
Endosymbiotic Bacterial Diversity of Corn Leaf Aphid, Rhopalosiphum maidis Fitch (Hemiptera: Aphididae) Associated with Maize Management Systems

 ,
,  , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Areas
2.2. Field Collection of Corn Leaf Aphids
2.3. Aphid-Associated Bacterial Community Analysis
2.4. Data Analyses
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Blackman, R.; Eastop, V. Aphids on the World’s Crops: An Identification and Information Guide, 2nd ed.; Wiley: Hoboken, NJ, USA, 2000. [Google Scholar]
- Emden, H.F.; Harrington, R. (Eds.) Aphids as Crop Pests; CABI Publishing: London, UK, 2007; ISBN 978-0-85199-819-0. [Google Scholar]
- Emden, H.F.; Harrington, R. (Eds.) Aphids as Crop Pests, 2nd ed.; CABI Publishing: London, UK, 2017; ISBN 978-0-85199-819-0. [Google Scholar]
- Russell, J.A.; Moran, N.A. Costs and Benefits of Symbiont Infection in Aphids: Variation among Symbionts and across Temperatures. Proc. R. Soc. B Biol. Sci. 2006, 273, 603–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Valero, L.; Soriano-Navarro, M.; Pérez-Brocal, V.; Heddi, A.; Moya, A.; García-Verdugo, J.M.; Latorre, A. Coexistence of Wolbachia with Buchnera aphidicola and a Secondary Symbiont in the Aphid Cinara cedri. J. Bacteriol. 2004, 186, 6626–6633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, K.M.; Moran, N.A.; Hunter, M.S. Variation in Resistance to Parasitism in Aphids Is Due to Symbionts Not Host Genotype. Proc. Natl. Acad. Sci. USA 2005, 102, 12795–12800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montllor, C.B.; Maxmen, A.; Purcell, A.H. Facultative Bacterial Endosymbionts Benefit Pea Aphids Acyrthosiphon pisum under Heat Stress. Ecol. Entomol. 2002, 27, 189–195. [Google Scholar] [CrossRef]
- Tsuchida, T.; Koga, R.; Horikawa, M.; Tsunoda, T.; Maoka, T.; Matsumoto, S.; Simon, J.-C.; Fukatsu, T. Symbiotic Bacterium Modifies Aphid Body Color. Science 2010, 330, 1102–1104. [Google Scholar] [CrossRef]
- Douglas, A.E. How Multi-Partner Endosymbioses Function. Nat. Rev. Microbiol. 2016, 14, 731–743. [Google Scholar] [CrossRef]
- Zhang, B.; Leonard, S.P.; Li, Y.; Moran, N.A. Obligate Bacterial Endosymbionts Limit Thermal Tolerance of Insect Host Species. PNAS 2019, 116, 24712–24718. [Google Scholar] [CrossRef]
- Brown, P.A.; Blackman, R.L. Karyotype Variation in the Corn Leaf Aphid, Rhopalosiphum maidis (Fitch), Species Complex (Hemiptera: Aphididae) in Relation to Host-Plant and Morphology. Bull. Entomol. Res. 1988, 78, 351–363. [Google Scholar] [CrossRef]
- Scott, F.G. Symbionts as an Epigenetic Source of Heritable Variation. Transform. Lamarckism Subtle Fluids Mol. Biol. 2011, 283–293. [Google Scholar]
- Jousselin, E.; Cruaud, A.; Genson, G.; Chevenet, F.; Foottit, R.G.; Cœur d’acier, A. Is Ecological Speciation a Major Trend in Aphids? Insights from a Molecular Phylogeny of the Conifer-Feeding Genus Cinara. Front. Zool. 2013, 10, 56. [Google Scholar] [CrossRef] [Green Version]
- Onstad, D.W. Insect Resistance Management: Biology, Economics, and Prediction; Elsevier: Amsterdam, The Netherlands, 2014; ISBN 978-0-12-396955-2. [Google Scholar]
- Field, L.M.; Blackman, R.L.; Tyler-Smith, C.; Devonshire, A.L. Relationship between Amount of Esterase and Gene Copy Number in Insecticide-Resistant Myzus persicae (Sulzer). Biochem. J. 1999, 339 Pt 3, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.H.; Chiu, M.C.; Perng, J.J. Temperature Effects on Life History Traits of the Corn Leaf Aphid, Rhopalosiphum maidis (Homoptera: Aphididae) on Corn in Taiwan. Appl. Entomol. Zool. 2006, 41, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Benedek, K.; Bálint, J.; Máthé, I.; Mara, G.; Felföldi, T.; Szabó, A.; Fazakas, C.; Albert, C.; Buchkowski, R.W.; Schmitz, O.J.; et al. Linking Intraspecific Variation in Plant Chemical Defence with Arthropod and Soil Bacterial Community Structure and N Allocation. Plant Soil 2019, 444, 383–397. [Google Scholar] [CrossRef] [Green Version]
- Csorba, A.B. Endosymbiotic Bacterial Diversity Associated with Corn Leaf Aphid, Rhopalosiphum maidis Fitch (Hemiptera: Aphididae) Populations under Different Maize Management Systems—Preliminary Study. North-West. J. Zool 2021, 17, 5. [Google Scholar]
- Herlemann, D.P.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in Bacterial Communities along the 2000 Km Salinity Gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor Revision to V4 Region SSU RRNA 806R Gene Primer Greatly Increases Detection of SAR11 Bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D. Reintroducing Mothur: 10 Years Later. Appl. Environ. Microbiol. 2020, 86, e02343-19. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Paul, D.R. Past: Paleontological Statistics Software Package for Education And Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- McLean, A.H.C.; van Asch, M.; Ferrari, J.; Godfray, H.C.J. Effects of Bacterial Secondary Symbionts on Host Plant Use in Pea Aphids. Proc. R. Soc. B Biol. Sci. 2011, 278, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Shigenobu, S.; Watanabe, H.; Hattori, M.; Sakaki, Y.; Ishikawa, H. Genome Sequence of the Endocellular Bacterial Symbiont of Aphids Buchnera sp. APS. Nature 2000, 407, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loxdale, H.D.; Balog, A. Aphid Specialism as an Example of Ecological–Evolutionary Divergence. Biol. Rev. 2018, 93, 642–657. [Google Scholar] [CrossRef] [PubMed]
- Loxdale, H.D.; Balog, A.; Biron, D.G. Aphids in Focus: Unravelling Their Complex Ecology and Evolution Using Genetic and Molecular Approaches. Biol. J. Linn. Soc. 2020, 129, 507–531. [Google Scholar] [CrossRef]
- Chong, R.A.; Moran, N.A. Evolutionary Loss and Replacement of Buchnera, the Obligate Endosymbiont of Aphids. ISME J. 2018, 12, 898–908. [Google Scholar] [CrossRef] [Green Version]
- Moran, N.A.; Bennett, G.M. The Tiniest Tiny Genomes. Annu. Rev. Microbiol. 2014, 68, 195–215. [Google Scholar] [CrossRef]
- Nováková, E.; Hypša, V.; Klein, J.; Foottit, R.G.; von Dohlen, C.D.; Moran, N.A. Reconstructing the Phylogeny of Aphids (Hemiptera: Aphididae) Using DNA of the Obligate Symbiont Buchnera aphidicola. Mol. Phylogenet. Evol. 2013, 68, 42–54. [Google Scholar] [CrossRef]
- Wegierek, P.; Michalik, A.; Wieczorek, K.; Kanturski, M.; Kobiałka, M.; Śliwa, K.; Szklarzewicz, T. Buchnera aphidicola of the Birch Blister Aphid, Hamamelistes betulinus (Horváth, 1896) (Insecta, Hemiptera, Aphididae: Hormaphidinae): Molecular Characterization, Transmission between Generations and Its Geographic Significance. Acta Zool. 2017, 98, 412–421. [Google Scholar] [CrossRef]
- Burke, G.; Fiehn, O.; Moran, N. Effects of Facultative Symbionts and Heat Stress on the Metabolome of Pea Aphids. ISME J. 2010, 4, 242–252. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Chen, J.; Qin, M.; Jiang, L.; Qiao, G. Geography-Dependent Symbiont Communities in Two Oligophagous Aphid Species. FEMS Microbiol. Ecol. 2021, 97, fiab132. [Google Scholar] [CrossRef]
- Chen, D.-Q.; Montllor, C.B.; Purcell, A.H. Fitness Effects of Two Facultative Endosymbiotic Bacteria on the Pea Aphid, Acyrthosiphon pisum, and the Blue Alfalfa Aphid, A kondoi. Entomol. Exp. et Appl. 2000, 95, 315–323. [Google Scholar] [CrossRef]
- Ferrari, J.; Darby, A.C.; Daniell, T.J.; Godfray, H.C.J.; Douglas, A.E. Linking the Bacterial Community in Pea Aphids with Host-Plant Use and Natural Enemy Resistance. Ecol. Entomol. 2004, 29, 60–65. [Google Scholar] [CrossRef]
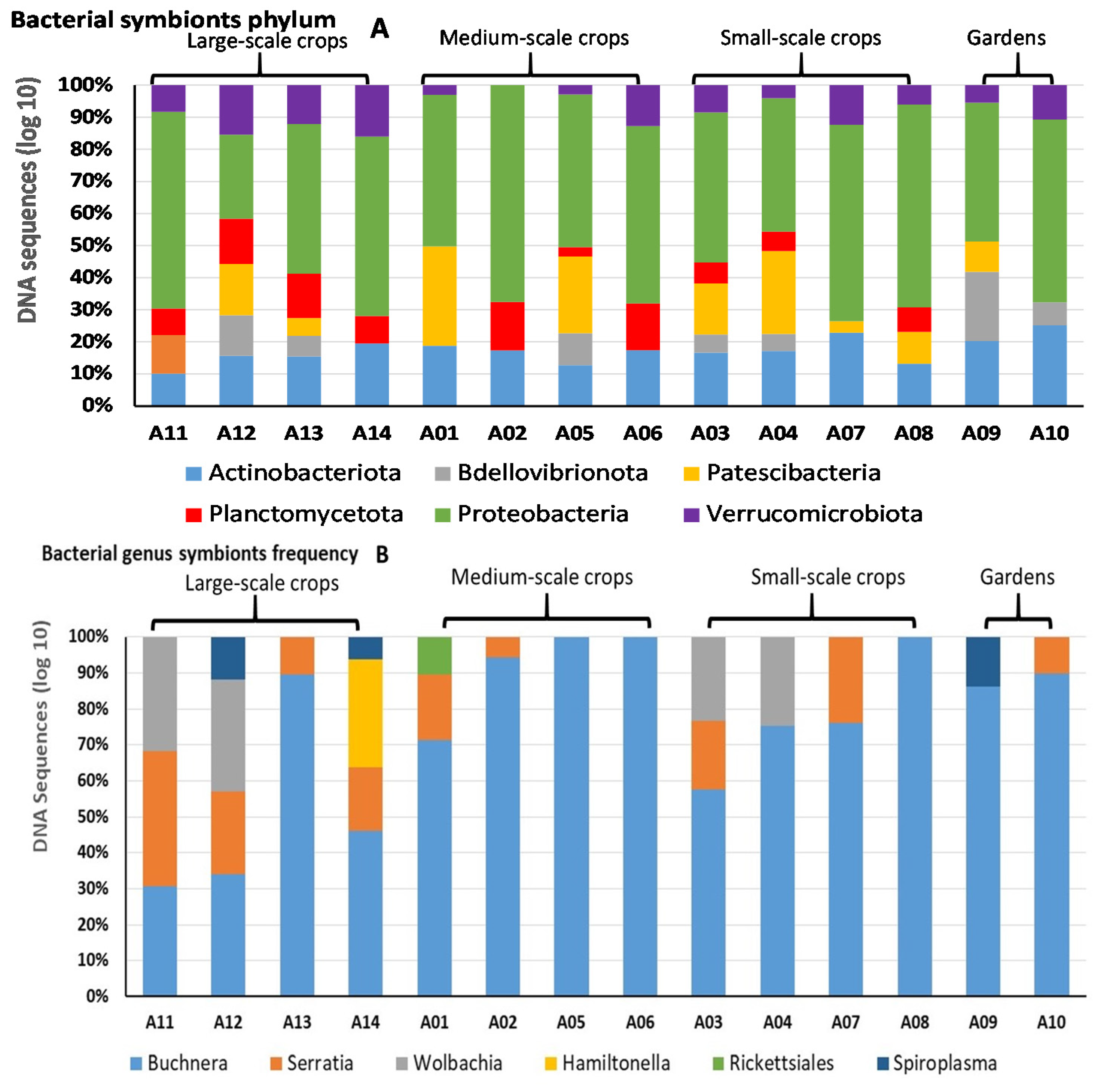


{kind=link}
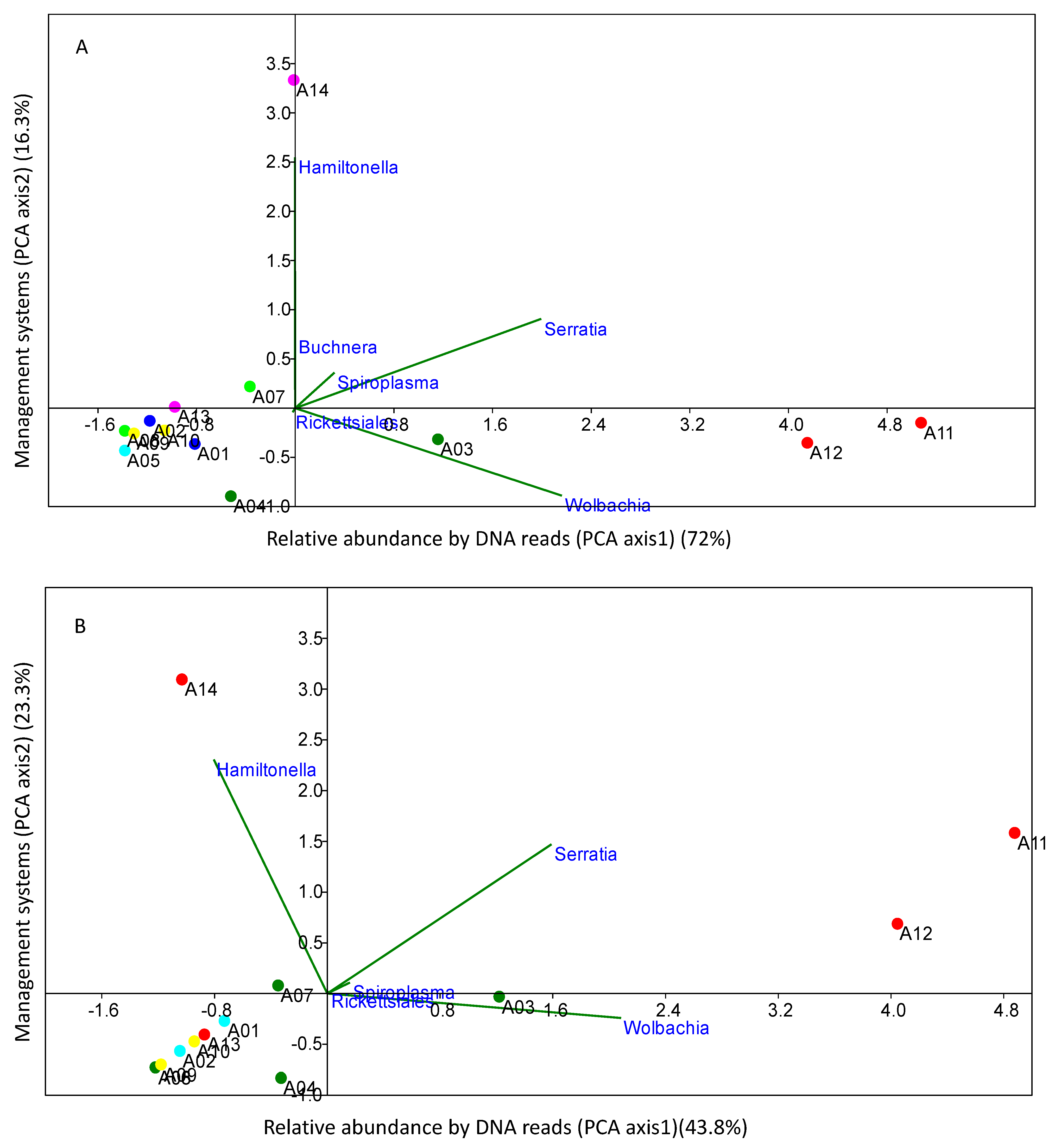
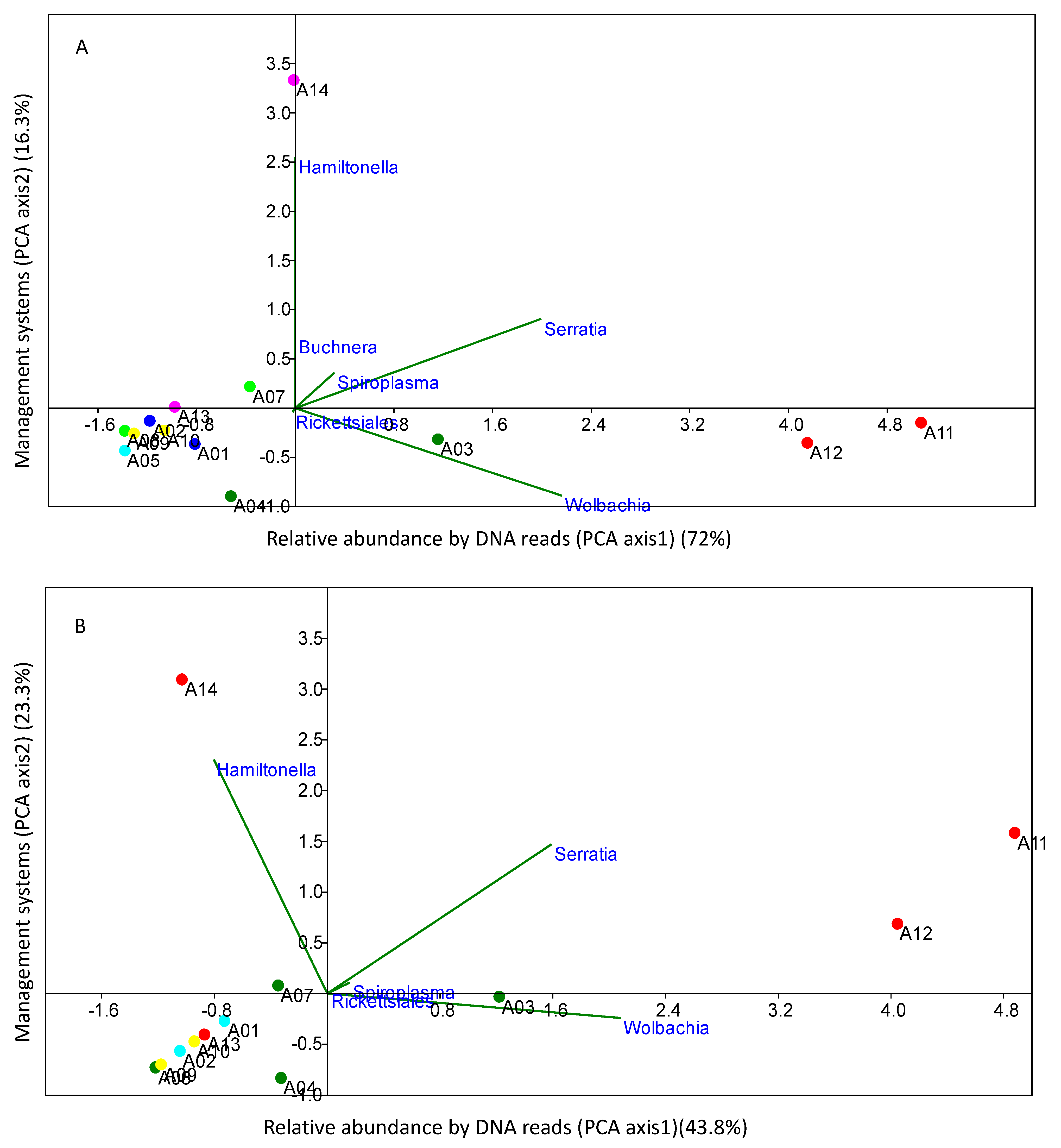
{kind=link}
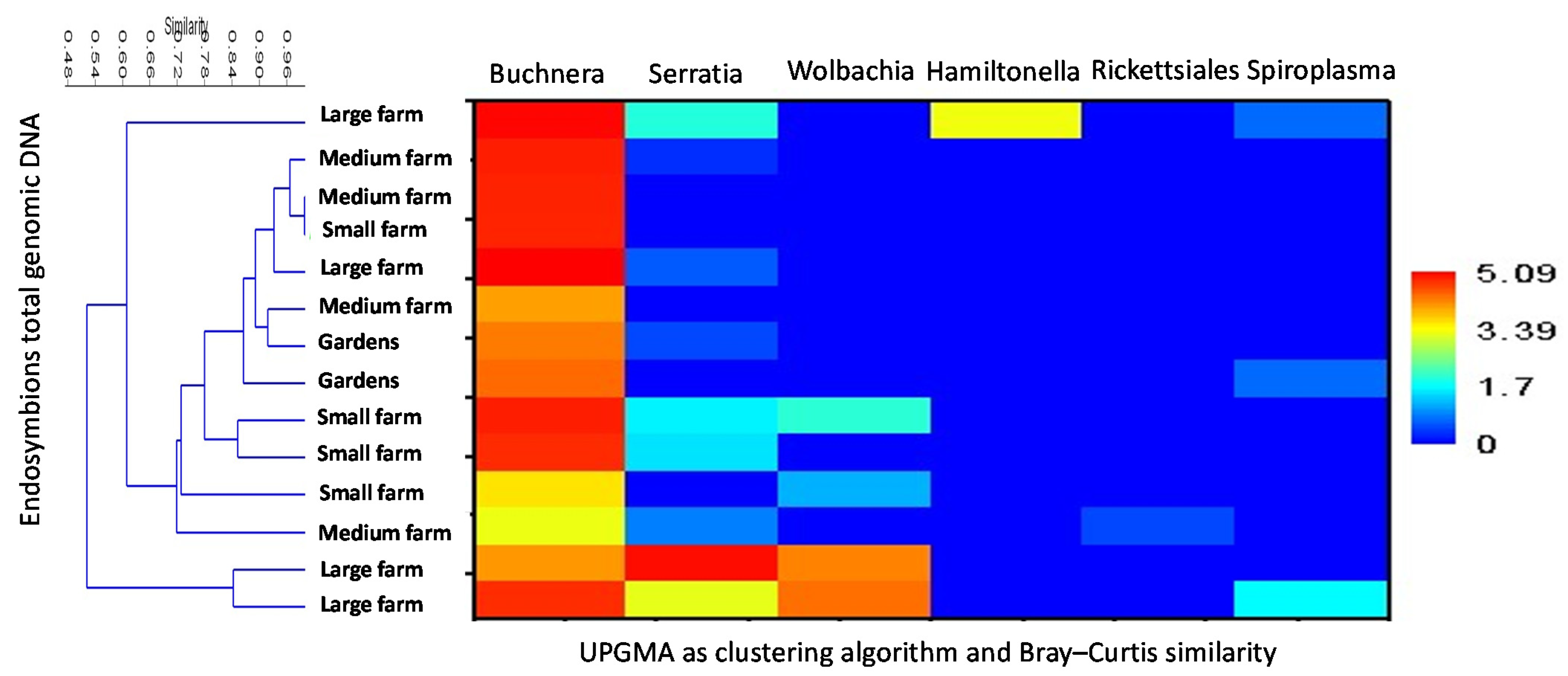
{kind=link}
| Cropping System | Management | Sample Code | Symbionts Detected |
|---|---|---|---|
| Large farms 2 year old culture, previous year crop sunflower Hybrid: P 9911 | Seed treatment: Imidacloprid 600 g/L (Seedoprid 600 FS 10 L/t) Herbicide pre-emergent: Isoxaflutole 225 g/L + Thiencarbazone-methyl 90 g/L + Cyprosulfamide (safener) 150 g/L (Adengo 0.35 L/ha) Herbicide post-emergent: Replaced by hoeing. Fertilizers: NPK 16:16:16 250 Kg/ha at soil preparation and N 33.3% 250 Kg/ha at hoeing and Foliar fertilizer N (195 g/L), MgSO4 (26 g/L), SO3 (55 g/L) + microelements (Boron, Copper, Iron, Manganese, Molybdenum, Zinc (14.3 g/L) (Plonvit Active Maize 3 L/ha) Insecticide: lambda—cyhalothrin (50 g/L) (Karate Zeon 250 mL/ha) against Diabrotica at tasseling | A11 A12 | Buchnera, Serratia, Wolbachia, Buchnera, Serratia, Wolbachia, Spiroplasma |
| Large farms 4 year old culture Hybrid: PR 37 N 01 | Seed treatment: Fludioxinil + Metalaxyl-M + Thiabendazole+Azoxystrobin Herbicidepre-emergent: Isoxaflutole 225 g/L + Thiencarbazone-methyl 90 g/L + Cyprosulfamide (safener) 150 g/L (Adengo 0.35 L/ha) Herbicide post-emergent: Nicosulfuron 40 g/L (Nicogan 40 OD 1 L/ha) and Florasulam 6.25 g/L + Acid 2,4-D EHE 300 g/L (Mustang 0.5 L/ha) Fertilizers: NPK 16:16:16 300 Kg/ha at soil preparation | A13 A14 | Buchnera, Serratia Buchnera, Serratia, Hamiltonella, Spiroplasma |
| Medium farms 3 year old culture Hybrid: LG 33.50 | Seed treatment: Prothioconazole + Metalaxyl. Herbicide pre-emergent: Isoxaflutole + Thiencarbazone-methyl. Fertilizers: 50 t/ha organic fertilizer at seeding. At 12 leaf stage: Inorganic fertilizer NH4NO3+CaMg(CO3) 2200 kg/ha. | A01 A02 | Buchnera, Serratia, Rickettsiales other than Spiroplasma Buchnera, Serratia |
| Medium farms 5 year old culture Hybrid: LG SHANNON | Seed treatment: Prothiocanazole + Metalaxyl. Herbicide pre-emergent: Isoxaflutole + Thiencarbazone-methyl. At 4–6 leaf stage Nicosulfuron. Fertilizers:70 t/ha organic fertilizer at seeding. At 12 leaf stage: Inorganic fertilizer NH4NO3+CaMg(CO3) 2200 kg/ha. | A05 A06 | Buchnera Buchnera |
| Small farms 1 year old culture, previous year crop wheat and before maize Hybrid: Not known | Only organic fertilizers 50 t/ha before seeding. | A03 A04 | Buchnera, Serratia, Wolbachia Buchnera, Wolbachia |
| Small farms 1 year old culture, previous year crop wheat and before maize Hybrid: Not known | Only organic fertilizers 50 t/ha before seeding. | A07 A08 | Buchnera, Serratia Buchnera |
| Gardens 1 year old culture, previous year crop potato. Hybrid: Not known | No treatment (Control crop) | A09 A10 | Buchnera, Spiroplasma Buchnera, Serratia |
| Farming | Large Scale-Farming | Medium-Scale Farming | Small-Scale Farming | Gardens | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Codes | A11 | A12 | A13 | A14 | A01 | A02 | A05 | A06 | A03 | A04 | A07 | A08 | A09 | A10 |
| No. of genera | 28 | 166 | 47 | 55 | 47 | 31 | 46 | 59 | 60 | 59 | 63 | 33 | 56 | 50 |
| Simpson | 0.920 | 0.993 | 0.964 | 0.969 | 0.966 | 0.937 | 0.966 | 0.967 | 0.971 | 0.971 | 0.973 | 0.944 | 0.969 | 0.966 |
| Shannon_H | 2.922 | 4.998 | 3.627 | 3.733 | 3.582 | 3.079 | 3.578 | 3.779 | 3.802 | 3.754 | 3.872 | 3.168 | 3.716 | 3.592 |
| Evenness_e^H/S | 0.664 | 0.893 | 0.800 | 0.760 | 0.765 | 0.701 | 0.778 | 0.742 | 0.746 | 0.724 | 0.763 | 0.720 | 0.734 | 0.726 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Csorba, A.B.; Fora, C.G.; Bálint, J.; Felföldi, T.; Szabó, A.; Máthé, I.; Loxdale, H.D.; Kentelky, E.; Nyárádi, I.-I.; Balog, A. Endosymbiotic Bacterial Diversity of Corn Leaf Aphid, Rhopalosiphum maidis Fitch (Hemiptera: Aphididae) Associated with Maize Management Systems. Microorganisms 2022, 10, 939. https://doi.org/10.3390/microorganisms10050939
Csorba AB, Fora CG, Bálint J, Felföldi T, Szabó A, Máthé I, Loxdale HD, Kentelky E, Nyárádi I-I, Balog A. Endosymbiotic Bacterial Diversity of Corn Leaf Aphid, Rhopalosiphum maidis Fitch (Hemiptera: Aphididae) Associated with Maize Management Systems. Microorganisms. 2022; 10(5):939. https://doi.org/10.3390/microorganisms10050939
Chicago/Turabian StyleCsorba, Artúr Botond, Ciprian George Fora, János Bálint, Tamás Felföldi, Attila Szabó, István Máthé, Hugh D. Loxdale, Endre Kentelky, Imre-István Nyárádi, and Adalbert Balog. 2022. "Endosymbiotic Bacterial Diversity of Corn Leaf Aphid, Rhopalosiphum maidis Fitch (Hemiptera: Aphididae) Associated with Maize Management Systems" Microorganisms 10, no. 5: 939. https://doi.org/10.3390/microorganisms10050939
APA StyleCsorba, A. B., Fora, C. G., Bálint, J., Felföldi, T., Szabó, A., Máthé, I., Loxdale, H. D., Kentelky, E., Nyárádi, I.-I., & Balog, A. (2022). Endosymbiotic Bacterial Diversity of Corn Leaf Aphid, Rhopalosiphum maidis Fitch (Hemiptera: Aphididae) Associated with Maize Management Systems. Microorganisms, 10(5), 939. https://doi.org/10.3390/microorganisms10050939