Abstract
Background: Botrytis cinerea (B. cinerea) is responsible for grape infection and damage to the winemaking and table grape sectors. Although anti-Botrytis chemicals are available, they are considered unsustainable for resistance phenomenon and adverse effects on the environment and human health. Research is focused on developing alternative approaches, such as exploiting biological control agents (BCAs). In this context, 19 yeasts of the genera Cryptococcus, Aureobasidium, Metschnikowia, Kluyveromyces and Wickerhamomyces were tested as antimicrobial agents against B. cinerea development. Methods: A combination of in vitro tests based on dual-culture methods, volatile organic compound production assay, laboratory tests on grape berries (punctured and sprayed with yeasts) and field experiments based on yeast treatments on grapes in vineyards allowed the selection of two potential BCAs. Results: M. pulcherrima DiSVA 269 and A. pullulans DiSVA 211 exhibited the best ability to contain the development of B. cinerea, showing the severity, the decay and the McKinney index lower than a commercial biological formulation consisting of a mixture of two different A. pullulans strains, which were used as positive controls. Conclusions: The results indicated that the selected strains were effective BCA candidates to counteract B. cinerea in the field, applying them in the partial or total replacement of conventional treatments.
1. Introduction
The fungus Botrytis cinerea (B. cinerea), also known as gray mold, is a natural component of grape microbiota [1] and is one of the main spoilage microorganisms that can cause consistent damage to crops worldwide [2,3]. B. cinerea occupies the second place in the world ranking of the top 10 pathogenic fungi in terms of diffusion and relative commercial loss, preceded only by Magnaporthe oryzae [4]. In the grape and wine industry, the impact of bunch rot is well established, because all cultivars are susceptible to this infection. A common way to counteract the development of B. cinerea is the application of chemical fungicides. Recently, it was introduced that a class of synthetic fungicides belongs to the succinate dehydrogenase inhibitors (SDHIs). Other chemicals, such as salts solutions recognized as safe (iron sulphate, ammonium bicarbonate, sodium silicate, sodium bicarbonate and sodium carbonate), are widely used to sanitise grapes surface. Ethanol vapours and other gas such as chlorine dioxide and ozone fumigation are also used, even if the sulphur dioxide remains the main method that is used [5].
These conventional anti-Botrytis treatments are considered unsustainable. Indeed, the frequent appearance of resistant strains, adverse effects of fungicides on the environment and human health and stuck fermentation using infected grapes have necessitated a new control strategy [3,5]. In this context, the development of complementary methods to synthetic agents, such as biological control agents (BCAs), could be considered an alternative approach to reducing gray mold [6,7,8,9]. However, only a few commercial products, based on fungal or bacterial genera, are available in Europe for the biological control of gray mold in vineyards [10]. Indeed, the applicability in cyclical seasons, different climatic trends and/or local agronomic conditions represent the actual limits to the full efficacy of natural treatments. Therefore, several studies have proposed natural microorganisms as effective, low-cost biological antagonists to counteract B. cinerea infection [6,8,9,10].
The use of yeast as a BCA offers some advantages, including the easy colonisation of dry surfaces for extended periods, simple nutrition requirements, rapid growth and potential antagonistic effects against pathogens [11]. Among the different antagonistic yeasts, Metschnikowiapulcherrima is a relevant yeast species that has been successfully applied to control pathogens of fruits and vegetables. The competition of M. pulcherrima for nutrients is the dominant mechanism during the biocontrol of B. cinerea [12]. In addition, Wickerhamomycesanomalus shows an antimicrobial activity against B. cinerea via multiple mechanisms, including the colonisation of wounds, a biological protective layer and the production of hydrolytic enzymes and volatile organic compounds [6]. Among other yeast genera, Debaryomyces hansenii [13], Aureobasidium pullulans [14], Cryptococcus laurentii [15] and Sporidiobolus pararoseus [16] have demonstrated the potential for the control of post-harvest decay of fruits and vegetables. In some cases, the attachment efficacy of yeasts depends on their abilities to secrete lytic enzymes, which might be involved in the biocontrol efficacy of antagonistic yeasts [17,18]. These antimicrobial properties of bioactive yeasts specifically change across genera and species and are affected by the yeast growth stage and the optimal yeast concentration [19].
The present study aimed to assess the effectiveness of 19 yeast strains belonging to five genera, isolated from natural environments, on B. cinerea. Preliminary screening was performed using an in vitro approach to select strains with a promising antimicrobial activity. The selected yeasts were evaluated in a vineyard to control the post-harvest decay of grapes before the grape harvest.
2. Materials and Methods
2.1. Yeasts and Culture Conditions
The yeasts used in the trials as BCAs against B. cinerea belonged to the Microbial Collection of the Department of Life and Environmental Sciences (DiSVA) of Polytechnic University of Marche (Italy). All yeasts reported in Table 1 were previously isolated from flowers, leaves and fruits.

Table 1.
Species and origins of the yeast strains tested as potential antifungal agents.
All yeast strains were stored on yeast extract peptone dextrose (YPD) agar (yeast extract, 1%; peptone, 2%; dextrose, 2%; agar, 1.8%) at 4 °C before use. The commercial mold B. cinerea N51 (DSMZ Germany Collection of Microorganisms and Cell Cultures GmbH, Braunschweig, Germany) was maintained and stored at 4 °C on potato dextrose agar (PDA; Oxoid, Basingstoke, UK) until use. A commercial formulation named Botector® (Manica S.p.a., Rovereto, Italy) was used as a bioactive positive control after reconstitution following the supplier’s suggestions (in 0.1% sterile water). This preparation was made up of freeze-dried cultures of two strains of Aureobasidium pullulans, as reported in the technical sheet according to Regulation (EC) No. 1907/2006.
2.2. Screening to Evaluate the Antagonistic Effects of Yeasts
Three different plate tests were set up to preliminarily determine the antimicrobial efficacy of yeasts against B. cinerea. In the first two cases, the competition for space and/or nutrients was simulated, while a contactless test was set up in the third experiment.
2.2.1. Control of B. cinerea in an Early Stage of Infection
The ability of yeasts to control B. cinerea simulating an early stage of infection was assessed through an in vitro assay based on a dual-culture method, following the procedure described by Wang et al. [3] with some modifications. Firstly, yeast strains were precultured in an YPD broth for 48 h at 25 °C, and molds were grown on PDA for 96 h at 25 °C under sunlight exposure. Yeast cells were washed twice and resuspended in sterile water to a final concentration of 107 cells/mL, which was obtained using a Thoma–Zeiss counting chamber [20]. B. cinerea spores were collected from a fresh plate with a sterile tip and diluted in a sterile 0.1% Triton X-100 solution to a final concentration of 105 spores/mL. Ten microlitres of each suspension (yeast and mold) were simultaneously spotted on the PDA surface 5 cm apart. The test was performed in triplicate, and negative control was performed by inoculating 10 µL of sterile water, instead of the yeast suspension. The positive control was a 0.1% Botector® (Manica S.p.a., Rovereto, Italy) solution adjusted to the final concentration similar to the other yeasts (107 cells/mL). The plates were incubated at 25 °C for 5–7 days. The inhibitory effect of each yeast was evaluated by measuring the radial growth of B. cinerea in millimetres compared to the growth of mold in the absence of yeast and with that of the positive control.
2.2.2. Containment of Advanced Botrytis Infection (Late)
The antagonistic activity of the same yeasts was evaluated through another in vitro assay based on a dual culture method, in which the mold was inoculated at the hyphal stage (simulation of advanced infection status). The test was carried out by streaking each yeast in a circular motion for 2 cm, starting from the edge toward the centre of the PDA plate medium. Together with a pre-incubation period of 24 h, this modality gave a double advantage to each yeast: it grew and released extracellular metabolites concentrated in the centre of the plate in the agar. After 24 h at 25 °C, 1 cm2 of B. cinerea mycelial plug (advanced development status) was placed in the centre of a Petri dish, 3 cm away from the yeast inoculum, and incubated for 5–7 days at 25 °C. Positive control was used, as described above. Three replicates were performed for each yeast/gray mold combination. The results were analysed by measuring the diameter (mm) of B. cinerea mycelial growth compared to the negative and positive controls.
2.2.3. Antifungal Effect of Volatile Organic Compounds (VOCs) of Yeasts
The possible antimold effect of VOCs produced by yeast was evaluated following the method described by Oro et al. [12]. Briefly, each yeast was seeded on YPD agar medium and incubated for 24 h at 25 °C, and then, the lid of the Petri dish was substituted with a plate upside down (face to face without any direct contact between microorganisms) containing a PDA medium where a plug of B. cinerea actively grew. The two plates were sealed with a Parafilm and incubated at 25 °C for seven days. Growth inhibition was determined by measuring the diameter of the fungal mycelial growth (mm) compared to that of the negative control carried out without yeast culture. Positive control was performed using the commercial preparation Botector® (Manica S.p.a., Rovereto, Italy), instead of the yeast culture. The test was performed in triplicate.
2.3. Extracellular Enzymatic Activities of Yeasts
All yeasts were characterised for their abilities to produce/secrete lytic enzymes with protease, esterase, amylase, β-glucosidase, cellobiose and killer activities. Protease, esterase and amylase activities were evaluated following the protocol described by Buzzini and Martini [21]. Briefly, the yeasts were spotted on a YEPG medium containing 2% casein, and a clear zone around the colony indicated the presence of the protease activity. The medium containing 1% peptone, 0.5% NaCl, 0.01% CaCl2·2H2O, 1% Tween-80 and 2% agar was used to assess the esterase activity through the presence of a visible precipitate around the yeast colony after incubation. Amylase activity was evaluated as starch hydrolysis by incubating the yeast on a medium containing soluble starch. After the yeast growth, the medium was flooded with an iodine solution, and a yellow zone around the colony indicated a starch-degrading ability. The evaluations of killer, cellobiose and β-glucosidase activities was carried out following the methods of Comitini et al. [22], Guo et al. [23] and Rosi et al. [24], respectively.
2.4. Antimicrobial Activity of Yeasts on Grape Berries
Based on the results obtained by plate assays, the six strains that showed the best antagonistic effects against B. cinerea were selected. These strains were then evaluated in harvested grape berries using two different methods. In both modalities, assays were carried out in batches (20 grapes per batch) previously sterilised in 1% sodium hypochlorite following the procedure proposed by Wang et al. [3]. In the first case, the berries of each batch were pierced with a sterile tip, causing a lesion that was lodged for the inoculation of the yeast suspensions (10 µL of 107 cells/mL), followed by 10 µL of a suspension of 2 × 105 spores/mL of B. cinerea. In the second modality, the berries were immersed in each yeast suspension; after 20 min of air drying, the B. cinerea suspension (2 × 105 spores/mL) was sprayed over the berries. These were stored in a sterile plastic bag to simulate a humid environment, incubated for 20 days at 4 °C, then transferred to 25 °C for three days and observed for B. cinerea growth.
After the two experimental preparations, the berries were monitored daily for three days to observe the damage caused by B. cinerea infection. Disease severity was recorded according to an empirical scale with six degrees: 0, healthy fruit; 1, 1–20% fruit surface infected; 2, 21–40% fruit surface infected; 3, 41–60% fruit surface infected; 4, 61–80% fruit surface infected; and 5, ≥81% fruit surface infected. The infection index or McKinney index incorporates both the incidence and severity of the disease and expresses the weighted means of the disease as a percentage of the maximum possible level [25]. Specifically, it was calculated using the following equation:
where d is the category of the rot intensity scored on the grapes, f is its frequency, N is the total number of grapes examined (healthy and rotted), and D is the highest category of the disease intensity that occurs on an empirical scale [12].
I = d × f/N × D × 100,
2.5. Evaluation of Two Selected Bioactive Yeasts by Field Experiments
Based on the results of experiments on grapes, two antagonistic yeast strains were selected. The efficacy of the two antagonists against bunch rots of wine grapes was evaluated during the 2018 harvest in a vineyard located in the centre of Italy. Untreated and Botector®-treated vines served as negative and positive controls, respectively. The two antagonistic strains were tested separately and together in a mixture (MIX) at a 1:1 volume ratio. Initially, the biomass of the two yeasts was produced in a 30 L bioreactor and harvested after ultracentrifugation at 4 °C under sterile conditions. The biomass was dissolved in sterile water and aliquoted (at a final concentration of 1 × 107 cells/mL) into three lots of 5 L with strains 1, 2 and MIX, respectively.
The experimental plots consisted of one to nine vines per treatment in the different experiments, arranged as randomised blocks with five replicates. The two bioactive strains, the MIX and the controls were applied until run-off, with a motor-driven back-sprayer, in two stages: (i) at the time of the bunch veraison; and (ii) at the time of grape maturity. After each application, representative grape samples were collected to test yeast colonisation in the laboratory through viable cell counts using WL nutrient agar (Wallerstein Laboratories, Oxoid, Hampshire, UK) with 0.02% biphenyl to prevent indigenous mold diffusion. Based on its differential properties, this media allowed us to distinguish M. pulcherrima and A. pullulans species from other indigenous yeast species even if it was not possible to distinguish native strains from those sprayed belonging to the same specie M. pulcherrima colonies appearing as red colonies, whereas A. pullulans appeared as a “branched star” colony. At the harvest time, according to the farmer, all treated bunches were not collected to evaluate the antagonistic effect of yeasts sprayed by monitoring viable plate counts until the 10th day. Natural infection with B. cinerea was expected.
2.6. Statistical Analyses
The data were analysed using one-way analysis of variance (ANOVA), followed by Duncan’s test (p < 0.05). The percentage data were arcsine-transformed before analysis to improve the homogeneity of variance, when the range of percentages was >40%. The actual values were also presented. All trials were repeated at least twice with at least three replicates. Data from two or more experiments were pooled, and statistical analysis to determine the homogeneity of variances was performed using Levene’s test.
3. Results
3.1. In Vitro Antagonistic Effect of Potential BCAs
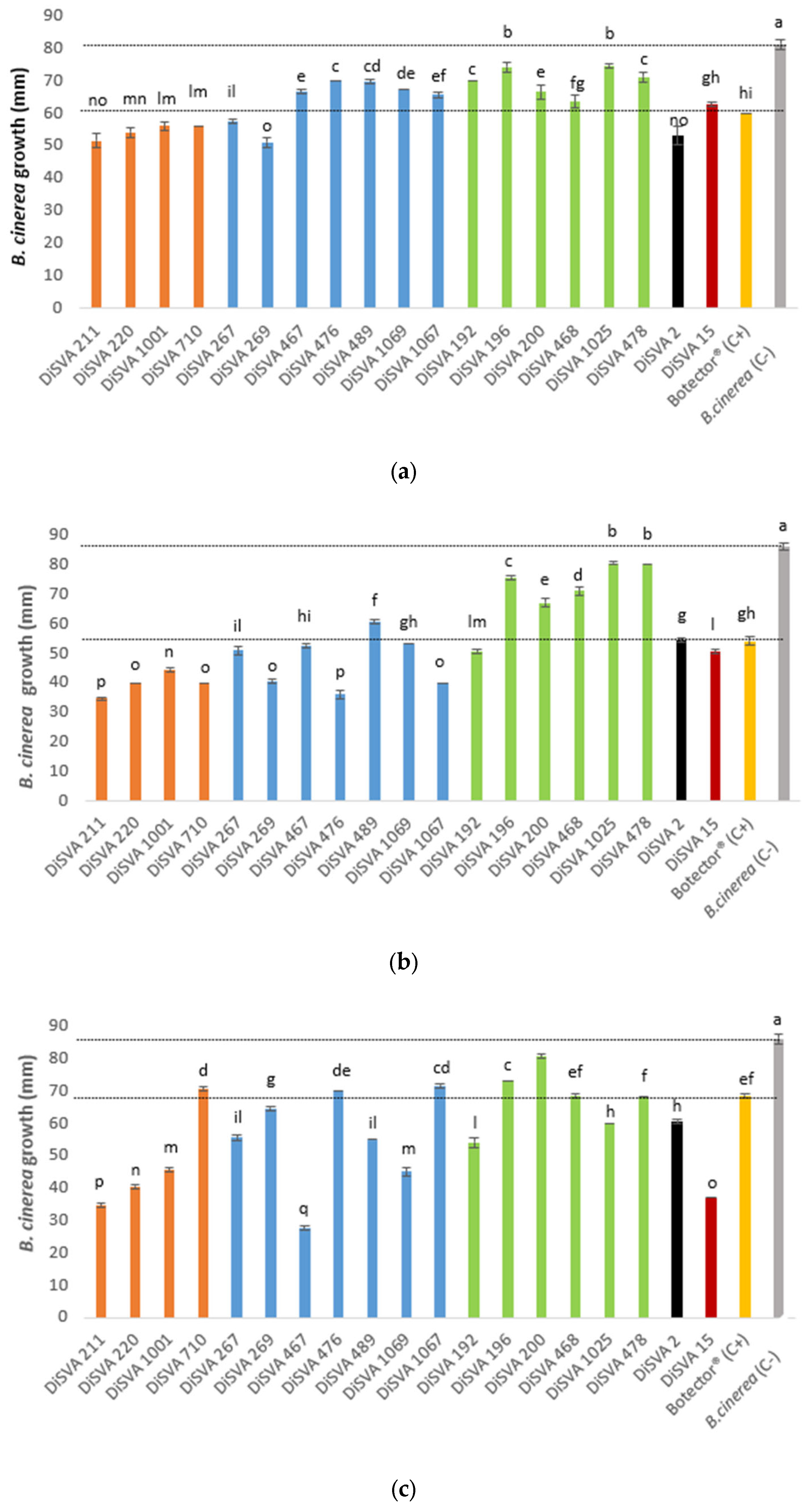
The abilities of the 19 potential BCAs were preliminarily tested in three plate assays. First, the possibility of yeasts carrying out a control action at the early stage of B. cinerea infection was tested using two co-inoculated suspensions of yeast and mold spores. The results of this assay are shown in Figure 1a.
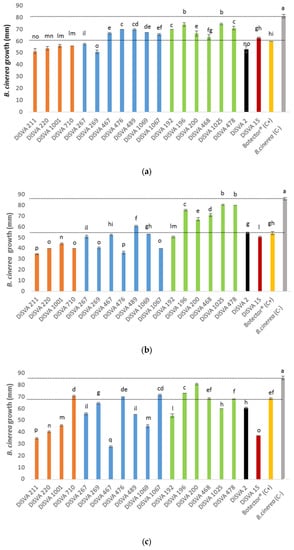
Figure 1.
Antagonistic activity of yeasts against Botrytis cinerea (B. cinerea) growth compared with the negative and positive controls: (a) trials carried out considering an early stage of mold infection; (b) trials carried out considering an advanced stage of mold infection. (c) yeast’s volatile organic compounds (VOCs) antimicrobial activity on B. cinerea infection. The dotted lines highlighted the positive and negative control values, and the different superscript letters (a, b, c, d, e, f, g, h, I, l, m, n, o, p) among the bars are significantly different, according to Duncan’s test (p < 0.05).
All strains tested showed a significant ability to control the development of mold than the negative control, in which a drop of sterile water was placed in the plate. However, different degrees of containment were observed. In particular, all A. pullulans, M. pulcherrima DiSVA 267 and DiSVA 269 strains, W. anomalus DiSVA 2 and Kluyveromyces wickerhamii DiSVA 15 showed a reduction in the mold growth of approximately 30 mm when compared with the negative control (hyphae radial growth of negative control = 80 mm), and an additional 10 mm of reduction compared with the positive control Botector® (hyphae radial growth = 60 mm).
Similarly, all potential BCAs showed the ability to inhibit the growth of B. cinerea in an advanced stage of infection (simulated with an inoculum of a plug with germinated mold) compared to the negative control, which showed a hyphal growth of approximately 85 mm (Figure 1b). In this thesis, the positive control reduced the mold hyphae growth by approximately 30 mm and all A. pullulans, all M. pulcherrima (apart from DiSVA 489 strain), W. anomalus DiSVA 2, K. wickerhamii DiSVA 15 and only C. albidus DiSVA 192 showed a better or equal reduction in B. cinerea hyphal growth. In particular, A. pullulans DiSVA 211, M. pulcherrima DiSVA 269 and DiSVA 1067 reduced the mold growth by approximately 50 mm.
Finally, Figure 1c shows the results of the test that assayed the possible mechanism of the control mediated by VOC production. In this test, the positive control reduced the mold growth by 15 mm compared with the negative control, while almost all potential BCA strains tested showed better results than the positive control (Botector®). A. pullulans DiSVA 211 and DiSVA 220, M. pulcherrima DiSVA 467 and K. wickerhamii DiSVA 15 allowed the development of approximately 40 mm of B. cinerea with a reduction of approximately 50 mm in comparison with the negative control and an additional growth reduction of approximately 30 mm in comparison with the positive control Botector®.
3.2. Extracellular Enzymatic Activities of Potential BCAs
The 19 yeast and yeast-like strains were also evaluated for extracellular lytic enzyme production and killer activity, and the results are reported in Table 2.
Table 2.
Enzymatic characterisation of potential bioactive yeasts.
All A. pullulans strains exhibited protease, β-glucosidase, amylase, cellobiose and esterase activities, except DiSVA 220 and DiSVA 710 strains that did not show a protease activity. In contrast, none of the A. pullulans strains exhibited a killer ability against the sensitive yeast. A greater variability was observed among the M. pulcherrima strains. All strains produced cellobiosidase enzymes but did not show killer and amylase activities. Only M. pulcherrima DiSVA 269 also exhibited an esterase activity, while DiSVA 269 and DiSVA 476 showed a β-glucosidase activity, and DiSVA 467 produced protease lytic enzymes. All the Cryptococcus strains exhibited amylase, cellobiose (except DiSVA 196) and esterase activities. Protease was detected only by DiSVA 192, DiSVA 196 and DiSVA 468, whereas no β-glucosidase and killer activities were observed. K. wickerhamii DiSVA 15 showed only cellobiose and killer activities, whereas W. anomalus DiSVA 2 also produced esterase enzymes.
3.3. Evaluation of the BCAs Effectiveness on Grapes
From a comparative analysis of the results obtained in the three plate tests (Figure 1a–c), the strains showing a significant control action superior to that of Botector® were selected and subsequently tested in the harvested berries in the laboratory. Seven strains of A. pullulans, DiSVA 211, DiSVA 220, DiSVA 1001, M. pulcherrima strains DiSVA 267 and DiSVA 269 and W. anomalus DiSVA 2 were evaluated.
To be used as a BCA in post-harvest applications, microorganisms should not have phytotoxic effects besides being effective against post-harvest pathogens. Artificially wounded berries treated with yeast did not show any evidence of phytotoxic effects for 25 days, because the wound size was not different from that of the negative control (data not shown). In addition, fruit sprayed with a yeast solution or inoculated in a berry wound, cold-stored and then exposed to shelf life did not show any phytotoxic effect. Thus, the pre-requisite for the possible consideration of these three strains of biocontrol agents was fulfilled. After the post-harvest treatment of lesioned berries, the damage caused by B. cinerea on the berries and the relative effectiveness of the seven selected BACs were evaluated based on the percentage of decay (Table 3) and the McKinney index (Figure 2).
Table 3.
Decay percentage (%) and relative rank analyses of the six potential bioactive yeasts on lesioned grape berries.
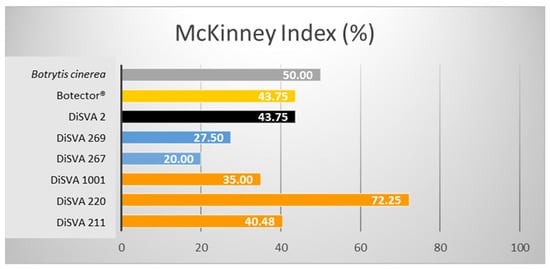
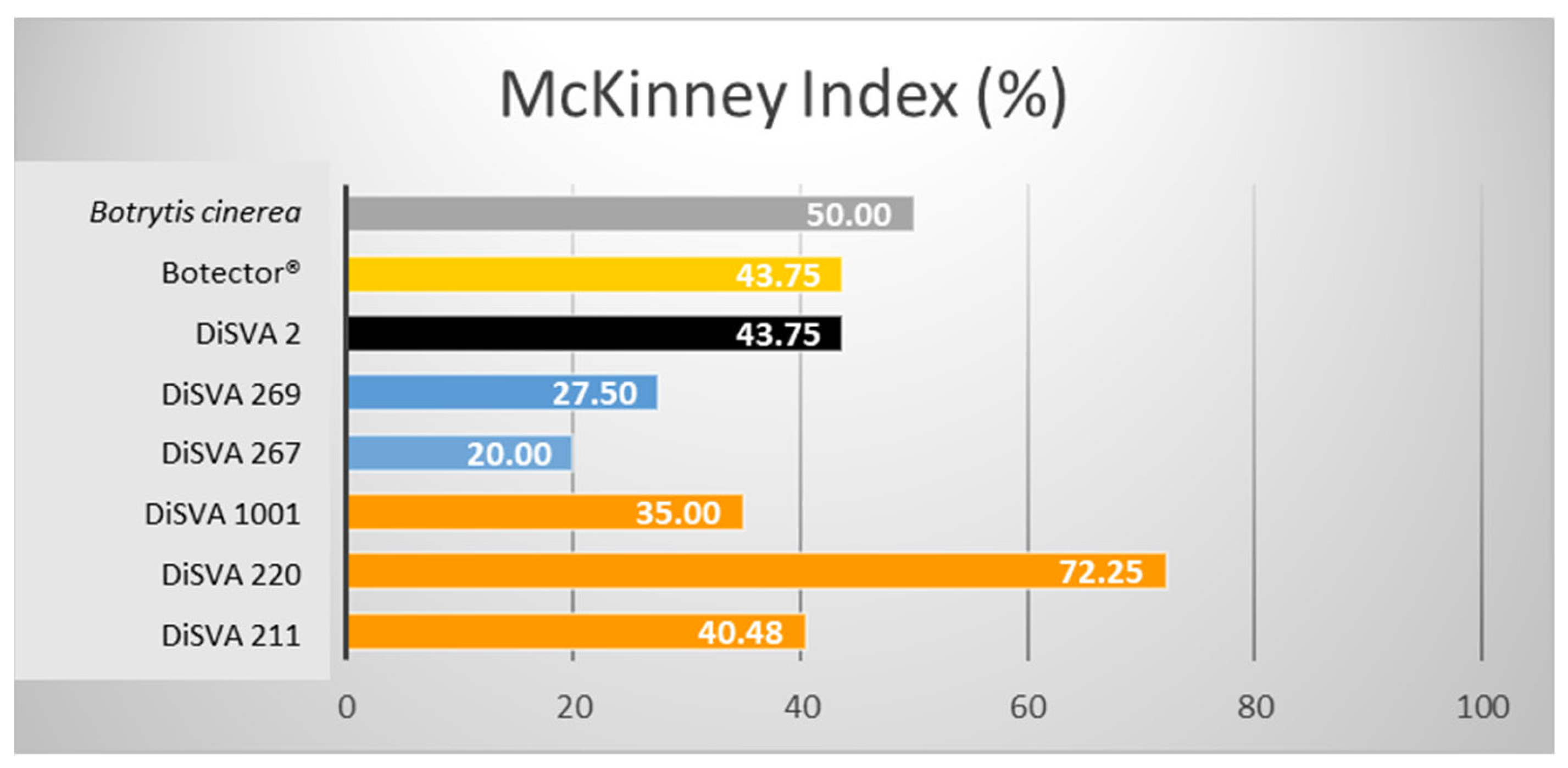
Figure 2.
McKinney indices (%) of the six potential bioactive yeasts on lesioned grape berries.
All strains effectively reduced the gray rot in grape berries during storage after post-harvest treatments. Lesioned fruits showed a significantly reduced decay compared with untreated controls, with higher effectiveness than Botector®. Indeed, after a rank analysis, all the strains were placed in a more favourable position than the Botector® (rank 6), except for W. anomalus DiSVA 2, which seemed to be the least effective (rank 7). A. pullulans DiSVA 211 proved to be the most effective in controlling the damage spread, placing it in the first rank.
A reduction in the McKinney index was evident in all strains compared to in the control. These antimicrobial action results were well correlated with each other: the two strains of M. pulcherrima and A. pullulans DiSVA 211 showed a more significant damage reduction and, therefore, a greater efficacy. Again, the W. anomalus strain DiSVA 2 showed the lowest control efficacy.
Similar results were obtained from the same experiment on berries not lesioned but sprayed with a suspension of the same BCAs (Table 4 and Figure 3).
Table 4.
Decay (%) and relative rank analyses of the six potential bioactive yeasts on surface-infected grape berries. The B. cinerea growth was observed after three days at 25 °C.
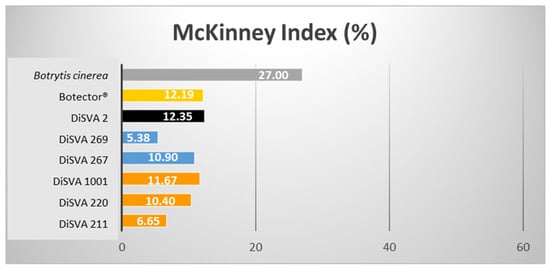
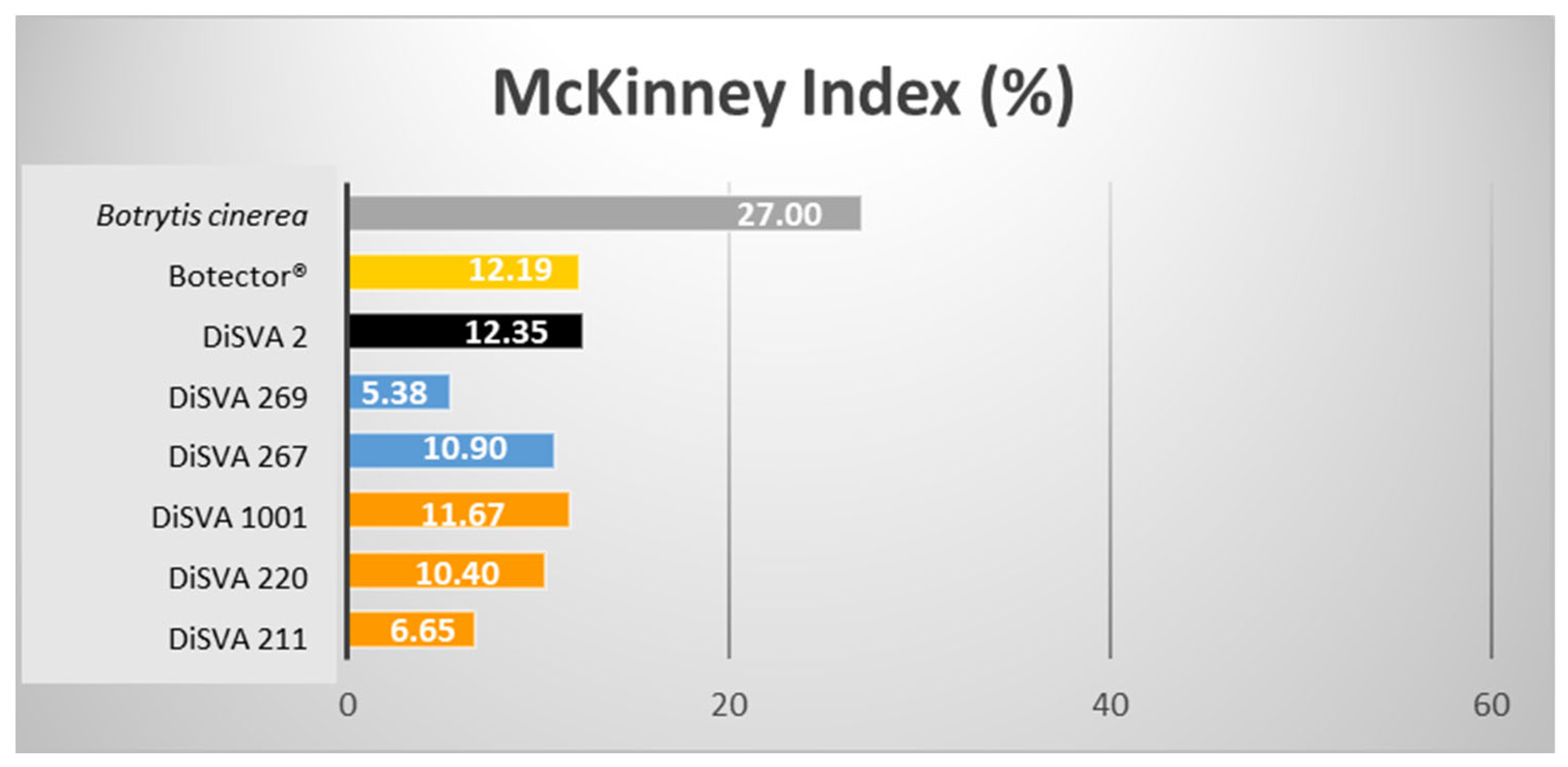
Figure 3.
McKinney indices (%) of the six potential bioactive yeasts on surface-sprayed harvested grape berries.
In this case, the best decay control was achieved by M. pulcherrima strain DiSVA 269 (rank 1 in the decay control). This strain confirmed its best behaviour showing a McKinney index of 5.38%, followed by A. pullulans strain DiSVA 211 with a McKinney index of 6.65%. As expected, the colonisation level of B. cinerea in damaged grapes was higher (McKinney index of 50%) than that in sprayed grapes (McKinney index of 27%). Consequently, the abilities of these selected BCAs to control gray mold were also lower in unpunctured grapes but not less effective.
3.4. Application of M. pulcherrima DiSVA 269 and A. pullulans DiSVA 211 in the Field
After a comprehensive analysis of all the results obtained, the strains which were most effective in controlling the development of B. cinerea in laboratory tests (both in plates and on grapes) were M. pulcherrima DiSVA 269 and A. pullulans DiSVA 211. Therefore, these two strains were used in the field during the pre-harvest treatment. In the first evaluation, after each field treatment (both at the veraison time and at the ripening of the bunches), the real colonisation of the two strains, used both separately and together in the MIX, was evaluated. The results reported in Table 5 showed the actual and stable colonisation of both yeasts for each treatment.
Table 5.
Evaluation of the colonization of the potential biological control agents (BCAs) on the grapes after each treatment and their persistence. Values are reported as total microflora counts, while the values inside the brackets represented the A. pullulans DiSVA 211 (Ap)- and M. pulcherrima DiSVA 269 (Mp)-inoculated species without the distinction between indigenous and inoculated strains. Data are represented as means ± standard deviations.
Ten days after treatment, the persistence results confirmed that both yeasts maintained the same viability level (Table 5).
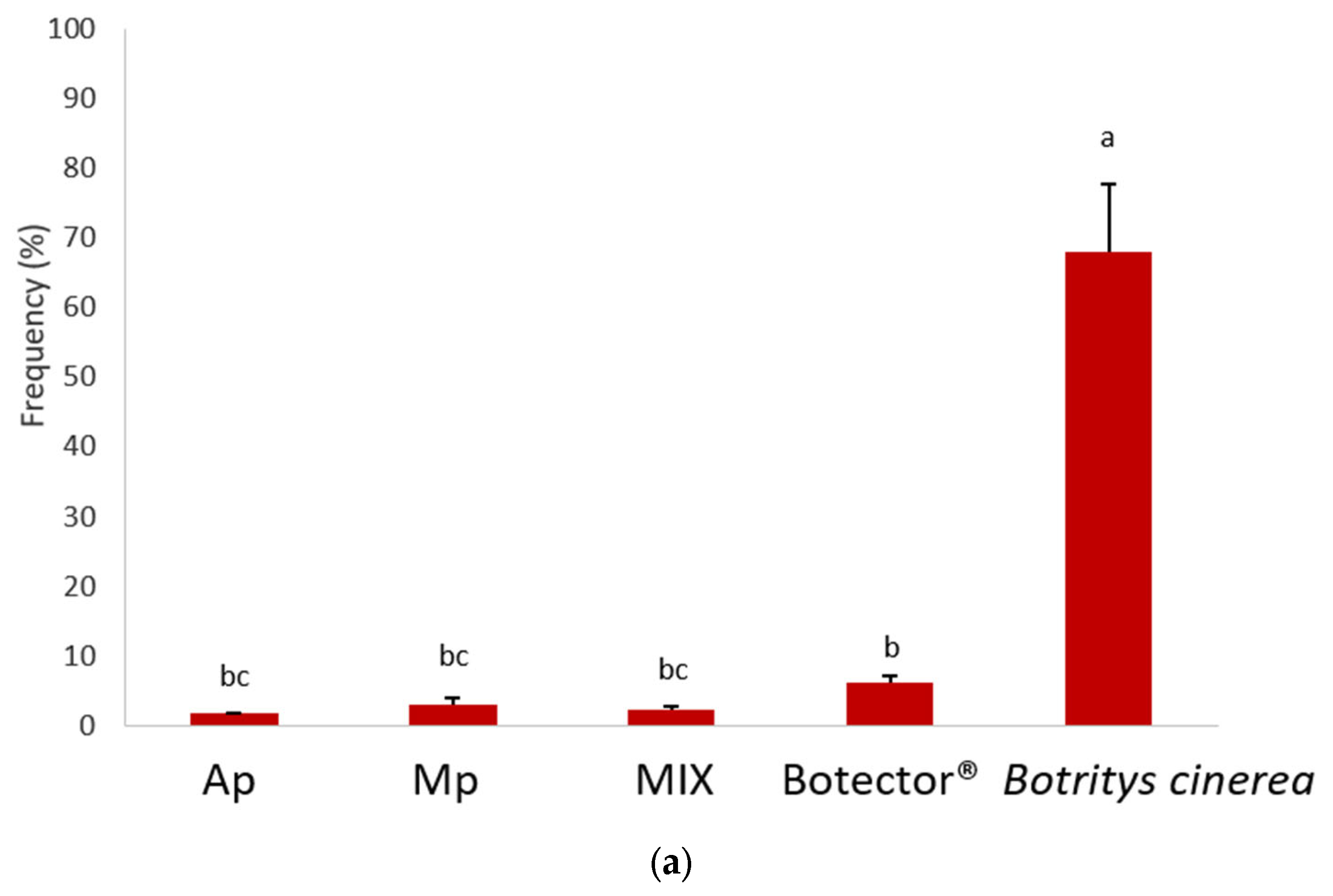
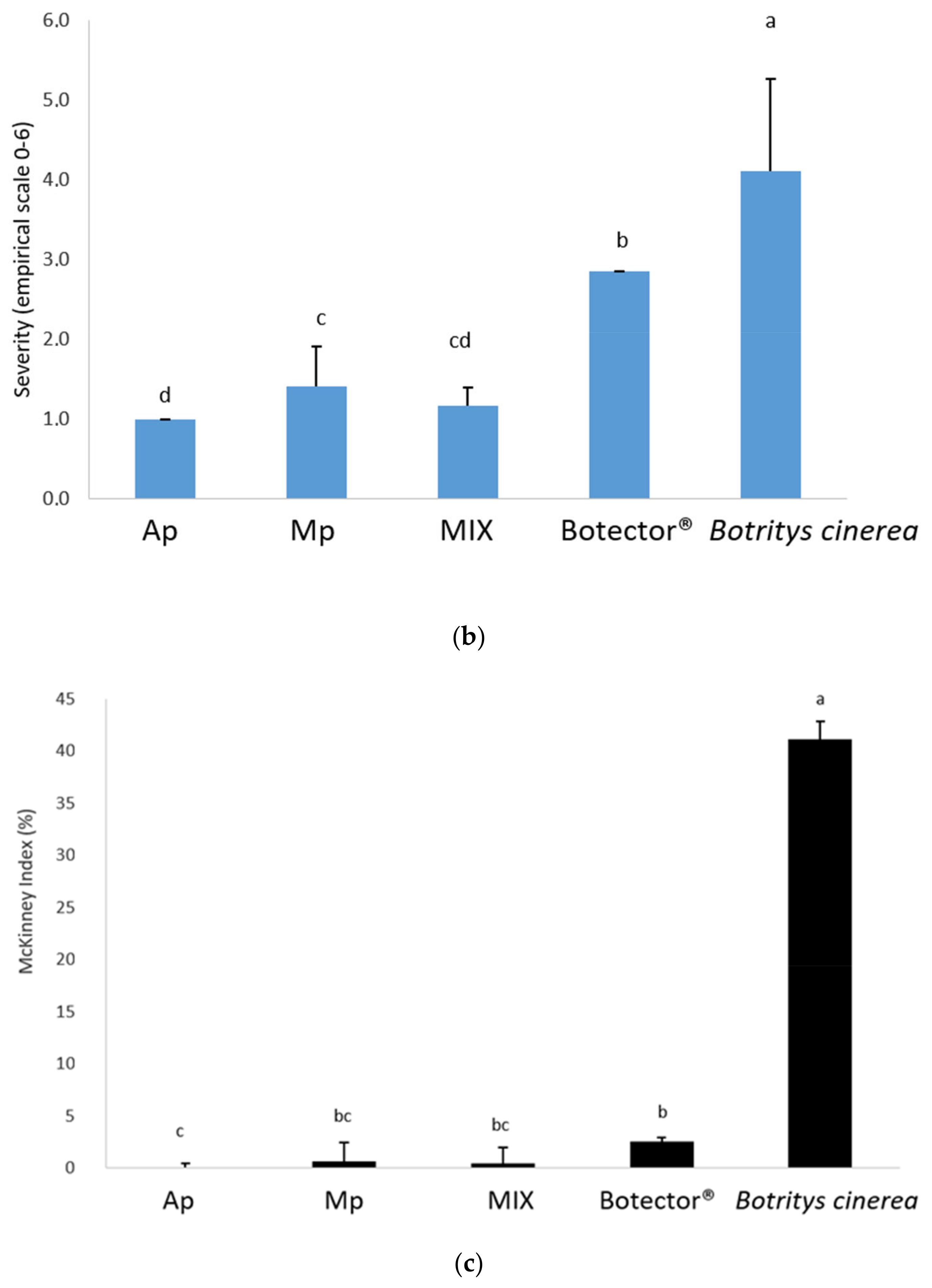
The effectiveness of post-harvest treatments (shelf life at room temperature) was evaluated by harvesting the berries 24 h after the second treatment and 10 days later. The growth of B. cinerea was evaluated by the statistical analysis of the randomised block frequency, the severity and the McKinney index (Figure 4). The results showed that B. cinerea grew on most of the berries monitored in the untreated samples.
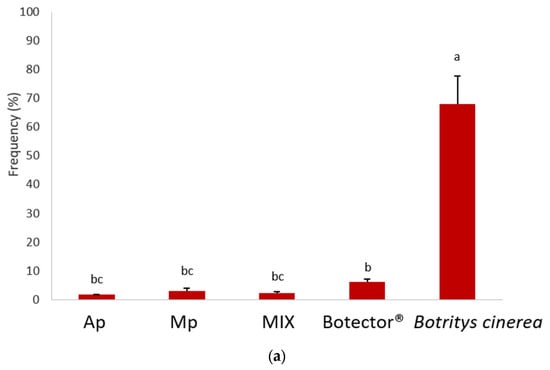
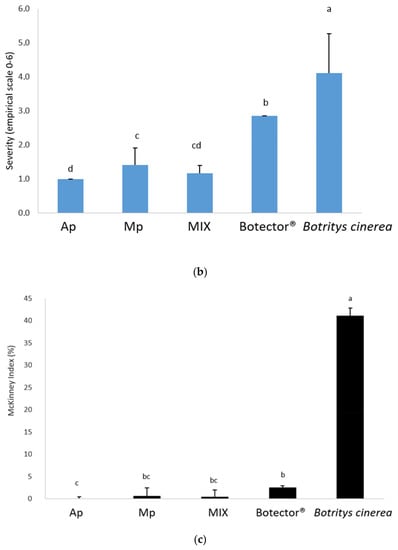
Figure 4.
Frequency (a), decay (b) and McKinney’s index (c) of gray mold on grapes. Different superscript letters (a, b, c, d) show significant differences according to Duncan’s test (p < 0.05).
The thesis treated with Botector® showed good mold control, although with nonhomogeneous data in the various samples. In the thesis Mp, M. pulcherrima DiSVA 269 showed a very low incidence of damage, but in berries presumably not colonised by BCA, B. cinerea grew considerably. A. pullulans strain DiSVA 211 showed a lower control of mold decay, but with more homogeneous results for all samples.
Finally, the MIX thesis exhibited a higher control capacity, combining the positive aspects of the two yeasts obtained separately. Indeed, the severity and the McKinney index were significantly lower in the latter case.
4. Discussion
Currently, there is increasing interest in using BCAs to suppress bunch rot caused by B. cinerea in grapes. Different filamentous fungi, bacteria and yeasts have been selected as potential biological suppressors of this pathogen [26,27]. Many of these suppressors suppress the growth of plant pathogens through competition for nutrients or by the production of inhibitory metabolites and/or parasitism. Although several small-scale studies have been carried out highlighting different antagonist strains, and for some of them, the mechanisms of action have been understood and the practical application of BCAs remains an unsolved problem. This difficulty is probably linked to the commercialisation of BCAs, which require a multistep process that involves the isolation and screening of potential antagonism, testing the efficiency of the isolate in the field, biomass production, formulation, toxicity studies, delivery, compatibility, registration and release [28]. However, the evidence that in vivo studies on antagonistic yeasts play efficient control actions in a large variety of vegetal matrices, including grapes, prompted us to research this area.
In this study, 19 strains, previously characterised as antagonists, were evaluated for their potential bio-preservative effects against B. cinerea in wine grapes.
Preliminary findings, carried out in plate assays, indicated that M. pulcherrima, W. anomalus and A. pullulans are yeast species involved in gray rot disease control, supporting previous results [3,4,6,29,30]. The antimicrobial activity of the two yeasts revealed under laboratory conditions, together with the results of the colonisation and persistence in a vineyard, indicates that the control of disease (diffusion and severity) is due to sprayed yeasts on grapes.
In this regard, several mechanisms of action are suggested. M. pulcherrima strain MPR3, isolated from spontaneous olive fermentation, has been demonstrated to have a great antifungal activity against B. cinerea, mediated by VOC production [31], while other strains appear to effectively control gray rot with other mechanisms associated with iron depletion [29,32]. Although the mechanism by which yeasts inhibit the development of B. cinerea has not been studied here, our preliminary results suggested that M. pulcherrima DiSVA 269 exerts its inhibitory action in three possible ways. The first could be the competition for nutrients. Indeed, it colonises grape berries and persists on grape berries for at least 10 days after treatment.
The second strategy could be the presence of β-glucosidase activity, which has been well studied as a potential cause of fungal wall damage [33]. In particular, after enzymatic lytic evaluation, M. pulcherrima strain DiSVA 269 was selected to be more effective in controlling B. cinerea. The third way M. pulcherrima strain DiSVA 269 controls gray rot could be due to pulcherriminic acid production that depletes iron present in the environment, making it unavailable to other microorganisms, as previously reported in an in vitro study [34].
Multiple modes of action of A. pullulans have been reported to explain its biocontrol efficacy, including competition for nutrients and space, production of cell wall-degrading enzymes, synthesis of antifungal compounds and mycoparasitism [2]. In this study, the evaluation of the principal enzymatic activities showed that all strains of A. pullulans tested were positive for all plate tests. This trend confirmed the ability of this species to produce and secrete different lytic enzymes [35].
A. pullulans is also known for inhibiting mold development by competing for space [2,35,36]. The results showed the ability to persist on the surface of the grapes for at least 10 days after treatment, and in the case of damaged grapes, A. pullulans DiSVA 211 showed the lowest decay percentage. The mechanism of action could be due to the expansion of this yeast-like structure in the wound, limiting the colonisation and development of B. cinerea. This inhibitory action can also be reinforced through VOC production, as reported by Don et al. [37]. On the other hand, A. pullulans is known to exhibit a high genotypic diversity [2]; therefore, the choice of strains to be used as biocontrol agents is a critical step. Indeed, among the four A. pullulans strains tested here, only two were more effective in plate tests, and only DiSVA 211 confirmed their significant efficacies in vivo during field treatments. Despite the availability of A. pullulans-based products currently marketed, such as Botector® or BIO-FERM [38,39,40], new candidates to control gray rot in vineyards are required. In this regard, the results of field treatments indicated that several factors should be evaluated, such as the fast colonisation of potential BCAs and their persistence, which are characteristic of M. pulcherrima DiSVA 269 and A. pullulans DiSVA 211, selected as the best potential BCAs. In this regard, the MIX trial showed promising results in gray mold inhibition, expressed as the frequency, the severity and the McKinney index, probably due to the synergistic action of competition for space and nutrients, specific enzymatic activities and production of VOCs exerted by both species.
Further investigations of the anti-B. cinerea by two selected yeasts, in pure and mixed applications and under different agronomical and environmental conditions, are needed to constitute a promising source of knowledge and to set up strategies to prevent or reduce harvested commodity damage.
5. Conclusions
In this study, A. pullulans and M. pulcherrima were the most promising potential BCAs for the development of B. cinerea mold. The trials carried out in vineyards showed their anti-B. cinerea action and could be proposed as a single species or in combination by exploiting the synergistic action of their antagonistic capacities through the rapid colonisation of the grapes and persistence on the grape surface. This approach meets consumer expectations and widely accepts the development of bio-based applications to exert microbial control in agro-food chains, according to eco-friendly approaches and products free of synthetic chemicals. For these purposes, the approach proposed here could be further investigated to understand the mechanisms of action and evaluate the safety of the proposed bioactive yeasts.
Author Contributions
A.A., L.C., T.P., G.R., M.C. and F.C. contributed equally to the design and discussion of the manuscript. A.A., L.C. and T.P. carried out the experimental part of the work. A.A., L.C., T.P., G.R., M.C. and F.C. carried out the analysis of the data and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research is a part of the project SYSTEMIC “An integrated approach to the challenge of sustainable food systems: adaptive and mitigatory strategies to address climate change and malnutrition”, knowledge hub on Nutrition and Food Security, has received funding from national re-search funding parties in Belgium (FWO), France (INRA), Germany (BLE), Italy (MIPAAF), Latvia (IZM), Norway (RCN), Portugal (FCT) and Spain (AEI) in a joint action of JPI HDHL, JPI-OCEANS and FACCE-JPI launched in 2019 under the ERA-NET ERA-HDHL (n° 696295).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Steel, C.C.; Blackman, J.W.; Schmidtke, L.M. Grapevine bunch rots: Impacts on wine composition, quality, and potential procedures for the removal of wine faults. J. Agric. Food Chem. 2013, 61, 5189–5206. [Google Scholar] [CrossRef] [PubMed]
- Bozoudi, D.; Tsaltas, D. The multiple and versatile roles of Aureobasidium pullulans in the vitivinicultural sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Romanazzi, G.; Smilanick, J.L.; Feliziani, E.; Droby, S. Integrated management of postharvest gray mold on fruit crops. Postharvest Biol. Technol. 2016, 113, 69–76. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Simone, N.; Pace, B.; Grieco, F.; Chimienti, M.; Tyibilika, V.; Santoro, V.; Capozzi, V.; Colelli, G.; Spano, G.; Russo, P. Botrytis cinerea and table grapes: A review of the main physical, chemical, and bio-based control treatments in post-harvest. Foods 2020, 9, 1138. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Biocontrol ability and action mechanism of food-isolated yeast strains against Botrytis cinerea causing postharvest bunch rot of table grape. Food Microbiol. 2015, 47, 85–92. [Google Scholar] [CrossRef]
- Lemos Junior, W.J.F.; Bovo, B.; Nadai, C.; Crosato, G.; Carlot, M.; Favaron, F.; Giacomini, A.; Corich, V. Biocontrol ability and action mechanism of Starmerella bacillaris (synonym Candida zemplinina) isolated from wine musts against gray mold disease agent Botrytis cinerea on grape and their effects on alcoholic fermentation. Front. Microbiol. 2016, 7, 1249. [Google Scholar] [CrossRef]
- Linares-Morales, J.R.; Gutiérrez-Méndez, N.; Rivera-Chavira, B.E.; Pérez-Vega, S.B.; Nevárez-Moorillón, G.V. Biocontrol processes in fruits and fresh produce, the use of lactic acid bacteria as a sustainable option. Front. Sustain. Food Syst. 2018, 2, 50. [Google Scholar] [CrossRef] [Green Version]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- Nicot, P.C.; Stewart, A.; Bardin, M.; Elad, Y. Biological control and biopesticide suppression of Botrytis-incited diseases. In Botrytis—The Fungus, the Pathogen and Its Management in Agricultural Systems; Fillinger, S., Elad, Y., Eds.; Springer International Publishing: Basel, Switzerlad, 2016; pp. 165–188. [Google Scholar]
- Dukare, A.S.; Paul, S.; Nambi, V.E.; Gupta, R.K.; Singh, R.; Sharma, K.; Vishwakarma, R.K. Exploitation of microbial antagonists for the control of postharvest diseases of fruits: A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1498–1513. [Google Scholar] [CrossRef]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Volatile organic compounds from Wickerhamomyces anomalus, Metschnikowia pulcherrima and Saccharomyces cerevisiae inhibit growth of decay causing fungi and control postharvest diseases of strawberries. Int. J. Food Microbiol. 2018, 265, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Montiel, L.G.; Gutierrez-Perez, E.D.; Murillo-Amador, B.; Vero, S.; Chiquito-Contreras, R.G.; Rincon-Enriquez, G. Mechanisms employed by Debaryomyces hansenii in biological control of anthracnose disease on papaya fruit. Postharvest Biol. Technol. 2018, 139, 31–37. [Google Scholar] [CrossRef]
- Roberti, R.; Di Francesco, A.; Innocenti, G.; Mari, M. Potential for biocontrol of Pleurotus ostreatus green mould disease by Aureobasidium pullulans De Bary (Arnaud). Biol. Control 2019, 135, 9–15. [Google Scholar] [CrossRef]
- Guo, J.; Fang, W.; Lu, H.; Zhu, R.; Lu, L.; Zheng, X.; Yu, T. Inhibition of green mold disease in mandarins by preventive applications of methyl jasmonate and antagonistic yeast Cryptococcus laurentii. Postharvest Biol. Technol. 2014, 88, 72–78. [Google Scholar] [CrossRef]
- Li, Q.; Li, C.; Li, P.; Zhang, H.; Zhang, X.; Zheng, X.; Yang, Q.; Apaliya, M.T.; Boateng, N.A.S.; Sun, Y. The biocontrol effect of Sporidiobolus pararoseus Y16 against postharvest diseases in table grapes caused by Aspergillus niger and the possible mechanisms involved. Biol. Control 2017, 113, 18–25. [Google Scholar] [CrossRef]
- Chan, Z.L.; Tian, S.P. Interaction of antagonistic yeast against postharvest pathogens of apple fruit and possible mode of action. Postha. Biol. Technol. 2005, 36, 215–223. [Google Scholar] [CrossRef]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit, The importance of elucidating the mechanisms of action of yeast Antagonists. Tre. Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Martins, S.; Mussatto, S.I.; Martínez-Avila, G.; Montañez-Saenz, J.; Aguilar, C.N.; Teixeira, J.A. Bioactive phenolic compounds: Production and extraction by solid-state fermentation. A review. Biotechnol. Adv. 2011, 29, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Casagrande Pierantoni, D.; Corte, L.; Roscini, L.; Cardinali, G. High-Throughput rapid and inexpensive assay for quantitative determination of low cell-density yeast cultures. Microorganisms 2019, 7, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzzini, P.; Martini, A. Extracellular enzymatic activity profiles in yeast and yeast-like strains isolated from tropical environments. J. Appl. Microbiol. 2002, 93, 1020–1025. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Agarbati, A.; Canonico, L.; Galli, E.; Ciani, M. Purification and characterization of WA18, a new mycocin produced by Wickerhamomyces anomalus active in wine against Brettanomyces bruxellensis spoilage yeasts. Microorganisms 2021, 9, 56. [Google Scholar] [CrossRef]
- Guo, Z.; Duquesne, S.; Bozonnet, S.; Cioci, G.; Nicaud, J.M.; Marty, A.; O’Donohue, M.J. Development of cellobiose-degrading ability in Yarrowia lipolytica strain by overexpression of endogenous genes. Biotechnol. Biofuels 2015, 8, 109. [Google Scholar] [CrossRef]
- Rosi, I.; Vinella, M.; Domizio, P. Characterization of β-glucosidase activity in yeasts of oenological origin. J. Appl. Bacteriol. 1994, 77, 519–527. [Google Scholar] [CrossRef]
- McKinney, H.H. Influence of soil temperature and moisture on infection of wheat seedlings by Helmintosporium sativum. J. Agric. Res. 1923, 26, 195–218. [Google Scholar]
- Elmer, P.A.G.; Reglinski, T. Biosuppression of Botrytis cinerea in grapes. Plant. Pathol. 2006, 55, 155–177. [Google Scholar] [CrossRef]
- Domingues, A.; Roberto, S.; Ahmed, S.; Shahab, M.; José Chaves Junior, O.; Sumida, C.; de Souza, R. Postharvest techniques to prevent the incidence of Botrytis mold of ‘BRS Vitoria’ seedless grape under cold storage. Horticulturae 2018, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Junaid, J.M.; Dar, N.A.; Bhat, T.A.; Bhat, A.H.; Bhat, M.A. Commercial biocontrol agents and their mechanism of action in the management of plant pathogens. Int. J. Mod. Plant. Anim. Sci. 2013, 1, 39–57. [Google Scholar]
- Saravanakumar, D.; Ciavorella, A.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Metschnikowia pulcherrima strain MACH1 outcompetes Botrytis cinerea, Alternaria alternate and Penicillium expansum in apples through iron depletion. Postharvest Biol. Technol. 2008, 49, 121–128. [Google Scholar] [CrossRef]
- Raspor, P.; Miklič-Milek, D.; Avbelj, M.; Čadež, N. Biocontrol of grey mould disease on grape caused by Botrytis cinerea with autochthonous wine yeasts. Food Technol. Biotechnol. 2010, 48, 336–343. [Google Scholar]
- Parafati, L.; Vitale, A.; Polizzi, G.; Restuccia, C.; Cirvilleri, G. Understanding the mechanism of biological control of postharvest phytopathogenic moulds promoted by food isolated yeasts. Acta Hortic. 2016, 93–110. [Google Scholar] [CrossRef]
- Sipiczki, M. Metschnikowia strains isolated from botrytised grapes antagonize fungal and bacterial growth by iron depletion. Appl. Environ. Microbiol. 2006, 72, 6716–6724. [Google Scholar] [CrossRef] [Green Version]
- Khalili, E.; Javed, M.A.; Huyop, F.; Wahab, R.A. Efficacy and cost study of green fungicide formulated from crude beta-glucosidase. Int. J. Environ. Sci. Technol. 2019, 16, 4503–4518. [Google Scholar] [CrossRef]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. App. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Agirman, B.; Erten, H. Biocontrol ability and action mechanisms of Aureobasidium pullulans GE17 and Meyerozyma guilliermondii KL3 against Penicillium digitatum DSM2750 and Penicillium expansum DSM62841 causing postharvest diseases. Yeast 2020, 37, 437–448. [Google Scholar] [CrossRef]
- Di Francesco, A.; Ugolini, L.; D’Aquino, S.; Pagnotta, E.; Mari, M. Biocontrol of Monilinia laxa by Aureobasidium pullulans strains: Insights on competition for nutrients and space. Int. J. Food Microbiol. 2017, 248, 32–38. [Google Scholar] [CrossRef]
- Don, S.M.Y.; Schmidtke, L.M.; Gambetta, J.M.; Steel, C.C. Volatile organic compounds produced by Aureobasidium pullulans induce electrolyte loss and oxidative stress in Botrytis cinerea and Alternaria alternata. Res. Microbiol. 2021, 172, 103788. [Google Scholar]
- Rathnayake, R.M.S.P.; Savocchia, S.; Schmidtke, L.M.; Steel, C.C. Characterisation of Aureobasidium pullulans isolates from Vitis vinifera and potential biocontrol activity for the management of bitter rot of grapes. Eur. J. Plant Pathol. 2018, 15, 593–611. [Google Scholar] [CrossRef]
- Schilder, A. Botector: A New Biofungicide for Control of Botrytis Bunch Rot in Grapes; Department of Plant, Soil and Microbial Sciences, Michigan State University Extension. 21 August 2013. Available online: https://www.canr.msu.edu/news/botector_a_new_biofungicide_for_control_of_botrytis_bunch_rot_in_grapes (accessed on 29 July 2021).
- Romanazzi, G.; Sanzani, S.M.; Bi, Y.; Tian, S.; Gutierrez-Martinez, P.; Alkan, N. Induced resistance to control postharvest decay of fruit and vegetables. Postharvest Biol. Technol. 2016, 122, 82–94. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).