
Genetic Diversity of Trypanosoma cruzi in Panama Inferred by Multi-locus Sequence Typing of Mitochondrial Genes

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and T. cruzi Genotyping
2.2. Sequence Editing and Bayesian Phylogenetic Inference
2.3. Haplotype Network Construction
2.4. Estimation of Evolutionary Divergency between TcI Mitochondrial Sequences
3. Results
3.1. Phylogenetic Tree and Haplotype Network Results
3.2. Nucleotide Divergence between Panamanian TcI Isolates and Reference Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hotez, P.J.; Damania, A.; Bottazzi, M.E. Central Latin America: Two decades of challenges in neglected tropical disease control. PLoS Neglected Trop. Dis. 2020, 14, e0007962. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J.; Woc-Colburn, L.; Bottazzi, M.E. Neglected tropical diseases in Central America and Panama: Review of their prevalence, populations at risk and impact on regional development. Int. J. Parasitol. 2014, 44, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Dorn, P.L.; McClure, A.G.; Gallaspy, M.D.; Waleckx, E.; Woods, A.S.; Monroy, M.C.; Stevens, L. The diversity of the Chagas parasite, Trypanosoma cruzi, infecting the main Central American vector, Triatoma dimidiata, from Mexico to Colombia. PLoS Neglected Trop. Dis. 2017, 11, e0005878. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.K.; Yoshioka, K.; Hashimoto, K.; Caranci, A.; Gottdenker, N.; Monroy, C.; Saldaña, A.; Rodriguez, S.; Dorn, P.; Zúniga, C. Chagas Disease Epidemiology in Central America: An Update. Curr. Trop. Med. Rep. 2019, 6, 92–105. [Google Scholar] [CrossRef]
- Whitlaw, J.T.; Chaniotis, B.N., Jr. Palm Trees and Chagas’ Disease in Panama. Am. J. Trop. Med. Hyg. 1978, 27, 873–881. [Google Scholar] [CrossRef]
- Vasquez, A.M.; Samudio, F.E.; Saldaña, A.; Paz, H.M.; Calzada, J.E. Eco-epidemiological aspects of Trypanosoma cruzi, Trypanosoma rangeli and their vector (Rhodnius pallescens) in Panama. Rev. Inst. Med. Trop. Sao Paulo. 2004, 46, 217–222. [Google Scholar] [CrossRef]
- Abad-Franch, F.; Lima, M.M.; Sarquis, O.; Gurgel-Gonçalves, R.; Sánchez-Martín, M.; Calzada, J.; Saldaña, A.; Monteiro, F.A.; Palomeque, F.S.; Santos, W.S.; et al. On palms, bugs, and Chagas disease in the Americas. Acta Trop. 2015, 151, 126–141. [Google Scholar] [CrossRef]
- Rodriguez, I.G.; Loaiza, J.R. American trypanosomiasis, or Chagas disease, in Panama: A chronological synopsis of ecological and epidemiological research. Parasites Vectors 2017, 10, 1–16. [Google Scholar] [CrossRef]
- Sousa, O.E.; Johnson, C.M. Frequency and Distribution of Trypanosoma cruzi and Trypanosoma Rangeli in the Republic of Panamá. Am. J. Trop. Med. Hyg. 1971, 20, 405–410. [Google Scholar] [CrossRef]
- Saldaña, A.; Samudio, F.; Miranda, A.; Herrera, L.M.; Saavedra, S.P.; Cáceres, L.; Bayard, V.; Calzada, J.E. Predominance of Trypanosoma rangeli infection in children from a Chagas disease endemic area in the west-shore of the Panama canal. Mem. Inst. Oswaldo Cruz. 2005, 100, 729–731. [Google Scholar] [CrossRef]
- Calzada, J.E.; Pineda, V.; Samudio, F.; Santamaría, A.M.; Cáceres, L.; Montalvo, E.; Bayard, V.; Saldaña, A.; Alvarez, D. Human Trypanosome Infection and the Presence of Intradomicile Rhodnius pallescens in the Western Border of the Panama Canal, Panama. Am. J. Trop. Med. Hyg. 2006, 74, 762–765. [Google Scholar] [CrossRef] [PubMed]
- Saldaña, A.; Pineda, V.; Martinez, I.; Santamaria, G.; Santamaría, A.M.; Miranda, A.; Calzada, J.E. A New Endemic Focus of Chagas Disease in the Northern Region of Veraguas Province, Western Half Panama, Central America. PLoS ONE 2012, 7, e34657. [Google Scholar] [CrossRef] [PubMed]
- Basso, B.; Moretti, E.; Fretes, R. Vaccination with epimastigotes of different strains of Trypanosoma rangeli protects mice against Trypanosoma cruzi infection. Mem. Inst. Oswaldo Cruz. 2008, 103, 370–374. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Marini, V.; Moretti, E.; Bermejo, D.; Basso, B. Vaccination with Trypanosoma rangeli modulates the profiles of immunoglobulins and IL-6 at local and systemic levels in the early phase of Trypanosoma cruzi experimental infection. Mem. Inst. Oswaldo Cruz. 2011, 106, 32–37. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Basso, B.; Marini, V. Experimental Chagas disease. Innate immune response in Balb/c mice previously vaccinated with Trypanosoma rangeli. I. The macrophage shows immunological memory: Reality or fiction? Immunobiology 2014, 219, 275–284. [Google Scholar] [CrossRef]
- Aparicio-Burgos, J.E.; Zepeda-Escobar, J.A.; De Oca-Jiménez, R.M.; Estrada-Franco, J.G.; Barbabosa-Pliego, A.; Ochoa-García, L.; Alejandre-Aguilar, R.; Rivas, N.; Penuelas-Rivas, G.; Val-Arreola, M.; et al. Immune Protection against Trypanosoma cruzi Induced by TcVac4 in a Canine Model. PLoS Neglected Trop. Dis. 2015, 9, e0003625. [Google Scholar] [CrossRef]
- Nuñez, J.M. Enfermedad de Chagas: Datos pertinentes y revisión de casos atendidos en el Hospital Santo Tomás 1955 a 1964. Arch. Med. Panameños 1966, 15, 35–47. [Google Scholar]
- Sousa, O. Anotaciones sobre la enfermedad de Chagas en Panamá. Frecuencia y distribución de Trypanosoma cruzi y Trypanosoma rangeli. Rev. Biol. Trop. 1972, 20, 167–179. [Google Scholar]
- Garisto-Risco, J.D.; Saldaña, A.; Zebede, S.; Calzada, J.E. Alteraciones cardiacas en pacientes seropositivos a la infección chagásica en Panamá. Rev. Esp. Cardiol. 2009, 62, 947–948. [Google Scholar] [CrossRef]
- Blandón, R.; Guevara, F.J.; Johnson, C.M. Tránsito esofágico en pacientes con enfermedad de Chagas en Panamá. Arq. Gastroent. 1969, 6, 189–196. [Google Scholar]
- Sousa, O.E.; Samudio, F.; De Juncá, C.; Calzada, J.E. Molecular characterization of human Trypanosoma cruzi isolates from endemic areas in Panama. Mem. Inst. Oswaldo Cruz. 2006, 101, 455–457. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Samudio, F.; Ortega-Barría, E.; Saldaña, A.; Calzada, J. Predominance of Trypanosoma cruzi I among Panamanian sylvatic isolates. Acta Trop. 2007, 101, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Brandão, A.; Samudio, F.; Fernandes, O.; Calzada, J.E.; Sousa, O.E. Genotyping of Panamanian Trypanosoma cruzi stocks using the calmodulin 3′UTR polymorphisms. Parasitol. Res. 2007, 102, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Saldaña, A.; Santamaría, A.M.; Pineda, V.; Vásquez, V.; Gottdenker, N.L.; Calzada, J.E. A darker chromatic variation of Rhodnius pallescens infected by specific genetic groups of Trypanosoma rangeli and Trypanosoma cruzi from Panama. Parasites Vectors 2018, 11, 423. [Google Scholar] [CrossRef] [PubMed]
- Prescilla-Ledezma, A.; Blandon, R.; Schijman, A.G.; Benatar, A.; Saldaña, A.; Osuna, A. Correction: Mixed infections by different Trypanosoma cruzi discrete typing units among Chagas disease patients in an endemic community in Panama. PLoS ONE 2021, 16, e0250184. [Google Scholar] [CrossRef]
- Zingales, B.; Andrade, S.G.; Briones, M.R.S.; Campbell, A.D.; Chiari, E.; Fernandes, O.; Guhl, F.; Lages-Silva, E.; Macedo, A.M.; Machado, C.R.; et al. A new consensus for Trypanosoma cruzi intraspecific nomenclature: Second revision meeting recommends TcI to TcVI. Mem. Inst. Oswaldo Cruz. 2009, 104, 1051–1054. [Google Scholar] [CrossRef]
- Zingales, B.; Miles, M.A.; Campbell, D.A.; Tibayrenc, M.; Macedo, A.M.; Teixeira, M.M.; Schijman, A.G.; Llewellyn, M.S.; Lages-Silva, E.; Machado, C.R.; et al. The revised Trypanosoma cruzi subspecific nomenclature: Rationale, epidemiological relevance and research applications. Infect. Genet. Evol. 2012, 12, 240–253. [Google Scholar] [CrossRef]
- Brenière, S.F.; Waleckx, E.; Barnabé, C. Over Six Thousand Trypanosoma cruzi Strains Classified into Discrete Typing Units (DTUs): Attempt at an Inventory. PLoS Neglected Trop. Dis. 2016, 10, e0004792. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; Miles, M.A.; Carrasco, H.J.; Lewis, M.D.; Yeo, M.; Vargas, J.; Torrico, F.; Diosque, P.; Valente, V.; Valente, S.A.; et al. Genome-Scale Multilocus Microsatellite Typing of Trypanosoma cruzi Discrete Typing Unit I Reveals Phylogeographic Structure and Specific Genotypes Linked to Human Infection. PLoS Pathog. 2009, 5, e1000410. [Google Scholar] [CrossRef]
- Messenger, L.A.; Llewellyn, M.S.; Bhattacharyya, T.; Franzén, O.; Lewis, M.D.; Ramírez, J.D.; Carrasco, H.J.; Andersson, B.; Miles, M.A. Multiple Mitochondrial Introgression Events and Heteroplasmy in Trypanosoma cruzi Revealed by Maxicircle MLST and Next Generation Sequencing. PLoS Neglected Trop. Dis. 2012, 6, e1584. [Google Scholar] [CrossRef]
- Zingales, B. Trypanosoma cruzi genetic diversity: Something new for something known about Chagas disease manifestations, serodiagnosis and drug sensitivity. Acta Trop. 2018, 184, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Telleria, J.; Tibayrenc, M. American Trypanosomiasis Chagas Disease: One Hundred Years of Research, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–826. Available online: https://www.elsevier.com/books/americantrypanosomiasis-chagas-disease/telleria/978-0-12-801029-7 (accessed on 15 December 2021).
- Silveira, A.C. BID/OPS/IDRC/CNZ: Programa Regional para el Control de la Enfermedad de Chagas en América Latina; Iniciativa de Bienes Públicos Regionales, Ed.; BID: Montevideo, Uruguay, 2010; p. 242. [Google Scholar]
- Hurtado, L.A.; Calzada, J.E.; Pineda, V.; González, K.; Santamaría, A.M.; Cáceres, L.; Wald, C.; Saldaña, A. Conocimientos y factores de riesgo relacionados con la enfermedad de Chagas en dos comunidades panameñas donde Rhodnius pallescens es el vector principal. Biomédica 2014, 34, 260–270. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gottdenker, N.L.; Carroll, C.R.; Saldaña, A.; Calzada, J.E. Association of Anthropogenic Land Use Change and Increased Abundance of the Chagas Disease Vector Rhodnius pallescens in a Rural Landscape of Panama. Am. J. Trop. Med. Hyg. 2011, 84, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Gottdenker, N.; Chaves, L.F.; Calzada, J.E.; Saldaña, A.; Carroll, C.R. Host Life History Strategy, Species Diversity, and Habitat Influence Trypanosoma cruzi Vector Infection in Changing Landscapes. PLoS Neglected Trop. Dis. 2012, 6, e1884. [Google Scholar] [CrossRef]
- Lewis, M.D.; Ma, J.; Yeo, M.; Carrasco, H.J.; Llewellyn, M.S.; Miles, M.A. Genotyping of Trypanosoma cruzi: Systematic Selection of Assays Allowing Rapid and Accurate Discrimination of All Known Lineages. Am. J. Trop. Med. Hyg. 2009, 81, 1041–1049. [Google Scholar] [CrossRef]
- Costales, J.A.; Kotton, C.N.; Zurita-Leal, A.; Garcia-Perez, J.; Llewellyn, M.S.; Messenger, L.A.; Bhattacharyya, T.; Burleigh, B.A. Chagas disease reactivation in a heart transplant patient infected by domestic Trypanosoma cruzi discrete typing unit I (TcIDOM). Parasites Vectors 2015, 8, 435. [Google Scholar] [CrossRef]
- Konechnikov, K.; Golosova, O.; Fursov, M.; Varlamov, A.; Vaskin, Y.; Efremov, I.; German Grehov, O.G.; Kandrov, D.; Rasputin, K.; Syabro, M.; et al. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Polzin, T.; Daneschmand, S.V. On Steiner trees and minimum spanning trees in hypergraphs. Op. Res. Lett. 2003, 31, 12–20. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- O’Dea, A.; Lessios, H.A.; Coates, A.G.; Eytan, R.I.; Restrepo-Moreno, S.A.; Cione, A.L.; Collins, L.S.; de Queiroz, A.; Farris, D.W.; Norris, R.D.; et al. Formation of the Isthmus of Panama. Sci. Adv. 2016, 2, e1600883. [Google Scholar] [CrossRef] [PubMed]
- Tomasini, N.; Diosque, P. Evolution of Trypanosoma cruzi: Clarifying hybridisations, mitochondrial introgressions and phylogenetic relationships between major lineages. Mem. Inst. Oswaldo Cruz. 2015, 110, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.D.; Llewellyn, M.S.; Yeo, M.; Acosta, N.; Gaunt, M.W.; Miles, M.A. Recent, Independent and Anthropogenic Origins of Trypanosoma cruzi Hybrids. PLoS Neglected Trop. Dis. 2011, 5, e1363. [Google Scholar] [CrossRef] [PubMed]
- Messenger, L.A.; Miles, M.A.; Bern, C. Between a bug and a hard place: Trypanosoma cruzi genetic diversity and the clinical outcomes of Chagas disease. Expert Rev. Anti. Infect. Ther. 2015, 13, 995–1029. [Google Scholar] [CrossRef] [PubMed]
- Majeau, A.; Murphy, L.; Herrera, C.; Dumonteil, E. Assessing Trypanosoma cruzi Parasite Diversity through Comparative Genomics: Implications for Disease Epidemiology and Diagnostics. Pathogens 2021, 10, 212. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.; Truyens, C.; Dumonteil, E.; Alger, J.; Sosa-Estani, S.; Cafferata, M.L.; Gibbons, L.; Ciganda, A.; Matute, M.L.; Zuniga, C.; et al. Phylogenetic Analysis of Trypanosoma cruzi from Pregnant Women and Newborns from Argentina, Honduras, and Mexico Suggests an Association of Parasite Haplotypes with Congenital Transmission of the Parasite. J. Mol. Diagn. 2019, 21, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Caballero, Z.C.; Sousa, O.E.; Marques, W.P.; Saez-Alquezar, A.; Umezawa, E.S. Evaluation of Serological Tests To Identify Trypanosoma cruzi Infection in Humans and Determine Cross-Reactivity with Trypanosoma rangeli and Leishmania spp. Clin. Vaccine Immunol. 2007, 14, 1045–1049. [Google Scholar] [CrossRef]
- Aguilar, H.M.; Abad-Franch, F.; Dias, J.C.; Junqueira, A.C.; Coura, J.R. Chagas disease in the Amazon region. Mem. Inst. Oswaldo Cruz. 2007, 102, 47–56, Erratum in Mem. Inst. Oswaldo Cruz. 2007, 102, 1009. [Google Scholar] [CrossRef]
- Talavera-López, C.; Messenger, L.A.; Lewis, M.D.; Yeo, M.; Reis-Cunha, J.L.; Matos, G.M.; Bartholomeu, D.C.; Calzada, J.E.; Saldaña, A.; Ramírez, J.D.; et al. Repeat-Driven Generation of Antigenic Diversity in a Major Human Pathogen, Trypanosoma cruzi. Front. Cell. Infect. Microbiol. 2021, 11, 614665. [Google Scholar] [CrossRef]
- Dumonteil, E.; Ramírez-Sierra, M.J.; Pérez-Carrillo, S.; Teh-Poot, C.; Herrera, C.; Gourbiere, S.; Waleckx, E. Detailed ecological associations of triatomines revealed by metabarcoding based on next-generation sequencing: Linking triatomine behavioral ecology and Trypanosoma cruzi transmission cycles. Sci. Rep. 2018, 8, 4140. [Google Scholar] [CrossRef]
- Villanueva-Lizama, L.; Teh-Poot, C.; Majeau, A.; Herrera, C.; Dumonteil, E. Molecular genotyping of Trypanosoma cruzi by next-generation sequencing of the mini-exon gene reveals infections with multiple parasite DTUs in Chagasic patients from Yucatan, Mexico. J. Infect. Dis. 2019, 219, 1980–1988. [Google Scholar] [CrossRef] [PubMed]
- Murillo-Solano, C.; Ramos-Ligonio, A.; López-Monteon, A.; Guzmán-Gómez, D.; Torres-Montero, J.; Herrera, C.; Dumonteil, E. Diversity of Trypanosoma cruzi parasites infecting Triatoma dimidiata in Central Veracruz, Mexico, and their One Health ecological interactions. Infect. Genet. Evol. 2021, 95, 105050. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, J.D.; Guhl, F.; Rendón, L.M.; Rosas, F.; Marin-Neto, J.A.; Morillo, C.A. Chagas Cardiomyopathy Manifestations and Trypanosoma cruzi Genotypes Circulating in Chronic Chagasic Patients. PLoS Neglected Trop. Dis. 2010, 4, e899. [Google Scholar] [CrossRef]
- Messenger, L.A.; Ramirez, J.D.; Llewellyn, M.S.; Guhl, F.; Miles, M.A. Importation of Hybrid Human-Associated Trypanosoma cruzi Strains of Southern South American Origin, Colombia. Emerg. Infect. Dis. 2016, 22, 1452–1455. [Google Scholar] [CrossRef] [PubMed]
- Hernández, C.; Cucunubá, Z.; Florez, C.; Olivera, M.; Valencia-Hernandez, C.A.; Zambrano, P.; León, C.; Ramirez, J.D. Correction: Molecular Diagnosis of Chagas Disease in Colombia: Parasitic Loads and Discrete Typing Units in Patients from Acute and Chronic Phases. PLoS Neglected Trop. Dis. 2016, 10, e0005112. [Google Scholar] [CrossRef] [PubMed]
- Gaunt, M.; Miles, M. The ecotopes and evolution of triatomine bugs (triatominae) and their associated trypanosomes. Mem. Inst. Oswaldo Cruz. 2000, 95, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Vivas, R.J.; García, J.E.; Guhl, F.; Hernández, C.; Velásquez, N.; Ramírez, J.D.; Carranza, J.C.; Vallejo, G.A. Systematic review on the biology, ecology, genetic diversity and parasite transmission potential of Panstrongylus geniculatus (Latreille 1811) in Latin America. Mem. Inst. Oswaldo Cruz. 2021, 116, e200528. [Google Scholar] [CrossRef]
- Peterson, J.K.; Hashimoto, K.; Yoshioka, K.; Dorn, P.L.; Gottdenker, N.L.; Caranci, A.; Stevens, L.; Zuniga, C.; Saldaña, A.; Rodriguez, S.; et al. Chagas Disease in Central America: Recent Findings and Current Challenges in Vector Ecology and Control. Curr. Trop. Med. Rep. 2019, 6, 76–91. [Google Scholar] [CrossRef]
- Martins, K.; Andrade, C.D.M.; Barbosa-Silva, A.N.; Nascimento, G.B.D.; Chiari, E.; Galvão, L.M.D.C.; da Câmara, A.C.J. Trypanosoma cruzi III causing the indeterminate form of Chagas disease in a semi-arid region of Brazil. Int. J. Infect. Dis. 2015, 39, 68–75. [Google Scholar] [CrossRef][Green Version]
- Souza, K.S.; Bastazini, V.A.; Colares, E.P. Feeding ecology of the Neotropical otter Lontra longicaudis in the Lower Arroio Grande River, southern Brazil. An. Acad. Bras. Cienc. 2013, 85, 285–294. [Google Scholar] [CrossRef]
- Schofield, C.J.; Dujardin, J.P. Theories on the evolution of Rhodnius. Actual. Biol. 2017, 21, 183–197. [Google Scholar]
- Da Silva, F.M.; Junqueira, A.C.V.; Campaner, M.; Rodrigues, A.C.; Crisante, G.; Ramirez, L.E.; Caballero, Z.C.E.; Monteiro, F.A.; Coura, J.R.; Añez, N.; et al. Comparative phylogeography of Trypanosoma rangeli and Rhodnius (Hemiptera: Reduviidae) supports a long coexistence of parasite lineages and their sympatric vectors. Mol. Ecol. 2007, 16, 3361–3373. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Antón, F.; Urrea, D.A.; Guhl, F.; Arévalo, C.; Azofeifa, G.; Urbina, A.; Blandón-Naranjo, M.; Sousa, O.E.; Zeledón, R.; Vallejo, G.A. Trypanosoma rangeli genotypes association with Rhodnius prolixus and R. pallescens allopatric distribution in Central America. Infect. Genet. Evol. 2009, 9, 1306–1310. [Google Scholar] [CrossRef] [PubMed]
- Dario, M.; Pavan, M.; Rodrigues, M.; Lisboa, C.; Kluyber, D.; Desbiez, A.; Herrera, H.; Roque, A.; Lima, L.; Teixeira, M.; et al. Trypanosoma rangeli Genetic, Mammalian Hosts, and Geographical Diversity from Five Brazilian Biomes. Pathogens 2021, 10, 736. [Google Scholar] [CrossRef]
- Calzada, J.E.; Pineda, V.; Garisto, J.D.; Samudio, F.; Santamaria, A.M.; Saldaña, A. Human Trypanosomiasis in the Eastern Region of the Panama Province: New Endemic Areas for Chagas Disease. Am. J. Trop. Med. Hyg. 2010, 82, 580–582. [Google Scholar] [CrossRef]
- Rodriguez, I.G.; Saldaña, A.; Gonzalez, K.; Pineda, V.; Perea, M.; Santamaría, A.M.; De Junca, C.C.; Chaves, L.F.; Calzada, E.J. Trypanosoma cruzi Infection in Rhodnius pallescens (Heteroptera: Reduviidae) Infesting Coyol Palms in the Dry Arch of Panamá. J. Med Èntomol. 2018, 55, 691–700. [Google Scholar] [CrossRef]
- Kay, C.; Williams, T.A.; Gibson, W. Mitochondrial DNAs provide insight into trypanosome phylogeny and molecular evolution. BMC Evol. Biol. 2020, 20, 1–13. [Google Scholar] [CrossRef]
- Berná, L.; Greif, G.; Pita, S.; Faral-Tello, P.; Díaz-Viraqué, F.; Souza, R.D.C.M.D.; Vallejo, G.A.; Alvarez-Valin, F.; Robello, C. Maxicircle architecture and evolutionary insights into Trypanosoma cruzi complex. PLoS Neglected Trop. Dis. 2021, 15, e0009719. [Google Scholar] [CrossRef]





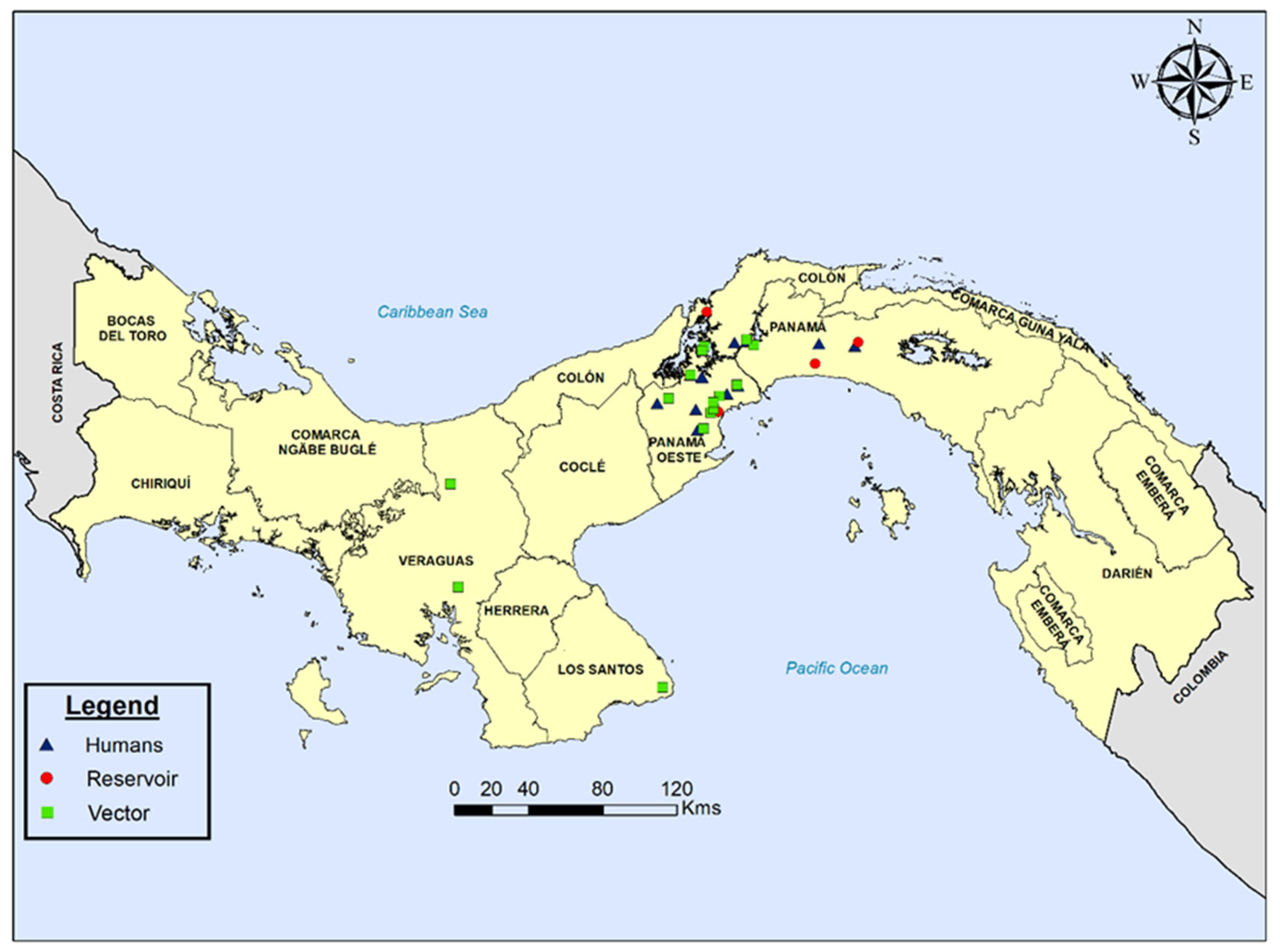{kind=link}
{kind=link}
{kind=link}
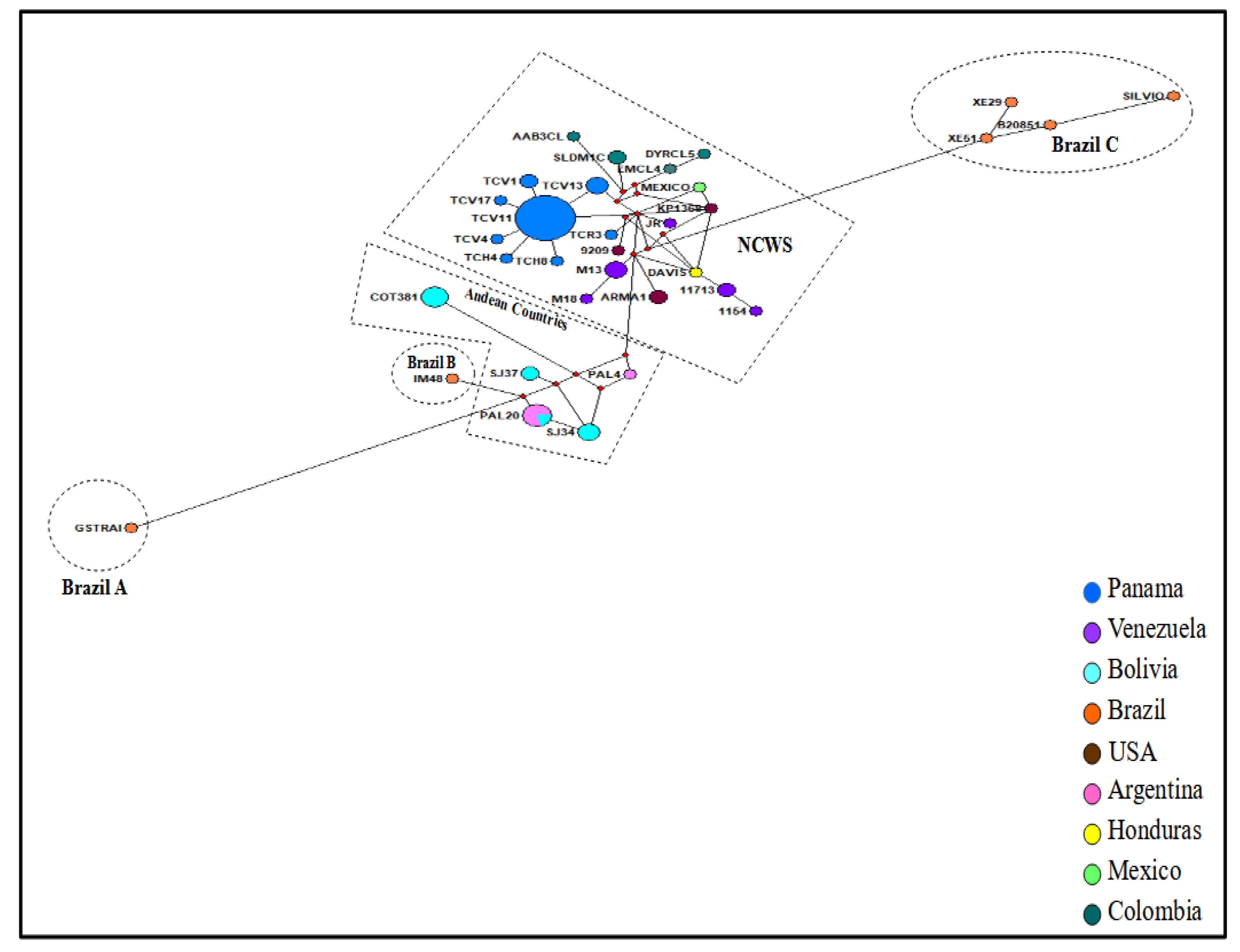{kind=link}
| Original Code | Host/Vector (Sex, Age) | Locality (Region) | Date | Clinical Status | Outcome | T. cruzi DTU |
|---|---|---|---|---|---|---|
| TCH1 | Human (Female, 9y) | Chorrera (WP) | Oct, 1980 | Acute | Chronic | TcVI |
| TCH2 | Human (Male, 78y) | Aguas Claras (Colón) | April, 1983 | Asymptomatic | Indetermined | TcI |
| TCH3 | Human (Female, 7y) | Chorrera (WP) | March, 1981 | Asymptomatic | Indetermined | TcI |
| TCH4 | Human (Male, 29y) | Chilibre (Colón) | July, l97l | Acute | Unknown | TcI |
| TCH5 | Human (Male, 20y) | Chepo (EP) | July, 1965 | Asymptomatic | Chronic | TcI |
| TCH6 | Human (Male, 42y) | Capira (WP) | Jan, 1987 | Acute | Indetermined | TcI |
| TCH7 | Human (Female, 17y) | Mendoza (WP) | July, 1970 | Asymptomatic | Indetermined | TcI |
| TCH8 | Human (Male, 18y) | Cerro Cama (WP) | April, 1979 | Asymptomatic | Unknown | TcI |
| TCH10 | Human (Female, 13y) | Pacora (EP) | Jan, 1995 | Acute | Indetermined | TcIII |
| TCH12 | Human (Male, 31y) | Bunrunga (WP) | Oct, 2002 | Acute | Unknown | TcI |
| TCH14 | Human (Female, 1y) | Nuevo Arraiján (WP) | Oct, 1973 | Acute | Chronic | TcI |
| TCH15 | Human (Male, 3y) | Capira (WP) | Aug, 1980 | Acute | Chronic | TcI |
| TCR1 | Monkey (Saguinus geoffroyi) | Pacora (EP) | June, 1966 | Asymptomatic | Unknown | TcI |
| TCR2 | Opossum (Didelphis marsupialis) | Chepo (EP) | June, 1966 | Asymptomatic | Unknown | TcI |
| TCR3 | Otter (Lontra longicaudis) | Colon | May, 1973 | Asymptomatic | Unknown | TcI |
| TCR4 | Canine (Canis familiaris) | Chorrera (WP) | 2004 | Acute | Fatal | TcI |
| TCV1 | Panstrongylus geniculatus | Barro Colorado (WP) | 1992 | TcI | ||
| TCV2 | Rhodnius pallescens | Chilibre (Colón) | 1991 | TcI | ||
| TCV3 | Triatoma dimidiata | Barro Colorado (WP) | 1992 | TcI | ||
| TCV4 | Rhodnius pallescens | Cerro Cama (WP) | 1985 | TcI | ||
| TCV5 | Rhodnius pallescens | Chilibre (Colón) | 2011 | TcI | ||
| TCV6 | Rhodnius pallescens | Chorrera (WP) | 2011 | TcI | ||
| TCV7 | Rhodnius pallescens | Chorrera (WP) | 2011 | TcI | ||
| TCV8 | Rhodnius pallescens | Burunga (WP) | 2011 | TcI | ||
| TCV9 | Rhodnius pallescens | Chorrera (WP) | 2011 | TcI | ||
| TCV10 | Rhodnius pallescens | Chorrera (WP) | 2012 | TcI | ||
| TCV11 | Rhodnius pallescens | Capira (WP) | 2012 | TcI | ||
| TCV12 | Rhodnius pallescens | Santa Fe (Veraguas) | 2014 | TcI | ||
| TCV13 | Rhodnius pallescens | Pedasi (Los Santos) | 2016 | TcI | ||
| TCV14 | Rhodnius pallescens | Capira (WP) | 2016 | TcI | ||
| TCV15 | Panstrongylus geniculatus | Montijo (Veraguas) | 2015 | TcIII | ||
| TCV17 | Rhodnius pallescens | Burunga (WP) | 2000 | TcI |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calzada, J.E.; Samudio, F.; de Juncá, C.; Pineda, V.; Burleigh, B.A.; Saldaña, A. Genetic Diversity of Trypanosoma cruzi in Panama Inferred by Multi-locus Sequence Typing of Mitochondrial Genes. Microorganisms 2022, 10, 287. https://doi.org/10.3390/microorganisms10020287
Calzada JE, Samudio F, de Juncá C, Pineda V, Burleigh BA, Saldaña A. Genetic Diversity of Trypanosoma cruzi in Panama Inferred by Multi-locus Sequence Typing of Mitochondrial Genes. Microorganisms. 2022; 10(2):287. https://doi.org/10.3390/microorganisms10020287
Chicago/Turabian StyleCalzada, Jose E., Franklyn Samudio, Corina de Juncá, Vanessa Pineda, Barbara A. Burleigh, and Azael Saldaña. 2022. "Genetic Diversity of Trypanosoma cruzi in Panama Inferred by Multi-locus Sequence Typing of Mitochondrial Genes" Microorganisms 10, no. 2: 287. https://doi.org/10.3390/microorganisms10020287
APA StyleCalzada, J. E., Samudio, F., de Juncá, C., Pineda, V., Burleigh, B. A., & Saldaña, A. (2022). Genetic Diversity of Trypanosoma cruzi in Panama Inferred by Multi-locus Sequence Typing of Mitochondrial Genes. Microorganisms, 10(2), 287. https://doi.org/10.3390/microorganisms10020287