
Biocontrol Potential of Chitin and Chitosan Extracted from Black Soldier Fly Pupal Exuviae against Bacterial Wilt of Tomato


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Sample Collection and Isolation of Ralstonia solanacearum
2.3. Cultural Characterization
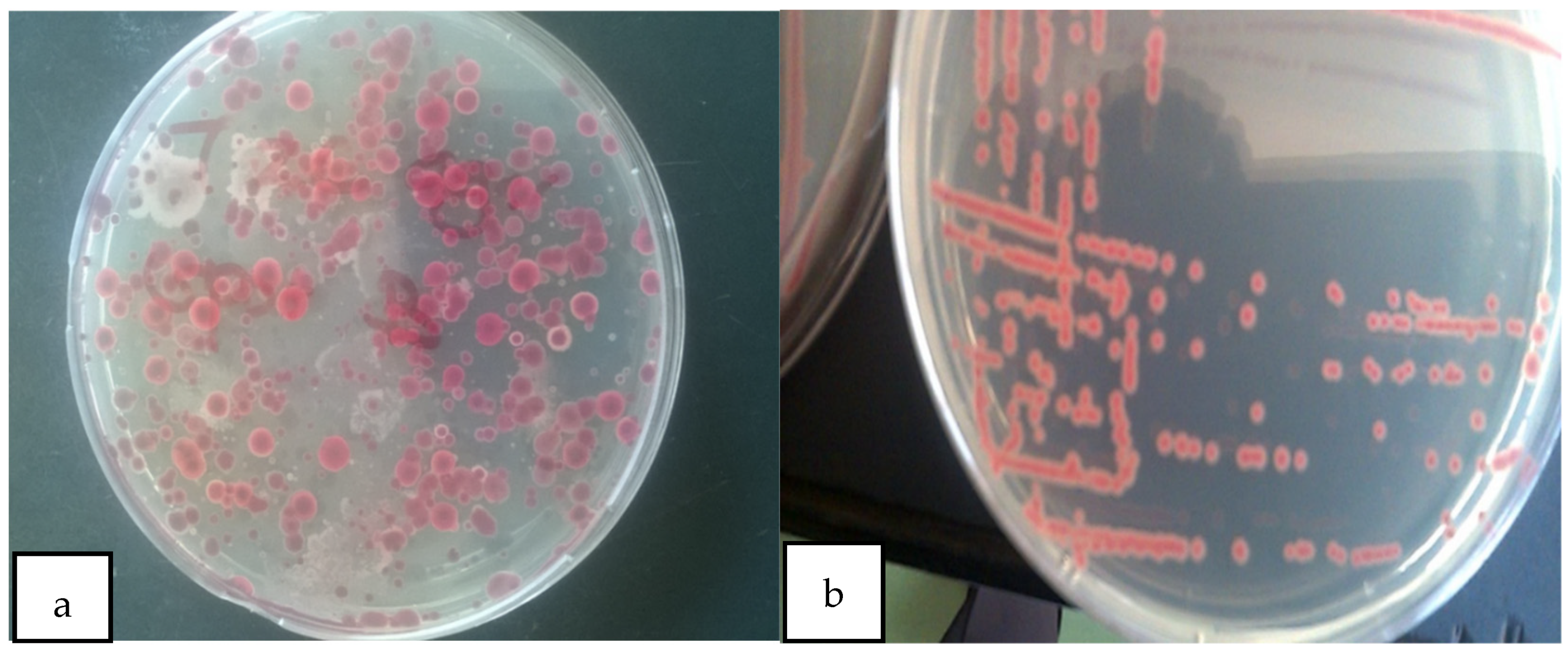
2.3.1. Morphological Characterization
2.3.2. Biochemical Characterization
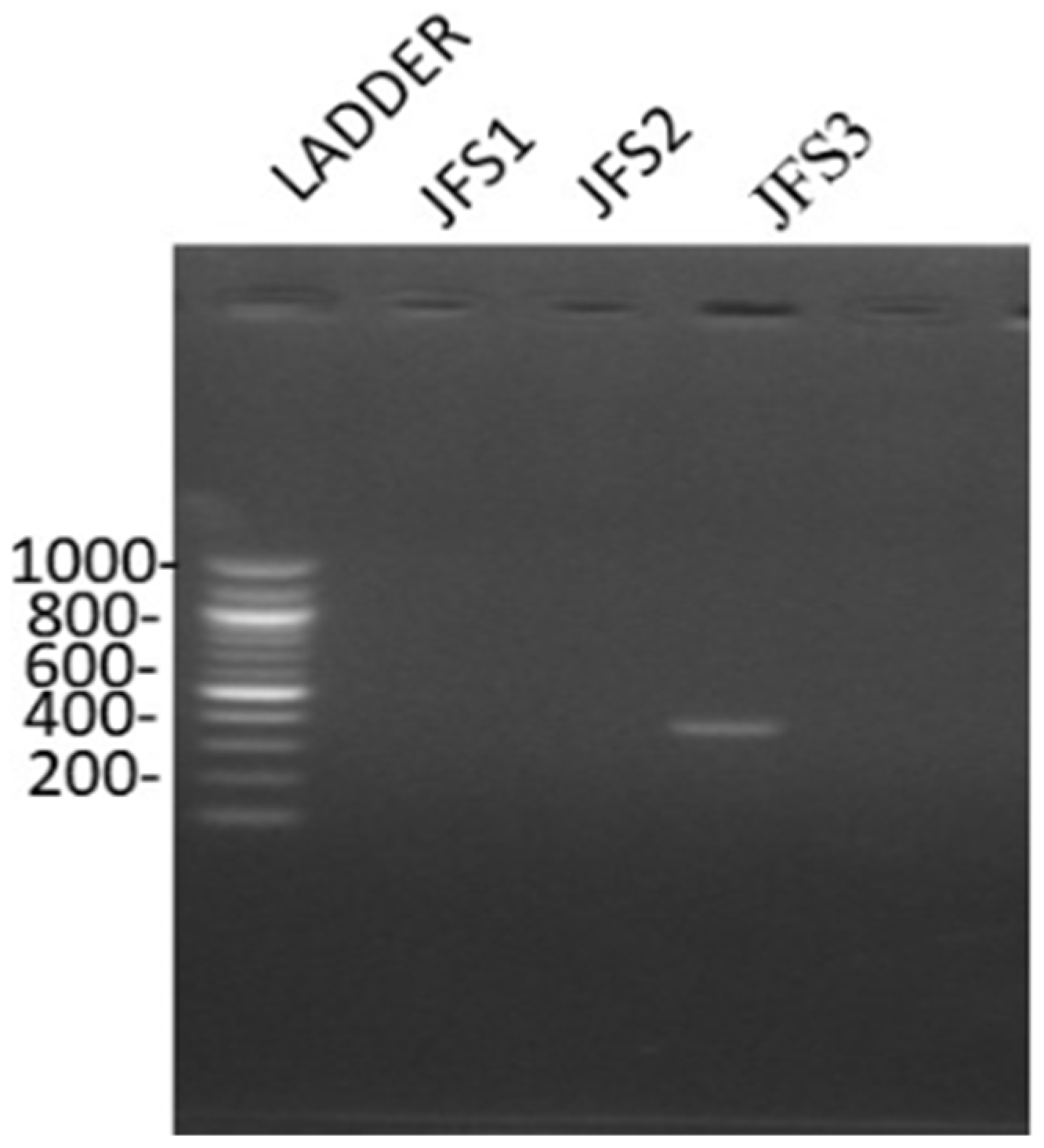
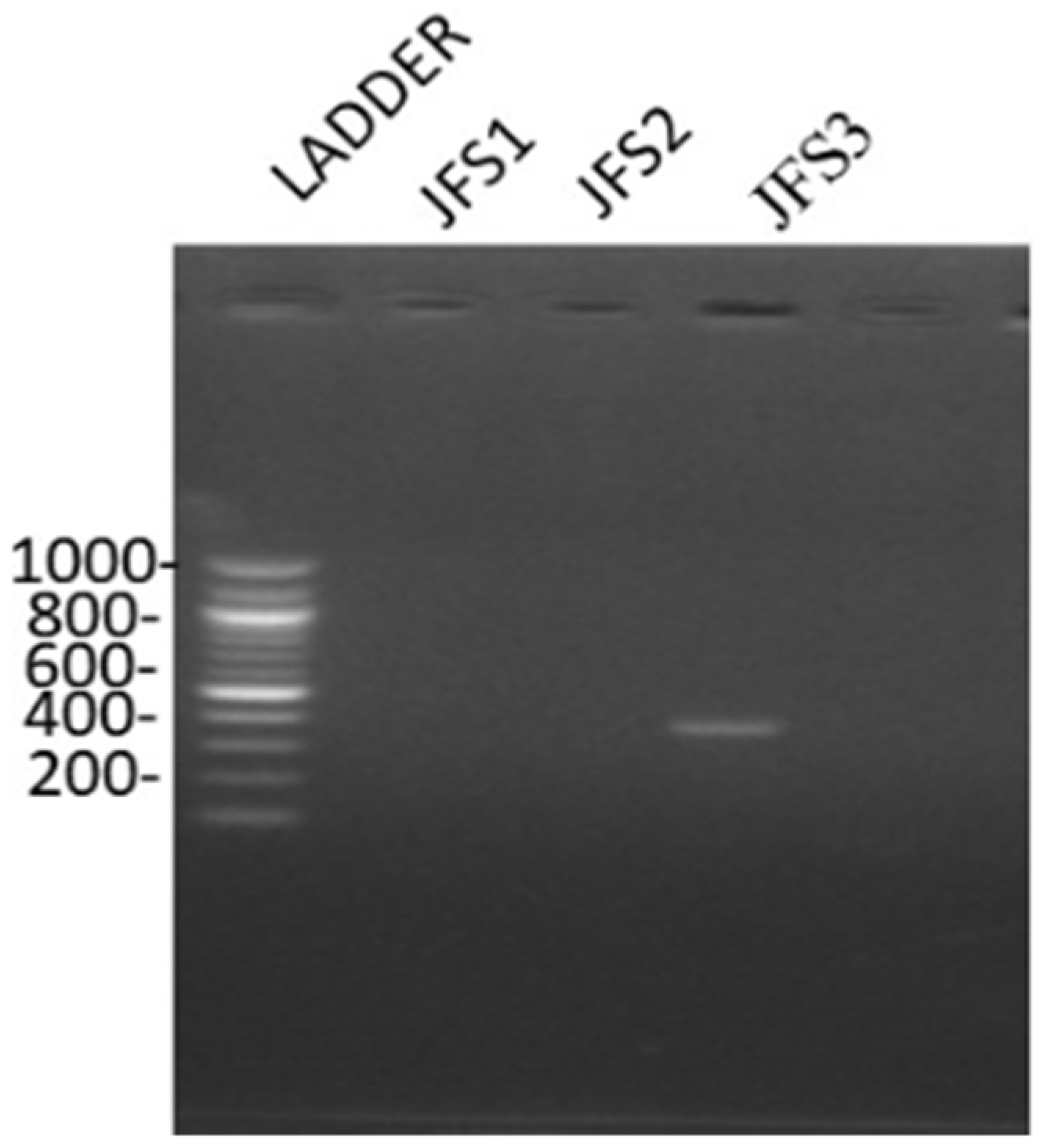
2.3.3. Molecular Characterization
2.4. Pathogenicity Test
2.5. Extraction of Chitin and Chitosan
2.6. Antimicrobial Effects of BSF Chitosan on Ralstonia solanacearum
2.7. Effects of BSF Chitosan on Ralstonia solanacearum Symptom Expression in Tomato
2.8. Data Collection and Analysis
3. Results and Discussion
3.1. Characterization of Ralstonia solanacearum
3.2. Pathogenicity Test
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chandrashekara, K.; Kumar, M.P.; Saroja, S. Aggressiveness of Ralstonia solanacearum isolates on tomato. J. Exp. Sci. 2012, 3, 5–9. [Google Scholar]
- Fegan, M.; Prior, P. How complex is the Ralstonia solanacearum species complex. Bact. Wilt Dis. Ralstonia Solanacearum Species Complex 2005, 1, 449–461. [Google Scholar]
- Tahat, M.M.; Sijam, K. Ralstoina solanacearum: The bacterial wilt causal agent. Asian J. Plant Sci. 2010, 9, 385. [Google Scholar] [CrossRef]
- Iraboneye, N.; Charimbu, M.K.; Mungai, N.W. Effect of Canola and Compound Fertilizer on Potato (Solanum Tuberosum L.) Bacterial Wilt Management. Eur. J. Agric. Food Sci. 2021, 3, 28–38. [Google Scholar] [CrossRef]
- Ahing, F.A.; Wid, N. Extraction and characterization of chitosan from shrimp shell waste in Sabah. Trans. Sci. Technol. 2016, 3, 227–237. [Google Scholar]
- Nion, Y.A.; Toyota, K. Recent trends in control methods for bacterial wilt diseases caused by Ralstonia solanacearum. Microbes Environ. 2015, 30, ME14144. [Google Scholar]
- Campos, M.D.; Felix, M.d.R.; Patanita, M.; Materatski, P.; Varanda, C. High throughput sequencing unravels tomato-pathogen interactions towards a sustainable plant breeding. Hortic. Res. 2021, 8, 171. [Google Scholar] [CrossRef] [PubMed]
- Elphinstone, J.G. The current bacterial wilt situation: A global overview. In Bacterial Wilt Disease And The Ralstonia Solanacearum Species Complex; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2005; pp. 9–28. [Google Scholar]
- Teli, K.; Shweta, H.; Kumar, M.P.; Kunduru, B.; Shekar, B.C. Isolation, identification and molecular characterization of Ralstonia solanacerum isolates collected from Southern Karnataka. J. Appl. Nat. Sci. 2018, 10, 886–893. [Google Scholar] [CrossRef]
- Hassan, O.; Chang, T. Chitosan for eco-friendly control of plant disease. Asian J. Plant Pathol. 2017, 11, 53–70. [Google Scholar] [CrossRef]
- Fan, Z.; Qin, Y.; Liu, S.; Xing, R.; Yu, H.; Chen, X.; Li, K.; Li, R.; Wang, X.; Li, P. The bioactivity of new chitin oligosaccharide dithiocarbamate derivatives evaluated against nematode disease (Meloidogyne incognita). Carbohydr. Polym. 2019, 224, 115155. [Google Scholar] [CrossRef] [PubMed]
- Malerba, M.; Cerana, R. Recent applications of chitin-and chitosan-based polymers in plants. Polymers 2019, 11, 839. [Google Scholar] [CrossRef] [Green Version]
- Chamedjeu, R.R. Prospecting for Biological Control Agents against Ralstonia Solanacearum in Potato. Ph.D. Thesis, Jkuat-Pausti, Juja, Kenya, 2018. [Google Scholar]
- Algam, S.; Xie, G.; Li, B.; Yu, S.; Su, T.; Larsen, J. Effects of Paenibacillus strains and chitosan on plant growth promotion and control of Ralstonia wilt in tomato. J. Plant Pathol. 2010, 92, 593–600. [Google Scholar]
- Buyela, D.K. Profiling and Pathogenicity of Ralstonia Solanacearum Disease of Tomato And It’s Control Using Senna Didymobotrya and Moringa Oleifera Plant Extracts in Maseno (Kenya). Ph.D. Thesis, Maseno University, Kisumu, Kenya, 2017. [Google Scholar]
- Gitahi, D.W. Evaluation of Knowledge and Practices of Managing CITRUS Pests and Diseases and the Willingness to Pay for an Integrated Pest Management Strategy in Selected Counties in Kenya. Ph.D. Thesis, Egerton University, Njoro, Kenya, 2018. [Google Scholar]
- Muzzarelli, R.A. Chitin; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Rajestary, R.; Landi, L.; Romanazzi, G. Chitosan and postharvest decay of fresh fruit: Meta-analysis of disease control and antimicrobial and eliciting activities. Compr. Rev. Food Sci. Food Saf. 2021, 20, 563–582. [Google Scholar] [CrossRef]
- Jana, S.; Jana, S. Functional Chitosan: Drug Delivery and Biomedical Applications; Springer Nature: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Aranaz, I.; Acosta, N.; Civera, C.; Elorza, B.; Mingo, J.; Castro, C.; Gandía, M.D.l.L.; Heras Caballero, A. Cosmetics and cosmeceutical applications of chitin, chitosan and their derivatives. Polymers 2018, 10, 213. [Google Scholar] [CrossRef] [Green Version]
- Kipkoech, C.; Kinyuru, J.N.; Imathiu, S.; Meyer-Rochow, V.B.; Roos, N. Invitro Study Of Cricket Chitosan’s Potential As A Prebiotic And A Promoter Of Probiotic Microorganisms To Control Pathogenic Bacteria In The Human Gut. Foods 2021, 10, 2310. [Google Scholar] [CrossRef]
- Sharma, D.; Singh, Y. Characterization of Ralstonia solanacearum isolates using biochemical, cultural, molecular methods and pathogenicity tests. J. Pharmacogn. Phytochem. 2019, 8, 2884–2889. [Google Scholar]
- Khasabulli, B.D.; Musyimi, D.M.; Miruka, D.M.; Opande, G.T.; Jeruto, P. Isolation and characterisation of Ralstonia solanacearum strains of tomato wilt disease from Maseno, Kenya. J. Asian Sci. Res. 2017, 7, 404–420. [Google Scholar] [CrossRef] [Green Version]
- Bawari, M.R.; Narendrappa, T. Cultural, biochemical and physiological studies of Ralstonia solanacearum causing wilt of brinjal. J. Pharmacogn. Phytochem. 2019, 8, 2042–2044. [Google Scholar]
- Pawaskar, J.; Joshi, M.; Navathe, S.; Agale, R. Physiological and biochemical characters of ralstonia solanacearum. Int. J. Res. Agric. Sci. 2014, 1, 2348–3997. [Google Scholar]
- Hossain, M.; Iqbal, A. Production and characterization of chitosan from shrimp waste. J. Bangladesh Agric. Univ. 2014, 12, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Kones, C.; Mwajita, M.; Kariuki, L.; Kiirika, L.; Kavoo, A. Isolation and characterization of rhizospheric microorganisms from bacterial wilt endemic areas in Kenya. Afr. J. Microbiol. Res. 2020, 14, 349–360. [Google Scholar]
- Kiirika, L.M.; Stahl, F.; Wydra, K. Phenotypic and molecular characterization of resistance induction by single and combined application of chitosan and silicon in tomato against Ralstonia solanacearum. Physiol. Mol. Plant Pathol. 2013, 81, 1–12. [Google Scholar] [CrossRef]
- Kariuki, C. Evaluation of Bacillus and Trichoderma Species for Biological Control of Bacterial Wilt Caused by Ralstonia Solanacearum in Tomato. Ph.D. Thesis, University of Nairobi, Nairobi, Kenya, 2020. [Google Scholar]
- Kaya, M.; Erdogan, S.; Mol, A.; Baran, T. Comparison of chitin structures isolated from seven Orthoptera species. Int. J. Biol. Macromol. 2015, 72, 797–805. [Google Scholar] [CrossRef]
- Kaya, M.; Baublys, V.; Šatkauskienė, I.; Akyuz, B.; Bulut, E.; Tubelytė, V. First chitin extraction from Plumatella repens (Bryozoa) with comparison to chitins of insect and fungal origin. Int. J. Biol. Macromol. 2015, 79, 126–132. [Google Scholar] [CrossRef]
- Spirescu, V.A.; Chircov, C.; Grumezescu, A.M.; Vasile, B.Ș.; Andronescu, E. Inorganic Nanoparticles and Composite Films for Antimicrobial Therapies. Int. J. Mol. Sci. 2021, 22, 4595. [Google Scholar] [CrossRef]
- Khameneh, B.; Eskin, N.; Iranshahy, M.; Fazly Bazzaz, B.S. Phytochemicals: A Promising Weapon in the Arsenal against Antibiotic-Resistant Bacteria. Antibiotics 2021, 10, 1044. [Google Scholar] [CrossRef]
- Kempe, J.; Sequeira, L. Biological control of bacterial wilt of potatoes: Attempts to induce resistance by treating tubers with bacteria. Plant Dis. 1983, 67, 499–503. [Google Scholar] [CrossRef]
- Ibrahim, Y.E.; Balabel, N.M.; Saleh, A.A.; Farag, N.S. Determination of differences in Ralstonia solanacearum phylotype II, sequevar 1 forms as related to their colony characteristics on Kelman medium and pathogenesis. J. Plant Pathol. 2020, 102, 59–66. [Google Scholar] [CrossRef]
- Mutimawurugo, M.; Wagara, I.; Muhinyuza, J.; Ogweno, J. Virulence and characterization of isolates of potato bacterial wilt caused by Ralstonia solanacearum (Smith) in Rwanda. Afr. J. Agric. Res. 2019, 14, 311–320. [Google Scholar]
- Tondo, M.L.; de Pedro-Jové, R.; Vandecaveye, A.; Piskulic, L.; Orellano, E.G.; Valls, M. KatE from the bacterial plant pathogen Ralstonia solanacearum is a monofunctional catalase controlled by HrpG that plays a major role in bacterial survival to hydrogen peroxide. Front. Plant Sci. 2020, 11, 1156. [Google Scholar] [CrossRef]
- Paudel, S.; Dobhal, S.; Alvarez, A.M.; Arif, M. Taxonomy and Phylogenetic Research on Ralstonia solanacearum Species Complex: A Complex Pathogen with Extraordinary Economic Consequences. Pathogens 2020, 9, 886. [Google Scholar] [CrossRef]
- Choi, C.H.; Lee, J.S.; Lee, Y.C.; Park, T.I.; Lee, J.C. Acinetobacter baumannii invades epithelial cells and outer membrane protein A mediates interactions with epithelial cells. BMC Microbiol. 2008, 8, 216. [Google Scholar] [CrossRef] [Green Version]
- Morin-Crini, N.; Lichtfouse, E.; Torri, G.; Crini, G. Applications of chitosan in food, pharmaceuticals, medicine, cosmetics, agriculture, textiles, pulp and paper, biotechnology, and environmental chemistry. Environ. Chem. Lett. 2019, 17, 1667–1692. [Google Scholar] [CrossRef] [Green Version]
- Kumaraswamy, R.; Kumari, S.; Choudhary, R.C.; Pal, A.; Raliya, R.; Biswas, P.; Saharan, V. Engineered chitosan based nanomaterials: Bioactivities, mechanisms and perspectives in plant protection and growth. Int. J. Biol. Macromol. 2018, 113, 494–506. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Mehwish, H.M.; Wu, Y.; Zhao, L.; Arfat, Y.; Majeed, K.; Anwaar, S. Chitin/chitosan derivatives and their interactions with microorganisms: A comprehensive review and future perspectives. Crit. Rev. Biotechnol. 2020, 40, 365–379. [Google Scholar] [CrossRef]
- Mota, L.C.; dos Santos, M.A. Chitin and chitosan on Meloidogyne javanica management and on chitinase activity in tomato plants. Trop. Plant Pathol. 2016, 41, 84–90. [Google Scholar] [CrossRef]
- Parada, R.Y.; Egusa, M.; Aklog, Y.F.; Miura, C.; Ifuku, S.; Kaminaka, H. Optimization of nanofibrillation degree of chitin for induction of plant disease resistance: Elicitor activity and systemic resistance induced by chitin nanofiber in cabbage and strawberry. Int. J. Biol. Macromol. 2018, 118, 2185–2192. [Google Scholar] [CrossRef]
- Malerba, M.; Cerana, R. Recent advances of chitosan applications in plants. Polymers 2018, 10, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Gairola, K.; Upadhyay, V.; Kumar, J. Chitosan: An elicitor and antimicrobial Bio-resource in plant protection. Agric. Rev. 2018, 39, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Shahrajabian, M.H.; Chaski, C.; Polyzos, N.; Tzortzakis, N.; Petropoulos, S.A. Sustainable Agriculture Systems in Vegetable Production Using Chitin and Chitosan as Plant Biostimulants. Biomolecules 2021, 11, 819. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | Observation |
|---|---|
| Gram stain reaction test | − |
| Potassium Hydroxide solubility test | + |
| Catalase test | + |
| Gas production test | + |
| Starch hydrolysis test | − |
| Treatment | Means of Inhibition (mm ± Standard Error of the Mean) | p-Value |
|---|---|---|
| Sterile distilled water | 0 ± 2.35 d | <0.001 |
| 1% acetic acid without chitosan | 11.67 ± 2.35 b | 0.002 |
| BSF chitosan | 19.83 ± 1.17 a | 0.004 |
| Commercial chitosan | 18.5 ± 1.17 a | <0.001 |
| Nalidixic acid (30 µg) | 26 ± 2.349153 e | 0.002 |
| Streptomycin (25 µg) | 10.67 ± 2.35 b | 0.041 |
| Sulphamethoxazole (200 µg) | 16.67 ± 2.35 c | 0.021 |
| Cotrimoxazole (25 µg) | 21 ± 2.35 c | 0.021 <0.001 |
| Gentamycin (10 µg) | 20 ± 2.35 c | <0.001 |
| Ampicillin (25 µg) | 0 ± 2.34 d | <0.001 |
| Tetracycline (100 µg) | 20 ± 2.34 c | 0.002 |
| Nitrofurantoin (200 µg) | 15.67 ± 2.34 c | 0.002 |
| Treatment | Mean % Disease Incidence ± Standard Error of the Mean | Mean % Disease Severity ± Standard Error of Mean |
|---|---|---|
| BSF-based pupal exuviae chitin | 30.31 ±1.36 a | 22.57 ± 2.01 a |
| BSF-based pupal exuviae chitosan | 34.95 ± 0.60 a | 23.66 ± 1.15 a |
| Positive control | 44.78 ± 1.56 b | 36.95 ± 1.49 b |
| p-values | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kemboi, V.J.; Kipkoech, C.; Njire, M.; Were, S.; Lagat, M.K.; Ndwiga, F.; Wesonga, J.M.; Tanga, C.M. Biocontrol Potential of Chitin and Chitosan Extracted from Black Soldier Fly Pupal Exuviae against Bacterial Wilt of Tomato. Microorganisms 2022, 10, 165. https://doi.org/10.3390/microorganisms10010165
Kemboi VJ, Kipkoech C, Njire M, Were S, Lagat MK, Ndwiga F, Wesonga JM, Tanga CM. Biocontrol Potential of Chitin and Chitosan Extracted from Black Soldier Fly Pupal Exuviae against Bacterial Wilt of Tomato. Microorganisms. 2022; 10(1):165. https://doi.org/10.3390/microorganisms10010165
Chicago/Turabian StyleKemboi, Violah Jepkogei, Carolyne Kipkoech, Moses Njire, Samuel Were, Mevin Kiprotich Lagat, Francis Ndwiga, John Mwibanda Wesonga, and Chrysantus Mbi Tanga. 2022. "Biocontrol Potential of Chitin and Chitosan Extracted from Black Soldier Fly Pupal Exuviae against Bacterial Wilt of Tomato" Microorganisms 10, no. 1: 165. https://doi.org/10.3390/microorganisms10010165
APA StyleKemboi, V. J., Kipkoech, C., Njire, M., Were, S., Lagat, M. K., Ndwiga, F., Wesonga, J. M., & Tanga, C. M. (2022). Biocontrol Potential of Chitin and Chitosan Extracted from Black Soldier Fly Pupal Exuviae against Bacterial Wilt of Tomato. Microorganisms, 10(1), 165. https://doi.org/10.3390/microorganisms10010165