

Multi-Functional Potential of Lactic Acid Bacteria Strains and Antimicrobial Effects in Minimally Processed Pomegranate (Punica granatum L. cv Jolly Red) Arils

 ,
,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Evaluation of Functional Traits
2.2.1. Tolerance to Acidic Conditions
2.2.2. Bile Salt Tolerance
2.2.3. Pepsin Tolerance
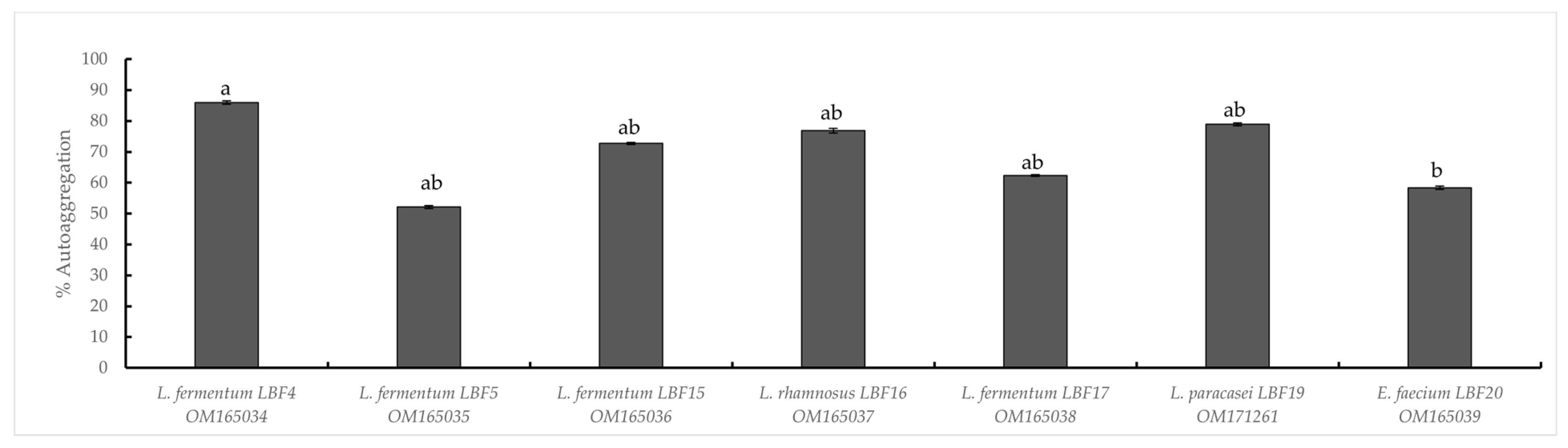
2.2.4. Auto-Aggregation Assay
2.3. Evaluation of Safety Traits
2.3.1. DNase, Gelatinase and Hemolytic Assays
2.3.2. Antibiotic Susceptibility Test
2.4. Antibacterial Activity
2.4.1. Target Strains and Growth Conditions
2.4.2. Antibacterial Activity of LAB Strains (as Cells or CFSs)
2.5. Fruit Processing and Treatments
2.5.1. Vegetal Matrix
2.5.2. Fruit Treatments
2.5.3. Physicochemical Analyses
2.6. Microbiological Analyses
2.7. Statistical Analyses
3. Results
3.1. Growth Rate of LAB Strains
3.2. Functional Traits Assessment
Acidic, Bile Salt, Pepsin Tolerance and Auto-Aggregation
3.3. Safety Evaluation
DNase, Gelatinase, Hemolytic Activity and Antibiotic Resistance
3.4. Antibacterial Activity of Lab Strains
3.5. Physicochemical Analyses of Minimally Processed Pomegranate Arils Differently Treated during Storage at Refrigerated Conditions
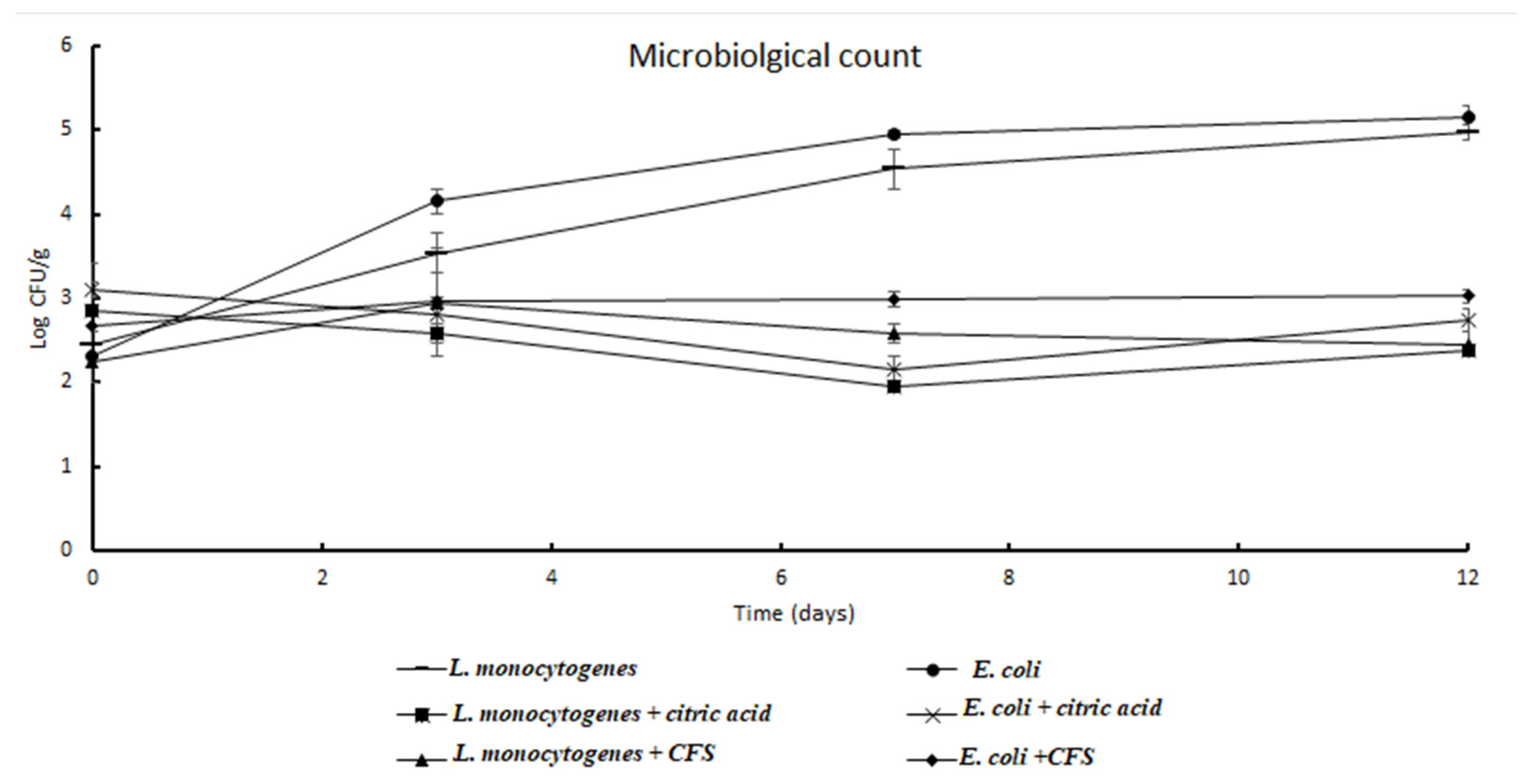
3.6. Microbiological Analyses of Minimally Processed Pomegranate Arils Differently Treated during Storage at Refrigerated Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Corato, U. Improving the shelf-life and quality of fresh and minimally-processed fruits and vegetables for a modern food industry: A comprehensive critical review from the traditional technologies into the most promising advancements. Crit. Rev. Food Sci. Nutr. 2020, 60, 940–975. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Microbiological Hazards in Fresh Leafy Vegetables and Herbs: Meeting Report; World Health Organization: Geneva, Switzerland, 2008; Volume 14. [Google Scholar]
- Caggia, C.; Scifò, G.O.; Restuccia, C.; Randazzo, C.L. Growth of acid-adapted Listeria monocytogenes in orange juice and in minimally processed orange slices. Food Control 2009, 20, 59–66. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Scifò, G.O.; Tomaselli, F.; Caggia, C. Polyphasic characterization of bacterial community in fresh cut salads. Int. J. Food Microbiol. 2009, 128, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Lokerse, R.; Maslowska-Corker, K.; Van de Wardt, L.; Wijtzes, T. Growth capacity of Listeria monocytogenes in ingredients of ready-to-eat salads. Food Control 2016, 60, 338–345. [Google Scholar] [CrossRef]
- Manzocco, L.; Da Pieve, S.; Bertolini, A.; Bartolomeoli, I.; Maifreni, M.; Vianello, A.; Nicoli, M.C. Surface decontamination of fresh-cut apple by UV-C light exposure: Effects on structure, colour and sensory properties. Postharvest Biol. Technol. 2011, 61, 165–171. [Google Scholar] [CrossRef]
- Zhang, H.; Yamamoto, E.; Murphy, J.; Locas, A. Microbiological safety of ready-to-eat fresh-cut fruits and vegetables sold on the Canadian retail market. Int. J. Food Microbiol. 2020, 335, 108855. [Google Scholar] [CrossRef]
- García-Pastor, M.E.; Zapata, P.J.; Castillo, S.; Martínez-Romero, D.; Guillén, F.; Valero, D.; Serrano, M. The effects of salicylic acid and its derivatives on increasing pomegranate fruit quality and bioactive compounds at harvest and during storage. Front. Plant Sci. 2020, 11, 668. [Google Scholar] [CrossRef]
- Oliveira, M.; Viñas, I.; Colàs, P.; Anguera, M.; Usall, J.; Abadias, M. Effectiveness of a bacteriophage in reducing Listeria monocytogenes on fresh-cut fruits and fruit juices. Food Microbiol. 2014, 38, 137–142. [Google Scholar] [CrossRef]
- Ashtari, M.; Khademi, O.; Soufbaf, M.; Afsharmanesh, H.; Sarcheshmeh, M.A.A. Effect of gamma irradiation on antioxidants, microbiological properties and shelf life of pomegranate arils cv.‘Malas Saveh’. Sci. Hortic. 2019, 244, 365–371. [Google Scholar] [CrossRef]
- Ali, A.; Yeoh, W.K.; Forney, C.; Siddiqui, M.W. Advances in postharvest technologies to extend the storage life of minimally processed fruits and vegetables. Crit. Rev. Food Sci. Nutr. 2018, 58, 2632–2649. [Google Scholar] [CrossRef]
- Martínez-Hernández, G.B.; Navarro-Rico, J.; Gómez, P.A.; Otón, M.; Artés, F.; Artés-Hernández, F. Combined sustainable sanitising treatments to reduce Escherichia coli and Salmonella Enteritidis growth on fresh-cut kailan-hybrid broccoli. Food Control 2015, 47, 312–317. [Google Scholar] [CrossRef]
- Sorrenti, V.; Randazzo, C.L.; Caggia, C.; Ballistreri, G.; Romeo, F.V.; Fabroni, S.; Timpanaro, N.; Raffaele, M.; Vanella, L. Beneficial effects of pomegranate peel extract and probiotics on pre-adipocyte differentiation. Front. Microbiol. 2019, 10, 660. [Google Scholar] [CrossRef] [PubMed]
- Laurindo, L.F.; Barbalho, S.M.; Marquess, A.R.; Grecco, A.I.d.S.; Goulart, R.d.A.; Tofano, R.J.; Bishayee, A. Pomegranate (Punica granatum L.) and metabolic syndrome risk factors and outcomes: A systematic review of clinical studies. Nutrients 2022, 14, 1665. [Google Scholar] [CrossRef] [PubMed]
- Caleb, O.J.; Opara, U.L.; Witthuhn, C.R. Modified atmosphere packaging of pomegranate fruit and arils: A review. Food Bioprocess Technol. 2012, 5, 15–30. [Google Scholar] [CrossRef]
- López-Rubira, V.; Conesa, A.; Allende, A.; Artés, F. Shelf life and overall quality of minimally processed pomegranate arils modified atmosphere packaged and treated with UV-C. Postharvest Biol. Technol. 2005, 37, 174–185. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Pitino, I.; Scifò, G.O.; Caggia, C. Biopreservation of minimally processed iceberg lettuces using a bacteriocin produced by Lactococcus lactis wild strain. Food Control 2009, 20, 756–763. [Google Scholar] [CrossRef]
- Arrioja-Bretón, D.; Mani-López, E.; Palou, E.; López-Malo, A. Antimicrobial activity and storage stability of cell-free supernatants from lactic acid bacteria and their applications with fresh beef. Food Control 2020, 115, 107286. [Google Scholar] [CrossRef]
- Bonaccorso, A.; Russo, N.; Romeo, A.; Carbone, C.; Grimaudo, M.A.; Alvarez-Lorenzo, C.; Randazzo, C.; Musumeci, T.; Caggia, C. Coating Lacticaseibacillus rhamnosus GG in Alginate Systems: An Emerging Strategy Towards Improved Viability in Orange Juice. AAPS PharmSciTech 2021, 22, 1–14. [Google Scholar]
- Moradi, M.; Kousheh, S.A.; Almasi, H.; Alizadeh, A.; Guimarães, J.T.; Yılmaz, N.; Lotfi, A. Postbiotics produced by lactic acid bacteria: The next frontier in food safety. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3390–3415. [Google Scholar] [CrossRef]
- Scarpellini, E.; Rinninella, E.; Basilico, M.; Colomier, E.; Rasetti, C.; Larussa, T.; Santori, P.; Abenavoli, L. From pre-and probiotics to post-biotics: A narrative review. Int. J. Environ. Res. Public Health 2021, 19, 37. [Google Scholar] [CrossRef]
- Vinderola, G.; Sanders, M.E.; Salminen, S. The Concept of Postbiotics. Foods 2022, 11, 1077. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.K.; Dong, N.T.N.; Nguyen, H.T.; Le, P.H. Lactic acid bacteria: Promising supplements for enhancing the biological activities of kombucha. Springerplus 2015, 4, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.C.; Gharaibeh, R.Z.; Mühlbauer, M.; Perez-Chanona, E.; Uronis, J.M.; McCafferty, J.; Fodor, A.A.; Jobin, C. Microbial genomic analysis reveals the essential role of inflammation in bacteria-induced colorectal cancer. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Meireles, A.; Giaouris, E.; Simões, M. Alternative disinfection methods to chlorine for use in the fresh-cut industry. Food Res. Int. 2016, 82, 71–85. [Google Scholar] [CrossRef]
- Siedler, S.; Balti, R.; Neves, A.R. Bioprotective mechanisms of lactic acid bacteria against fungal spoilage of food. Curr. Opin. Biotechnol. 2019, 56, 138–146. [Google Scholar] [CrossRef]
- Baccouri, O.; Ben Farhat, L.; Boukerb, A.M.; Zébré, A.; Zimmermann, K.; Domann, E.; Cambronel, M.; Barreau, M.; Maillot, O.; Rincé, I. Probiotic potential and safety evaluation of Enterococcus faecalis OB14 and OB15, isolated from traditional Tunisian Testouri cheese and Rigouta, using physiological and genomic analysis. Front. Microbiol. 2019, 10, 881. [Google Scholar] [CrossRef]
- Pino, A.; Bartolo, E.; Caggia, C.; Cianci, A.; Randazzo, C.L. Detection of vaginal lactobacilli as probiotic candidates. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Mokdad, F.H.; Benmechernene, Z.; Benyoucef, A.; Russo, N.; Randazzo, C.L.; Caggia, C.; Kihal, M. Characterization of bioactive Leuconostoc mesenteroides producing bacteriocin strains isolated from camel’s and goat’s Algerian raw milks. Int. J. Sci. Res. 2020, 76, 32–61. [Google Scholar]
- Thakur, M.; Sharma, R. Development of probiotic pomegranate beverage and its physico-chemical and microbial characterization. Int. J. Pure App. Biosci. 2017, 5, 35–41. [Google Scholar] [CrossRef]
- Russo, N.; Caggia, C.; Pino, A.; Coque, T.M.; Arioli, S.; Randazzo, C.L. Enterococcus spp. in ragusano PDO and pecorino siciliano cheese types: A snapshot of their antibiotic resistance distribution. Food Chem. Toxicol. 2018, 120, 277–286. [Google Scholar] [CrossRef]
- Rawat, S. Food Spoilage: Microorganisms and their prevention. Asian J. Plant Sci. Res. 2015, 5, 47–56. [Google Scholar]
- Committee, E.S. Guidance on selected default values to be used by the EFSA Scientific Committee, Scientific Panels and Units in the absence of actual measured data. EFSA J. 2012, 10, 2579. [Google Scholar]
- Milk, I. Milk Products: Determination of the Minimal Inhibitory Concentration (MIC) of Antibiotics Applicable to Bifidobacteria and Non-enterococcal Lactic Acid Bacteria. ISO Stand. 2010, 10932, 2010. [Google Scholar]
- Pundir, R.K.; Rana, S.; Kashyap, N.; Kaur, A. Probiotic potential of lactic acid bacteria isolated from food samples: An in vitro study. J. Appl. Pharm. Sci. 2013, 3, 85. [Google Scholar]
- Kumherová, M.; Veselá, K.; Kosová, M.; Mašata, J.; Horáčková, Š.; Šmidrkal, J. Novel potential probiotic Lactobacilli for prevention and treatment of vulvovaginal infections. Probiotics Antimicrob. Proteins 2021, 13, 163–172. [Google Scholar] [CrossRef]
- Klimko, A.I.; Cherdyntseva, T.A.; Brioukhanov, A.L.; Netrusov, A.I. In vitro evaluation of probiotic potential of selected lactic acid bacteria strains. Probiotics Antimicrob. Proteins 2020, 12, 1139–1148. [Google Scholar] [CrossRef]
- Holland, D.; Hatib, K.; Bar-Ya’akov, I. 2 Pomegranate: Botany, Horticulture, Breeding. Hortic. Rev. 2009, 35, 127–191. [Google Scholar]
- Linares-Morales, J.R.; Gutiérrez-Méndez, N.; Rivera-Chavira, B.E.; Pérez-Vega, S.B.; Nevárez-Moorillón, G.V. Biocontrol processes in fruits and fresh produce, the use of lactic acid bacteria as a sustainable option. Front. Sustain. Food Syst. 2018, 2, 50. [Google Scholar] [CrossRef]
- Bhatia, K.; Asrey, R. Minimal processing of pomegranates (Punica granatum L.)—A review on processing, quality, and shelf life. J. Food Process. Preserv. 2019, 43, e14281. [Google Scholar] [CrossRef]
- Snyder, A.B.; Worobo, R.W. The incidence and impact of microbial spoilage in the production of fruit and vegetable juices as reported by juice manufacturers. Food Control 2018, 85, 144–150. [Google Scholar] [CrossRef]
- Atta, O.M.; Manan, S.; Ul-Islam, M.; Ahmed, A.A.Q.; Ullah, M.W.; Yang, G. Development and characterization of plant oil-incorporated carboxymethyl cellulose/bacterial cellulose/glycerol-based antimicrobial edible films for food packaging applications. Adv. Compos. Hybrid Mater. 2022, 5, 973–990. [Google Scholar] [CrossRef]
- Salah, R.B.; Trabelsi, I.; Mansour, R.B.; Lassoued, S.; Chouayekh, H.; Bejar, S. A new Lactobacillus plantarum strain, TN8, from the gastro intestinal tract of poultry induces high cytokine production. Anaerobe 2012, 18, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Benmechernene, Z.; Chentouf, H.F.; Yahia, B.; Fatima, G.; Quintela-Baluja, M.; Calo-Mata, P.; Barros-Velázquez, J. Technological aptitude and applications of Leuconostoc mesenteroides bioactive strains isolated from Algerian raw camel milk. BioMed Res. Int. 2013, 2013, 418132. [Google Scholar] [CrossRef]
- Shazadi, K.; Arshad, N. Evaluation of inhibitory and probiotic properties of lactic acid bacteria isolated from vaginal microflora. Folia Microbiol. 2022, 1–19. [Google Scholar] [CrossRef]
- Ganguly, N.; Bhattacharya, S.; Sesikeran, B.; Nair, G.; Ramakrishna, B.; Sachdev, H.; Batish, V.; Kanagasabapathy, A.; Muthuswamy, V.; Kathuria, S. ICMR-DBT guidelines for evaluation of probiotics in food. Indian J. Med. Res. 2011, 134, 22. [Google Scholar]
- Sirichokchatchawan, W.; Pupa, P.; Praechansri, P.; Am-In, N.; Tanasupawat, S.; Sonthayanon, P.; Prapasarakul, N. Autochthonous lactic acid bacteria isolated from pig faeces in Thailand show probiotic properties and antibacterial activity against enteric pathogenic bacteria. Microb. Pathog. 2018, 119, 208–215. [Google Scholar] [CrossRef]
- da Costa, W.K.A.; de Souza, G.T.; Brandão, L.R.; de Lima, R.C.; Garcia, E.F.; dos Santos Lima, M.; de Souza, E.L.; Saarela, M.; Magnani, M. Exploiting antagonistic activity of fruit-derived Lactobacillus to control pathogenic bacteria in fresh cheese and chicken meat. Food Res. Int. 2018, 108, 172–182. [Google Scholar] [CrossRef]
- Giles-Gómez, M.; García, J.G.S.; Matus, V.; Quintana, I.C.; Bolívar, F.; Escalante, A. In vitro and in vivo probiotic assessment of Leuconostoc mesenteroides P45 isolated from pulque, a Mexican traditional alcoholic beverage. SpringerPlus 2016, 5, 1–10. [Google Scholar] [CrossRef]
- Shah, N.; Patel, A.; Ambalam, P.; Holst, O.; Ljungh, A.; Prajapati, J. Determination of an antimicrobial activity of Weissella confusa, Lactobacillus fermentum, and Lactobacillus plantarum against clinical pathogenic strains of Escherichia coli and Staphylococcus aureus in co-culture. Ann. Microbiol. 2016, 66, 1137–1143. [Google Scholar] [CrossRef]
- Mani-López, E.; Arrioja-Bretón, D.; López-Malo, A. The impacts of antimicrobial and antifungal activity of cell-free supernatants from lactic acid bacteria in vitro and foods. Compr. Rev. Food Sci. Food Saf. 2022, 21, 604–641. [Google Scholar] [CrossRef] [PubMed]
- Nanda, S.; Rao, D.S.; Krishnamurthy, S. Effects of shrink film wrapping and storage temperature on the shelf life and quality of pomegranate fruits cv. Ganesh. Postharvest Biol. Technol. 2001, 22, 61–69. [Google Scholar] [CrossRef]
- D’Aquino, S.; Palma, A.; Schirra, M.; Continella, A.; Tribulato, E.; La Malfa, S. Influence of film wrapping and fludioxonil application on quality of pomegranate fruit. Postharvest Biol. Technol. 2010, 55, 121–128. [Google Scholar] [CrossRef]
- Lufu, R.; Ambaw, A.; Opara, U.L. The Influence of internal packaging (liners) on moisture dynamics and physical and physiological quality of pomegranate fruit during cold storage. Foods 2021, 10, 1388. [Google Scholar] [CrossRef]
- Martí, N.; Pérez-Vicente, A.; García-Viguera, C. Influence of storage temperature and ascorbic acid addition on pomegranate juice. J. Sci. Food Agric. 2002, 82, 217–221. [Google Scholar] [CrossRef]
- Kapetanakou, A.; Stragkas, I.; Skandamis, P. Developing an antimicrobial packaging of ready-to-eat pomegranate arils based on vapors of brandy or distillery ethanol. Food Res. Int. 2015, 69, 141–150. [Google Scholar] [CrossRef]
- Zahran, A.; Hassanein, R.A.; AbdelWahab, A.T. Effect of chitosan on biochemical composition and antioxidant activity of minimally processed ‘Wonderful’pomegranate arils during cold storage. J. Appl. Bot. Food Qual. 2015, 88, 241–248. [Google Scholar]
- Cirillo, A.; Magri, A.; Scognamiglio, M.; D’Abrosca, B.; Fiorentino, A.; Petriccione, M.; Di Vaio, C. Evaluation of morphological, qualitative, and metabolomic traits during fruit ripening in pomegranate (Punica granatum L.). Horticulturae 2022, 8, 384. [Google Scholar] [CrossRef]
- Feng, T.; Wang, J.; Yang, G.; Yang, J.; Wu, M. Pomegranate-structured ZnMn2O4 microspheres for long cycle life lithium ion anode and elucidation of its conversion mechanism. J. Electrochem. Soc. 2020, 167, 060507. [Google Scholar] [CrossRef]
- Ranjbar, A.; Ramezanian, A.; Shekarforoush, S.; Niakousari, M.; Eshghi, S. Antifungal activity of thymol against the main fungi causing pomegranate fruit rot by suppressing the activity of cell wall degrading enzymes. LWT 2022, 161, 113303. [Google Scholar] [CrossRef]
- Dong, F.; Wang, X. Effects of carboxymethyl cellulose incorporated with garlic essential oil composite coatings for improving quality of strawberries. Int. J. Biol. Macromol. 2017, 104, 821–826. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Wu, R.; Strik, B.C.; Zhao, Y. Effect of edible coatings on the quality of fresh blueberries (Duke and Elliott) under commercial storage conditions. Postharvest Biol. Technol. 2011, 59, 71–79. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LAB Strains | Viable Count (Log CFU/mL) | Survival Rate (%) | ||||
|---|---|---|---|---|---|---|
| T0 | T3 | SR pH 2 | SR pH 3 | |||
| pH 3 | pH 2 | pH 3 | pH 2 | |||
| L. fermentum LBF4 | 6.80 ± 0.01 a | 4.60 ± 0.22 c | 6.48 ± 0.08 | 4.17 ± 0.28 d | 90.65 | 95.29 |
| L. fermentum LBF5 | 6.47 ± 0.03 c | 6.60 ± 0.11 a | 6.34 ± 0.16 | 6.38 ± 0.05 a | 96.67 | 97.99 |
| L. fermentum LBF15 | 6.80 ± 0.02 a | 5.75 ± 0.05 b | 6.42 ± 0.05 | 5.33 ± 0.03 b | 92.69 | 94.41 |
| L. rhamnosus LBF16 | 6.47 ± 0.03 c | 5.66 ± 0.03 b | 6.15 ± 0.27 | 5.08 ± 0.11 ab | 89.75 | 95.05 |
| L. fermentum LBF17 | 6.60 ± 0.07 b | 5.51 ± 0.19 b | 6.37 ± 0.09 | 5.07 ± 0.07 ab | 91.01 | 96.51 |
| L. paracasei LBF19 | 6.57 ± 0.01 bc | 5.79 ± 0.10 b | 6.25 ± 0.02 | 5.38 ± 0.16 b | 92.91 | 95.13 |
| E. faecium LBF20 | 6.52 ± 0.02 bc | 5.54 ± 0.05 b | 6.17 ± 0.03 | 4.75 ± 0.05 c | 85.74 | 94.63 |
| Sig. | ** | ** | n.s. | ** | ||
| LAB Strains | Viable Count (Log CFU/mL) | Survival Rate (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| T0 | T3 | ||||||||
| 0.5% | 1.0% | 2.0% | 0.5% | 1.0% | 2.0% | SR 0.5% | SR 1.0% | SR 2.0% | |
| L. fermentum LBF4 | 8.72 ± 0.03 | 7.43 ± 0.07 a | 6.27 ± 0.11 b | 8.56 ± 0.05 a | 7.17 ± 0.07 a | 5.52 ± 0.31 ab | 98.16 | 96.50 | 88.04 |
| L. fermentum LBF5 | 8.81 ± 0.01 | 6.33 ± 0.31 c | 5.51 ± 0.04 c | 8.61 ± 0.18 a | 6.02 ± 0.10 b | 4.93 ± 0.00 b | 97.73 | 95.10 | 89.47 |
| L. fermentum LBF15 | 8.93 ± 0.08 | 7.15 ± 0.21 ab | 6.75 ± 0.05 a | 8.68 ± 0.03 a | 6.79 ± 0.11 a | 6.27 ± 0.02 a | 97.20 | 94.96 | 92.89 |
| L. rhamnosus LBF16 | 8.52 ± 0.14 | 5.02 ± 0.34 d | 2.79 ± 0.04 d | 8.29 ± 0.28 ab | 4.69 ± 0.33 c | 2.38 ± 0.31 c | 97.30 | 93.43 | 85.30 |
| L. fermentum LBF17 | 8.14 ± 0.18 | 7.54 ± 0.00 a | 6.66 ± 0.01 a | 7.91 ± 0.01 b | 7.30 ± 0.01 a | 5.51 ± 0.04 ab | 97.17 | 96.82 | 82.73 |
| L. paracasei LBF19 | 8.61 ± 0.07 | 6.53 ± 0.07 bc | 5.74 ± 0.16 c | 8.33 ± 0.16 ab | 6.21 ± 0.03 b | 5.06 ± 0.64 b | 96.75 | 95.10 | 88.15 |
| E. faecium LBF20 | 8.49 ± 0.52 | 6.47 ± 0.12 bc | 6.41 ± 0.10 ab | 8.10 ± 0.11 ab | 5.68 ± 0.02 b | 5.25 ± 0.02 ab | 95.41 | 87.79 | 81.90 |
| Sig. | n.s. | ** | ** | ** | ** | ** | |||
| Strains | Viable Count (Log CFU/mL) | Survival Rate (%) | ||||
|---|---|---|---|---|---|---|
| T0 | T3 | SR pH3 | SR pH2 | |||
| pH 3 | pH 2 | pH 3 | pH 2 | |||
| L. fermentum LBF4 | 7.22 ± 0.25 a | 6.83 ± 0.05 a | 6.97 ± 0.26 | 6.38 ± 0.22 | 96.54 | 93.41 |
| L. fermentum LBF5 | 6.55 ± 0.01 b | 6.54 ± 0.05 abc | 6.39 ± 0.04 | 6.23 ± 0.03 | 97.56 | 95.26 |
| L. fermentum LBF15 | 6.72 ± 0.03 b | 6.79 ± 0.11 ab | 6.42 ± 0.02 | 6.29 ± 0.00 | 95.54 | 92.64 |
| L. rhamnosus LBF16 | 6.59 ± 0.02 b | 6.43 ± 0.03 c | 6.38 ± 0.16 | 6.14 ± 0.33 | 96.81 | 95.49 |
| L. fermentum LBF17 | 6.56 ± 0.04 b | 6.60 ± 0.08 abc | 6.37 ± 0.54 | 6.31 ± 0.04 | 97.10 | 95.61 |
| L. paracasei LBF19 | 6.56 ± 0.07 b | 6.60 ± 0.08 abc | 6.35 ± 0.53 | 6.32 ± 0.02 | 96.80 | 95.76 |
| E. faecium LBF20 | 6.60 ± 0.11 b | 6.47 ± 0.12 bc | 6.38 ± 0.06 | 6.20 ± 0.09 | 96.67 | 95.83 |
| Sig. | ** | * | n.s. | n.s. | ||
| Strains | Antibiotic (µg/mL) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| AMP (≥2 a) | VAN (nr) | GEN (≥16 a) | KAN (≥32 a) | STRE (≥64 a) | ERY (≥1 a) | CLI (≥1 a) | TET (≥8 a) | CHL (≥4 a) | |
| L. fermentum LBF4 | 2 a | 0.5 nr | 16 a | 16 a | 32 a | 1 a | 0.5 a | 4 a | 4 a |
| L. fermentum LBF5 | 2 a | 16 nr | 16 a | 32 a | 64 a | 1 a | 1 a | 8 a | 4 a |
| L. fermentum LBF15 | 4 R | 4 nr | 8 a | 128 R | 8 a | 2 R | 1 a | 4 a | 1 a |
| L. rhamnosus LBF16 | 4 a | 32 nr | 8 a | 64 a | 8 a | 1 a | 0.5 a | 1 a | 4 a |
| L. fermentum LBF17 | 32 R | 16 nr | 4 a | 256 R | 4 a | 4 R | 0.5 a | 4 a | 4 a |
| L. paracasei LBF19 | 4 | 16 nr | 16 a | 64 a | 256 R | 0.5 a | 0.5 a | 4 | 16 R |
| E. faecium LBF20 | 32 R | 16 R | 8 a | 256 a | 256 R | 1 a | 4 a | 16 R | 32 R |
| Samples | Time (Days) | Firmness (N) | L* (D65) | a* (D65) | b* (D65) |
|---|---|---|---|---|---|
| Fresh pomegranate arils | T0 | 10.18 ± 1.39 | 34.93 ± 1.17 | 5.67 ± 1.39 | 1.35 ± 0.57 |
| Control | T3 | 7.31 ± 1.97 n.s | 33.09 ± 2.19 n.s. | 9.91 ± 2.44 a | 3.86 ± 1.77 a |
| Pathogens + citric acid | T3 | 6.75 ± 1.94 n.s. | 32.59 ± 1.09 n.s. | 5.97 ± 3.58 ab | 1.48 ± 1.37 ab |
| Pathogens | T3 | 7.48 ± 0.56 n.s. | 32.06 ± 0.96 n.s. | 6.28 ± 2.55 a | 1.46 ± 0.93 ab |
| Pathogens + CFS | T3 | 6.39 ± 0.77 n.s | 32.68 ± 0.94 n.s. | 5.17 ± 2.31 b | 1.13 ± 0.19 b |
| Control | T7 | 9.15 ± 1.56 a | 34.00 ± 1.19 n.s | 6.98 ± 2.24 n.s | 1.54 ± 1.20 n.s. |
| Pathogens + citric acid | T7 | 7.72 ± 1.67 ab | 34.22 ± 1.54 n.s. | 8.91 ± 3.95 n.s | 2.63 ± 1.60 n.s. |
| Pathogens | T7 | 9.05 ± 0.58 a | 33.62 ± 1.60 n.s. | 8.56 ± 3.95 n.s. | 2.22 ± 1.45 n.s. |
| Pathogens + CFS | T7 | 7,08 ± 0.93 b | 33.50 ± 1.20 n.s. | 7.49 ± 1.57 n.s. | 2.76 ± 1.35 n.s. |
| Control | T12 | 6.94 ± 1.99 n.s. | 34.97 ± 1.73 a | 6.38 ± 1.98 ab | 1.62 ± 1.07 ab |
| Pathogens + citric acid | T12 | 6.75 ± 0.84 n.s. | 31.73 ± 1.09 b | 5.01 ± 2.03 b | 0.84 ± 1.07 b |
| Pathogens | T12 | 7.48 ± 0.56 n.s. | 31.64 ± 1.20 b | 6.79 ± 1.37 ab | 1.76 ± 0.63 ab |
| Pathogens + CFS | T12 | 6.38 ± 0.77 n.s. | 32.47 ± 0.69 b | 8.76 ± 2.65 a | 2.00 ± 0.53 a |
| Sample | Time (Days) | pH | TSS (mg/L) | Citric Acid (mg/L) | Lactic Acid (mg/L) | Acetic Acid (mg/L) | Propionic Acid (mg/L) |
|---|---|---|---|---|---|---|---|
| Fresh pomegranate arils | T0 | 4.42 ± 0.03 | 19.97 ± 0.06 | 2443.47 ± 0.22 | 72.75 ± 3.45 | 0.00 ± 0.00 | 72.77 ± 0.03 |
| CFS | T0 | 4.16 ± 0.05 | ---- | 3023.48 ± 30.43 | 6785.38 ± 50.02 | 2878.54 ± 67.08 | 31.63 ± 1.3 |
| Control | T3 | 4.13 ± 0.01 a | 16.94 ± 0.02 n.s | 2458.12 ± 25.32 a | 2155.86 ± 169.16 a | 0.00 ± 0.00 b | 453.05 ± 7.77 n.s. |
| Pathogens + citric acid | T3 | 3.95 ± 0.02 ab | 16.31 ± 0.01 n.s | 2374.97 ± 0.01 ab | 1405.00 ± 33.05 b | 0.00 ± 0.00 b | 452.76 ± 0.89 n.s. |
| Pathogens | T3 | 3.90 ± 0.01 b | 16.41 ± 0.03 n.s | 2311.49 ± 41.66 b | 2305.90 ± 33.12 a | 228.63 ± 20.64 a | 420.83 ± 23.67 n.s. |
| Pathogens + CFS | T3 | 4.12 ± 0.04 a | 16.32 ± 0.02 n.s | 2425.13 ± 24.64 a | 1625.35 ± 92.74 ab | 0.00 ± 0.00 b | 372.00 ± 3.00 n.s |
| Control | T7 | 3.90 ± 0.03 a | 16.31 ± 0.01 n.s | 2317.06 ± 6.46 n.s. | 1575.06 ± 75.89 n.s | 175.50 ± 32.10 n.s. | 434.90 ± 8.29 a |
| Pathogens + citric acid | T7 | 3.83 ± 0.01 b | 16.06 ± 0.07 n.s | 2307.32 ± 4.66 n.s | 1541.71 ± 176.52 n.s | 183.24 ± 8.06 n.s. | 341.11 ± 9.00 c |
| Pathogens | T7 | 3.85 ± 0.01 b | 16.53 ± 0.02 n.s | 2314.23 ± 18.37n.s. | 1767.40 ± 105.99 n.s | 204.33 ± 20.23 n.s. | 413.45 ± 30.75 ab |
| Pathogens + CFS | T7 | 3.86 ± 0.02 ab | 16.26 ± 0.08 n.s | 2386.38 ± 14.7 n.s. | 1430.47 ± 203.20 n.s | 159.13 ± 5.02 n.s. | 380.83 ± 8.52 bc |
| Control | T12 | 3.77 ± 0.06 a | 15.09 ± 0.01 b | 2317.39 ± 13.45 a | 1327.94 ± 201.57 a | 568.72 ± 59.13 b | 434.33 ± 5.51 n.s. |
| Pathogens + citric acid | T12 | 3.51 ± 0.02 c | 14.94 ± 0.12 b | 2288.66 ± 15.54 ab | 895.26 ± 43.98 b | 1072.07 ± 29.41 a | 407.34 ± 16.70 n.s. |
| Pathogens | T12 | 3.66 ± 0.01 b | 14.75 ± 0.08 c | 2248.04 ± 12.16 b | 916.13 ± 23.14 b | 601.48 ± 72.15 b | 368.14 ± 26.11 n.s. |
| Pathogens + CFS | T12 | 3.49 ± 0.02 c | 15.33 ± 0.01 a | 2287.71 ± 6.57 ab | 1165.06 ± 41.77 a | 1179.62 ± 8.21 a | 425.32 ± 20.91 n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben Farhat, L.; Romeo, F.V.; Foti, P.; Russo, N.; Randazzo, C.L.; Caggia, C.; Abidi, F. Multi-Functional Potential of Lactic Acid Bacteria Strains and Antimicrobial Effects in Minimally Processed Pomegranate (Punica granatum L. cv Jolly Red) Arils. Microorganisms 2022, 10, 1876. https://doi.org/10.3390/microorganisms10101876
Ben Farhat L, Romeo FV, Foti P, Russo N, Randazzo CL, Caggia C, Abidi F. Multi-Functional Potential of Lactic Acid Bacteria Strains and Antimicrobial Effects in Minimally Processed Pomegranate (Punica granatum L. cv Jolly Red) Arils. Microorganisms. 2022; 10(10):1876. https://doi.org/10.3390/microorganisms10101876
Chicago/Turabian StyleBen Farhat, Leila, Flora Valeria Romeo, Paola Foti, Nunziatina Russo, Cinzia Lucia Randazzo, Cinzia Caggia, and Ferid Abidi. 2022. "Multi-Functional Potential of Lactic Acid Bacteria Strains and Antimicrobial Effects in Minimally Processed Pomegranate (Punica granatum L. cv Jolly Red) Arils" Microorganisms 10, no. 10: 1876. https://doi.org/10.3390/microorganisms10101876
APA StyleBen Farhat, L., Romeo, F. V., Foti, P., Russo, N., Randazzo, C. L., Caggia, C., & Abidi, F. (2022). Multi-Functional Potential of Lactic Acid Bacteria Strains and Antimicrobial Effects in Minimally Processed Pomegranate (Punica granatum L. cv Jolly Red) Arils. Microorganisms, 10(10), 1876. https://doi.org/10.3390/microorganisms10101876