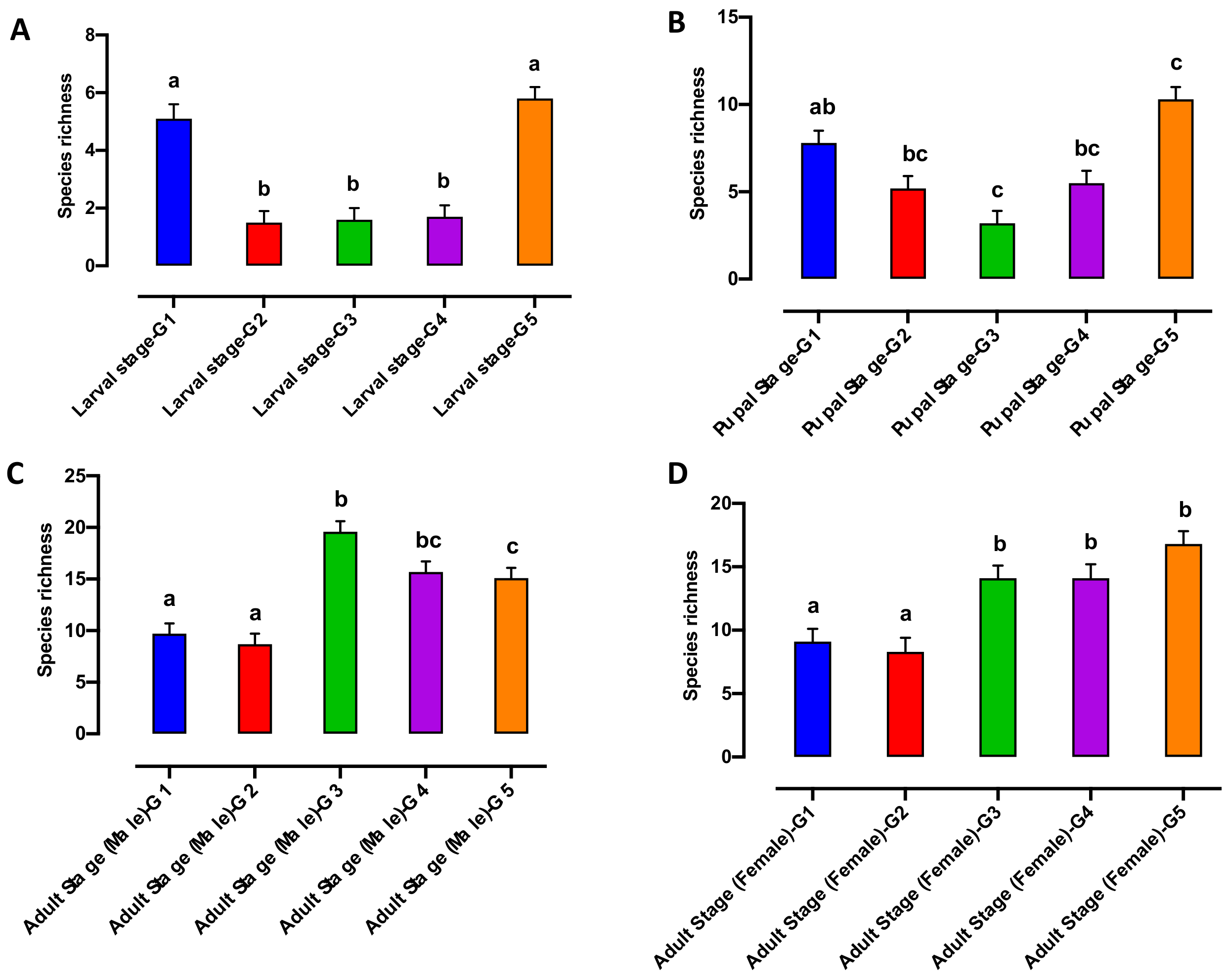
3.2. Bacterial Alpha and Beta Diversity during Qfly Domestication
3.3. Bacterial Communities Associated with Domestication of the Qfly
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Enkerlin, W. Impact of fruit fly control programmes using the sterile insect technique. In Sterile Insect Technique; Springer: Berlin/Heidelberg, Germany, 2005; pp. 651–676. [Google Scholar]
- Hendrichs, J.; Franz, G.; Rendon, P. Increased effectiveness and applicability of the sterile insect technique through male-only releases for control of Mediterranean fruit flies during fruiting seasons. J. Appl. Èntomol. 1995, 119, 371–377. [Google Scholar] [CrossRef]
- Knipling, E.F. Possibilities of Insect Control or Eradication Through the Use of Sexually Sterile Males1. J. Econ. Èntomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Vreysen, M.J.B.; Barclay, H.J.; Hendrichs, J. Modeling of Preferential Mating in Areawide Control Programs That Integrate the Release of Strains of Sterile Males Only or Both Sexes. Ann. Èntomol. Soc. Am. 2006, 99, 607–616. [Google Scholar] [CrossRef]
- Chambers, D.L. Quality control in mass rearing. Annu. Rev. Entomol. 1977, 22, 289–308. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Hallas, R.; Sinclair, C.; Partridge, L. Rapid loss of stress resistance in Drosophila melanogaster under adaptation to laboratory culture. Evolution 2001, 55, 436–438. [Google Scholar] [CrossRef]
- Hoffmann, A.; Ross, P. Rates and Patterns of Laboratory Adaptation in (Mostly) Insects. J. Econ. Èntomol. 2018, 111, 501–509. [Google Scholar] [CrossRef]
- Ochieng’-Odero, J. Does adaptation occur in insect rearing systems, or is it a case of selection, acclimatization and domestication? Int. J. Trop. Insect Sci. 1994, 15, 1–7. [Google Scholar] [CrossRef]
- Pérez, J.; Mendez, V.; Yuval, B.; Taylor, P.W. Domestication-related changes in sexual performance of Queensland fruit fly. Insect Sci. 2020, 28, 1491–1503. [Google Scholar] [CrossRef]
- Gilchrist, A.S.; Cameron, E.C.; Sved, J.A.; Meats, A.W. Genetic Consequences of Domestication and Mass Rearing of Pest Fruit Fly Bactrocera tryoni (Diptera: Tephritidae). J. Econ. Èntomol. 2012, 105, 1051–1056. [Google Scholar] [CrossRef]
- Majumder, R.; Sutcliffe, B.; Adnan, S.M.; Mainali, B.; Dominiak, B.C.; Taylor, P.W.; Chapman, T.A. Artificial Larval Diet Mediates the Microbiome of Queensland Fruit Fly. Front. Microbiol. 2020, 11, 576156. [Google Scholar] [CrossRef]
- Pérez, J.; Park, S.J.; Taylor, P.W. Domestication modifies the volatile emissions produced by male Queensland fruit flies during sexual advertisement. Sci. Rep. 2018, 8, 16503. [Google Scholar] [CrossRef]
- Schutze, M.K.; Dammalage, T.; Jessup, A.; Vreysen, M.J.; Wornoayporn, V.; Clarke, A. Effects of laboratory colonization on Bactrocera dorsalis (Diptera, Tephritidae) mating behaviour: ‘what a difference a year makes’. ZooKeys 2015, 540, 369–383. [Google Scholar] [CrossRef]
- Zygouridis, N.E.; Argov, Y.; Nemnylavy, E.; Augustinos, A.A.; Nestel, D.; Mathiopoulos, K.D. Genetic changes during laboratory domestication of an olive fly SIT strain. J. Appl. Èntomol. 2013, 138, 423–432. [Google Scholar] [CrossRef]
- Jurkevitch, E. Riding the Trojan horse: Combating pest insects with their own symbionts. Microb. Biotechnol. 2011, 4, 620–627. [Google Scholar] [CrossRef] [Green Version]
- Dillon, R.J.; Dillon, V.M. The Gut Bacteria of Insects: Nonpathogenic Interactions. Annu. Rev. Èntomol. 2004, 49, 71–92. [Google Scholar] [CrossRef]
- Broderick, N.; Raffa, K.F.; Goodman, R.M.; Handelsman, J. Census of the Bacterial Community of the Gypsy Moth Larval Midgut by Using Culturing and Culture-Independent Methods. Appl. Environ. Microbiol. 2004, 70, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Robinson, C.J.; Schloss, P.; Ramos, Y.; Raffa, K.; Handelsman, J. Robustness of the Bacterial Community in the Cabbage White Butterfly Larval Midgut. Microb. Ecol. 2010, 59, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Akami, M.; Ren, X.-M.; Qi, X.; Mansour, A.; Gao, B.; Cao, S.; Niu, C.-Y. Symbiotic bacteria motivate the foraging decision and promote fecundity and survival of Bactrocera dorsalis (Diptera: Tephritidae). BMC Microbiol. 2019, 19, 229. [Google Scholar] [CrossRef] [Green Version]
- Baumann, P. Biology of bacteriocyte-associated endosymbionts of plant sap-sucking insects. Annu. Rev. Microbiol. 2005, 59, 155–189. [Google Scholar] [CrossRef]
- Nogge, G. Significance of symbionts for the maintenance of an optimal nutritional state for successful reproduction in hematophagous arthropods. In Parasitology; Cambridge University Press: Cambridge, UK, 1989; pp. 101–104. [Google Scholar]
- Douglas, A.E. Nutritional Interactions in Insect-Microbial Symbioses: Aphids and Their Symbiotic Bacteria Buchnera. Annu. Rev. Èntomol. 1998, 43, 17–37. [Google Scholar] [CrossRef] [Green Version]
- Benemann, J.R. Nitrogen Fixation in Termites. Science 1973, 181, 164–165. [Google Scholar] [CrossRef]
- Ben-Yosef, M.; Behar, A.; Jurkevitch, E.; Yuval, B. Bacteria-diet interactions affect longevity in the medfly-Ceratitis capitata. J. Appl. Èntomol. 2008, 132, 690–694. [Google Scholar] [CrossRef]
- Dinh, H.; Mendez, V.; Tabrizi, S.T.; Ponton, F. Macronutrients and infection in fruit flies. Insect Biochem. Mol. Biol. 2019, 110, 98–104. [Google Scholar] [CrossRef]
- Majumder, R.; Sutcliffe, B.; Taylor, P.W.; Chapman, T.A. Microbiome of the Queensland Fruit Fly through Metamorphosis. Microorganisms 2020, 8, 795. [Google Scholar] [CrossRef]
- Yong, H.-S.; Song, S.-L.; Chua, K.-O.; Lim, P.-E. High Diversity of Bacterial Communities in Developmental Stages of Bactrocera carambolae (Insecta: Tephritidae) Revealed by Illumina MiSeq Sequencing of 16S rRNA Gene. Curr. Microbiol. 2017, 74, 1076–1082. [Google Scholar] [CrossRef]
- Andongma, A.A.; Wan, L.; Dong, Y.-C.; Li, P.; Desneux, N.; White, J.A.; Niu, C.-Y. Pyrosequencing reveals a shift in symbiotic bacteria populations across life stages of Bactrocera dorsalis. Sci. Rep. 2015, 5, srep09470. [Google Scholar] [CrossRef]
- Yong, H.-S.; Song, S.-L.; Chua, K.-O.; Lim, P.-E. Predominance of Wolbachia endosymbiont in the microbiota across life stages of Bactrocera latifrons (Insecta: Tephritidae). Meta Gene 2017, 14, 6–11. [Google Scholar] [CrossRef]
- Andongma, A.A.; Wan, L.; Dong, Y.-C.; Wang, Y.-L.; He, J.; Niu, C.-Y. Assessment of the Bacteria community structure across life stages of the Chinese Citrus Fly, Bactrocera minax (Diptera: Tephritidae). BMC Microbiol. 2019, 19, 285. [Google Scholar] [CrossRef] [Green Version]
- Malacrinò, A.; Campolo, O.; Medina, R.; Palmeri, V. Instar- and host-associated differentiation of bacterial communities in the Mediterranean fruit fly Ceratitis capitata. PLoS ONE 2018, 13, e0194131. [Google Scholar] [CrossRef]
- Ventura, C.; Briones-Roblero, C.I.; Hernandez, E.; Rivera-Orduña, F.N.; Zúñiga, G. Comparative Analysis of the Gut Bacterial Community of Four Anastrepha Fruit Flies (Diptera: Tephritidae) Based on Pyrosequencing. Curr. Microbiol. 2018, 75, 966–976. [Google Scholar] [CrossRef]
- Noman, S.; Shi, G.; Liu, L.; Li, Z. Diversity of bacteria in different life stages and their impact on the development and reproduction of Zeugodacus tau (Diptera: Tephritidae). Insect Sci. 2020, 28, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.R.; Powell, K.S.; Weldon, C.W.; Taylor, P.W. The ecology of Bactrocera tryoni (Diptera: Tephritidae): What do we know to assist pest management? Ann. Appl. Biol. 2011, 158, 26–54. [Google Scholar] [CrossRef] [Green Version]
- Dominiak, B.; Westcott, A.; Barchia, I. Release of sterile Queensland fruit fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae), at Sydney, Australia. Aust. J. Exp. Agric. 2003, 43, 519–528. [Google Scholar] [CrossRef]
- Fanson, B.; Sundaralingam, S.; Jiang, L.; Dominiak, B.; D’Arcy, G. A review of 16 years of quality control parameters at a mass-rearing facility producing Queensland fruit fly, Bactrocera tryoni. EÈntomol. Exp. Appl. 2014, 151, 152–159. [Google Scholar] [CrossRef]
- Jessup, A.J.; Dominiak, B.; Woods, B.; De Lima, C.P.F.; Tomkins, A.; Smallridge, C.J. Area-Wide Management of Fruit Flies in Australia. In Area-Wide Control of Insect Pests; Springer: Berlin/Heidelberg, Germany, 2007; pp. 685–697. [Google Scholar] [CrossRef]
- Deutscher, A.T.; Burke, C.M.; Darling, A.E.; Riegler, M.; Reynolds, O.L.; Chapman, T.A. Near full-length 16S rRNA gene next-generation sequencing revealed Asaia as a common midgut bacterium of wild and domesticated Queensland fruit fly larvae. Microbiome 2018, 6, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumder, R.; Sutcliffe, B.; Taylor, P.W.; Chapman, T.A. Next-Generation Sequencing reveals relationship between the larval microbiome and food substrate in the polyphagous Queensland fruit fly. Sci. Rep. 2019, 9, 14292. [Google Scholar] [CrossRef] [Green Version]
- Majumder, R.; Sutcliffe, B.; Taylor, P.W.; Chapman, T.A. Fruit host-dependent fungal communities in the microbiome of wild Queensland fruit fly larvae. Sci. Rep. 2020, 10, 16550. [Google Scholar] [CrossRef]
- Fitt, G.; O’Brien, R.W. Bacteria associated with four species of Dacus (Diptera: Tephritidae) and their role in the nutrition of the larvae. Oecologia 1985, 67, 447–454. [Google Scholar] [CrossRef]
- Morrow, J.L.; Frommer, M.; Shearman, D.C.A.; Riegler, M. The Microbiome of Field-Caught and Laboratory-Adapted Australian Tephritid Fruit Fly Species with Different Host Plant Use and Specialisation. Microb. Ecol. 2015, 70, 498–508. [Google Scholar] [CrossRef]
- Thaochan, N.; Drew, R.A.I.; Hughes, J.M.; Vijaysegaran, S.; Chinajariyawong, A. Alimentary tract bacteria isolated and identified with API-20E and molecular cloning techniques from Australian tropical fruit flies, Bactrocera cacuminata and B. tryoni. J. Insect Sci. 2010, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Woruba, D.N.; Morrow, J.L.; Reynolds, O.L.; Chapman, T.A.; Collins, D.P.; Riegler, M. Diet and irradiation effects on the bacterial community composition and structure in the gut of domesticated teneral and mature Queensland fruit fly, Bactrocera tryoni (Diptera: Tephritidae). BMC Microbiol. 2019, 19, 281. [Google Scholar] [CrossRef] [Green Version]
- Mainali, B.P.; Moadeli, T.; Ponton, F.; Taylor, P.W. Comparison of Gel Larval Diet with Traditional Lucerne Chaff and Carrot Solid Diets for Rearing of Queensland Fruit Fly (Diptera: Tephritidae). J. Econ. Èntomol. 2019, 112, 2278–2286. [Google Scholar] [CrossRef]
- Moadeli, T.; Taylor, P.W.; Ponton, F. High productivity gel diets for rearing of Queensland fruit fly, Bactrocera tryoni. J. Pest Sci. 2016, 90, 507–520. [Google Scholar] [CrossRef]
- Collins, S.R.; Weldon, C.; Banos, C.; Taylor, P. Effects of irradiation dose rate on quality and sterility of Queensland fruit flies, Bactrocera tryoni (Froggatt). J. Appl. Èntomol. 2008, 132, 398–405. [Google Scholar] [CrossRef]
- Lane, D. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Wiley: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Fouts, D.E.; Szpakowski, S.; Purushe, J.; Torralba, M.; Waterman, R.C.; MacNeil, M.D.; Alexander, L.J.; Nelson, K.E. Next Generation Sequencing to Define Prokaryotic and Fungal Diversity in the Bovine Rumen. PLoS ONE 2012, 7, e48289. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, P. Greenfield Hybrid Analysis Pipeline (GHAP), v1; CSIRO: Canberra, Australia, 2017. [Google Scholar]
- Sutcliffe, B.; Chariton, A.; Harford, A.; Hose, G.; Greenfield, P.; Elbourne, L.; Oytam, Y.; Stephenson, S.; Midgley, D.J.; Paulsen, I.T. Effects of uranium concentration on microbial community structure and functional potential. Environ. Microbiol. 2017, 19, 3323–3341. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Maidak, B.L.; Olsen, G.J.; Larsen, N.; Overbeek, R.; McCaughey, M.J.; Woese, C.R. The Ribosomal Database Project (RDP). Nucleic Acids Res. 1996, 24, 82–85. [Google Scholar] [CrossRef] [Green Version]
- Behar, A.; Yuval, B.; Jurkevitch, E. Gut bacterial communities in the Mediterranean fruit fly (Ceratitis capitata) and their impact on host longevity. J. Insect Physiol. 2008, 54, 1377–1383. [Google Scholar] [CrossRef]
- Behar, A.; Yuval, B.; Jurkevitch, E. Enterobacteria-mediated nitrogen fixation in natural populations of the fruit fly Ceratitis capitata. Mol. Ecol. 2005, 14, 2637–2643. [Google Scholar] [CrossRef]
- Khan, M.; Mahin, A.; Pramanik, M.; Akter, H. Identification of Gut Bacterial Community and Their Effect on the Fecundity of Pumpkin Fly, Bactrocera tau (Walker). J. Èntomol. 2014, 11, 68–77. [Google Scholar] [CrossRef]
- Reddy, K.; Sharma, K.; Singh, S. Attractancy potential of culturable bacteria from the gut of peach fruit fly, Bactrocera zonata (Saunders). Phytoparasitica 2014, 42, 691–698. [Google Scholar] [CrossRef]
- Capuzzo, C.; Firrao, G.; Mazzon, L.; Squartini, A.; Girolami, V. ‘Candidatus Erwinia dacicola’, a coevolved symbiotic bacterium of the olive fly Bactrocera oleae (Gmelin). Int. J. Syst. Evol. Microbiol. 2005, 55, 1641–1647. [Google Scholar] [CrossRef]
- Kuzina, L.V.; Peloquin, J.J.; Vacek, D.C.; Miller, T.A. Isolation and Identification of Bacteria Associated with Adult Laboratory Mexican Fruit Flies, Anastrepha ludens (Diptera: Tephritidae). Curr. Microbiol. 2001, 42, 290–294. [Google Scholar] [CrossRef]
- Xiang, H.; Wei, G.-F.; Jia, S.; Huang, J.; Miao, X.-X.; Zhou, Z.; Zhao, L.-P.; Huang, Y.-P. Microbial communities in the larval midgut of laboratory and field populations of cotton bollworm (Helicoverpa armigera). Can. J. Microbiol. 2006, 52, 1085–1092. [Google Scholar] [CrossRef]
- Kudo, R.; Masuya, H.; Endoh, R.; Kikuchi, T.; Ikeda, H. Gut bacterial and fungal communities in ground-dwelling beetles are associated with host food habit and habitat. ISME J. 2018, 13, 676–685. [Google Scholar] [CrossRef]
- Quan, A.S.; Eisen, M.B. The ecology of the Drosophila-yeast mutualism in wineries. PLoS ONE 2018, 13, e0196440. [Google Scholar] [CrossRef] [Green Version]
- Broderick, N.A.; Lemaitre, B. Gut-associated microbes of Drosophila melanogaster. Gut Microbes 2012, 3, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Chandler, J.A.; Lang, J.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial Communities of Diverse Drosophila Species: Ecological Context of a Host-Microbe Model System. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef]
- Mason, C.J.; Couture, J.J.; Raffa, K.F. Plant-associated bacteria degrade defense chemicals and reduce their adverse effects on an insect defoliator. Oecologia 2014, 175, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.-H.; Roh, S.W.; Whon, T.W.; Jung, M.-J.; Kim, M.-S.; Park, D.-S.; Yoon, C.; Nam, Y.-D.; Kim, Y.-J.; Choi, J.-H.; et al. Insect Gut Bacterial Diversity Determined by Environmental Habitat, Diet, Developmental Stage, and Phylogeny of Host. Appl. Environ. Microbiol. 2014, 80, 5254–5264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravenscraft, A.; Berry, M.; Hammer, T.; Peay, K.; Boggs, C. Structure and function of the bacterial and fungal gut microbiota of Neotropical butterflies. Ecol. Monogr. 2019, 89, e01346. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographic Location of Collection | Fruit Source and Number of Fruit Collected | Collection Date |
|---|---|---|
| Coomealla, NSW GPS: Lat 34°5′50.97′′, Long. 142°3′7.21′′ | Pomegranate (Punica granatum) 37 pieces | 5 May 2017 |
| St. Germains, between Tatura and Echuca, Victoria GPS: Lat 36°10′48.86′′, Long. 145°8′50.74′′ | Green Apple (Malus pumila) 41 pieces | 5 May 2017 |
| Downer Road, between Tatura and Toolamba, Victoria GPS: Lat 26°38′34.92′′, Long. 152°56′22.99′′ | Quince (Cydonia oblonga) 52 pieces | 5 May 2017 |
| Ingredients | 1 kg Diet Preparation | Company Name and Catalogue Number |
|---|---|---|
| Brewer’s Yeast (g) | 204 | Lallemand Australia Pty Ltd., Edwardstown, SA, Australia |
| Sugar (g) | 121.8 | MP Biomedicals LLC, France (Cat. no02902978) |
| Agar (g) | 10 | Sigma–Aldrich® St. Louis, MO, USA |
| Citric Acid (g) | 23.1 | Sigma–Aldrich®, St. Louis, MO, USA |
| Nipagin (g) | 2 | Southern Biological, Knoxfield, VIC, Australia (Cat no MC11.2) |
| Sodium benzoate (g) | 2 | Sigma–Aldrich® St. Louis, MO, USA |
| Wheat Germ Oil (mL) | 2 | Melrose Laboratories Pty Ltd., Australia |
| Water (mL) | 1000 | Milli-Q water |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majumder, R.; Taylor, P.W.; Chapman, T.A. Dynamics of the Queensland Fruit Fly Microbiome through the Transition from Nature to an Established Laboratory Colony. Microorganisms 2022, 10, 291. https://doi.org/10.3390/microorganisms10020291
Majumder R, Taylor PW, Chapman TA. Dynamics of the Queensland Fruit Fly Microbiome through the Transition from Nature to an Established Laboratory Colony. Microorganisms. 2022; 10(2):291. https://doi.org/10.3390/microorganisms10020291
Chicago/Turabian StyleMajumder, Rajib, Phillip W. Taylor, and Toni A. Chapman. 2022. "Dynamics of the Queensland Fruit Fly Microbiome through the Transition from Nature to an Established Laboratory Colony" Microorganisms 10, no. 2: 291. https://doi.org/10.3390/microorganisms10020291
APA StyleMajumder, R., Taylor, P. W., & Chapman, T. A. (2022). Dynamics of the Queensland Fruit Fly Microbiome through the Transition from Nature to an Established Laboratory Colony. Microorganisms, 10(2), 291. https://doi.org/10.3390/microorganisms10020291