On the History and Applications of Congenic Strains in Cryptococcus Research


Abstract
1. Introduction
2. Diversity of Cryptococcus Pathogenic Species
3. The First Cryptococcus Congenic Pair
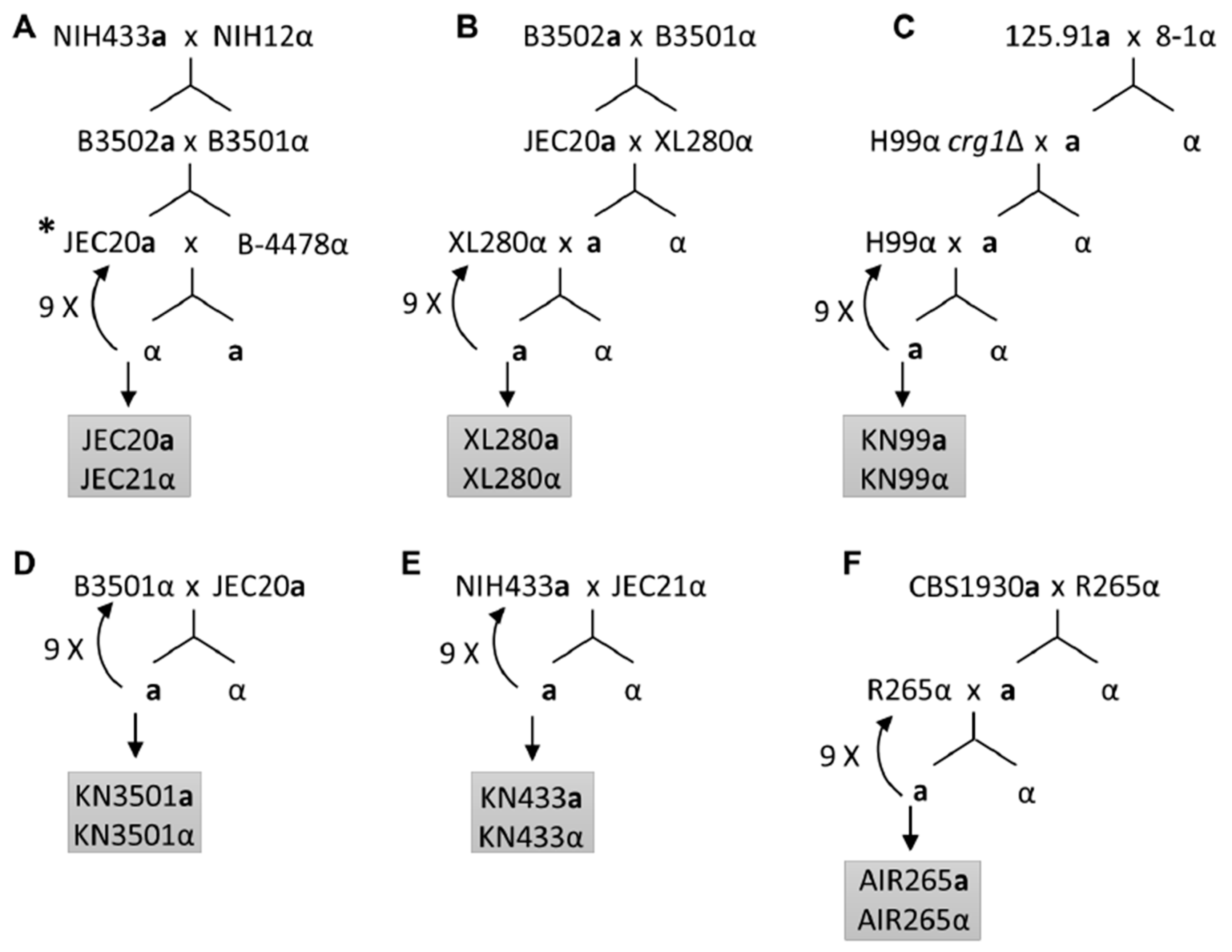
4. Summary of the Current Congenic Pairs in Cryptococcus
5. Examine the Impact of Mating Type on Virulence and Tissue Tropism
6. Examine the Impact of Mating Type on Morphogenesis
7. Determine the Genetic Factors Contributing to Uniparental Mitochondrial Inheritance
8. Limitations and Improvement of Congenic Pair Construction
9. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Charlesworth, B.; Charlesworth, D. Population genetics from 1966 to 2016. Heredity 2017, 118, 2–9. [Google Scholar] [CrossRef]
- Schacherer, J. Beyond the simplicity of Mendelian inheritance. Comptes Rendus Biol. 2016, 339, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Sahu, M.; Prasuna, J.G. Twin studies: A unique epidemiological tool. Indian J. Community Med. 2016, 41, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Snell, G.D. Methods for the study of histocompatibility genes. J. Genet. 1948, 49, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.R. A genetic analysis of species differences in Columbidae. Genetics 1939, 24, 709–721. [Google Scholar] [PubMed]
- Carlson, M.; Osmond, B.C.; Botstein, D. Genetic evidence for a silent SUC gene in yeast. Genetics 1981, 98, 41–54. [Google Scholar] [PubMed]
- Allen, S.L.; Lee, P.H.T. The preparation of congenic strains of Tetrahymena. J. Protozool. 1971, 18, 214–218. [Google Scholar] [CrossRef]
- Clark, A.G.; Bundgaard, J. Selection components in background replacement lines of Drosophila. Genetics 1984, 108, 181–200. [Google Scholar]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef]
- Bennett, J.E.; Kwon-Chung, K.J.; Theodore, T.S. Biochemical differences between serotypes of Cryptococcus neoformans. Sabouraudia 1978, 16, 167–174. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Polacheck, I.; Bennett, J.E. Improved diagnostic medium for separation of Cryptococcus neoformans var. neoformans (serotypes A and D) and Cryptococcus neoformans var. gattii (serotypes B and C). J. Clin. Microbiol. 1982, 15, 535–537. [Google Scholar] [CrossRef] [PubMed]
- Cogliati, M. Global molecular epidemiology of Cryptococcus neoformans and Cryptococcus gattii: An atlas of the molecular types. Scientifica 2013, 2013, 675213. [Google Scholar] [CrossRef]
- Hagen, F.; Khayhan, K.; Theelen, B.; Kolecka, A.; Polacheck, I.; Sionov, E.; Falk, R.; Parnmen, S.; Lumbsch, H.T.; Boekhout, T. Recognition of seven species in the Cryptococcus gattii/Cryptococcus neoformans species complex. Fungal Genet. Biol. 2015, 78, 16–48. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Vilgalys, R.; Mitchell, T.G. Multiple gene genealogies reveal recent dispersion and hybridization in the human pathogenic fungus Cryptococcus neoformans. Mol. Ecol. 2000, 9, 1471–1481. [Google Scholar] [CrossRef] [PubMed]
- Moorjani, P.; Amorim, C.E.G.; Arndt, P.F.; Przeworski, M. Variation in the molecular clock of primates. Proc. Natl. Acad. Sci. USA 2016, 113, 10607–10612. [Google Scholar] [CrossRef] [PubMed]
- Hicks, J.K.; Heitman, J. Divergence of protein kinase A catalytic subunits in Cryptococcus neoformans and Cryptococcus gattii illustrates evolutionary reconfiguration of a signaling cascade. Eukaryot. Cell 2007, 6, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Bahn, Y.-S.; Kojima, K.; Cox, G.M.; Heitman, J. Specialization of the HOG pathway and its impact on differentiation and virulence of Cryptococcus neoformans. Mol. Biol. Cell 2005, 16, 2285–2300. [Google Scholar] [CrossRef]
- Ngamskulrungroj, P.; Gilgado, F.; Faganello, J.; Litvintseva, A.P.; Leal, A.L.; Tsui, K.M.; Mitchell, T.G.; Vainstein, M.H.; Meyer, W. Genetic diversity of the Cryptococcus species complex suggests that Cryptococcus gattii deserves to have varieties. PLoS ONE 2009, 4, e5862. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J. Morphogenesis of Filobasidiella neoformans, the sexual state of Cryptococcus neoformans. Mycologia 1976, 68, 821–833. [Google Scholar] [CrossRef]
- Herskowitz, I. Life cycle of the budding yeast Saccharomyces cerevisiae. Microbiol. Rev. 1988, 52, 536–553. [Google Scholar] [CrossRef]
- Levitz, S.M. Cryptococcus neoformans by Casadevall, Arturo & Perfect, John R. (1998) ASM Press, Washington, DC. Hardcover. 542 pp. $89.95. (ASM Member price: $79.95). Med. Mycol. 1999, 37, 371. [Google Scholar] [CrossRef]
- Litvintseva, A.P.; Kestenbaum, L.; Vilgalys, R.; Mitchell, T.G. Comparative analysis of environmental and clinical populations of Cryptococcus neoformans. J. Clin. Microbiol. 2005, 43, 556–564. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lin, X.; Patel, S.; Litvintseva, A.P.; Floyd, A.; Mitchell, T.G.; Heitman, J. Diploids in the Cryptococcus neoformans serotype A population homozygous for the alpha mating type originate via unisexual mating. PLoS Pathog. 2009, 5, e1000283. [Google Scholar] [CrossRef] [PubMed]
- Cogliati, M.; D’Amicis, R.; Zani, A.; Montagna, M.T.; Caggiano, G.; De Giglio, O.; Balbino, S.; De Donno, A.; Serio, F.; Susever, S.; et al. Environmental distribution of Cryptococcus neoformans and C. gattii around the Mediterranean basin. FEMS Yeast Res. 2016, 16, fow045. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Edman, J.C.; Wickes, B.L. Genetic association of mating types and virulence in Cryptococcus neoformans. Infect. Immun. 1992, 60, 602–605. [Google Scholar] [CrossRef] [PubMed]
- Moore, T.D.; Edman, J.C. The alpha-mating type locus of Cryptococcus neoformans contains a peptide pheromone gene. Mol. Cell Biol. 1993, 13, 1962–1970. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Loftus, B.J.; Fung, E.; Roncaglia, P.; Rowley, D.; Amedeo, P.; Bruno, D.; Vamathevan, J.; Miranda, M.; Anderson, I.J.; Fraser, J.A.; et al. The genome of the basidiomycetous yeast and human pathogen Cryptococcus neoformans. Science 2005, 307, 1321–1324. [Google Scholar] [CrossRef] [PubMed]
- Hoang, L.M.N.; Maguire, J.A.; Doyle, P.; Fyfe, M.; Roscoe, D.L. Cryptococcus neoformans infections at Vancouver Hospital and Health Sciences Centre (1997–2002): Epidemiology, microbiology and histopathology. J. Med. Microbiol. 2004, 53, 935–940. [Google Scholar] [CrossRef]
- Zhu, P.; Zhai, B.; Lin, X.; Idnurm, A. Congenic strains for genetic analysis of virulence traits in Cryptococcus gattii. Infect. Immun. 2013, 81, 2616–2625. [Google Scholar] [CrossRef]
- Barchiesi, F.; Cogliati, M.; Esposto, M.C.; Spreghini, E.; Schimizzi, A.M.; Wickes, B.L.; Scalise, G.; Viviani, M.A. Comparative analysis of pathogenicity of Cryptococcus neoformans serotypes A, D and AD in murine cryptococcosis. J. Infect. 2005, 51, 10–16. [Google Scholar] [CrossRef]
- Lin, X.; Litvintseva, A.P.; Nielsen, K.; Patel, S.; Floyd, A.; Mitchell, T.G.; Heitman, J. alpha AD alpha hybrids of Cryptococcus neoformans: Evidence of same-sex mating in nature and hybrid fitness. PLoS Genet. 2007, 3, 1975–1990. [Google Scholar] [CrossRef] [PubMed]
- Feretzaki, M.; Hardison, S.E.; Wormley, F.L., Jr.; Heitman, J. Cryptococcus neoformans hyperfilamentous strain is hypervirulent in a murine model of cryptococcal meningoencephalitis. PLoS ONE 2014, 9, e104432. [Google Scholar] [CrossRef]
- Zhai, B.; Zhu, P.; Foyle, D.; Upadhyay, S.; Idnurm, A.; Lin, X. Congenic strains of the filamentous form of Cryptococcus neoformans for studies of fungal morphogenesis and virulence. Infect. Immun. 2013, 81, 2626–2637. [Google Scholar] [CrossRef]
- Nielsen, K.; Marra, R.E.; Hagen, F.; Boekhout, T.; Mitchell, T.G.; Cox, G.M.; Heitman, J. Interaction between genetic background and the mating-type locus in Cryptococcus neoformans virulence potential. Genetics 2005, 171, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Marra, R.E.; Huang, J.C.; Fung, E.; Nielsen, K.; Heitman, J.; Vilgalys, R.; Mitchell, T.G. A genetic linkage map of Cryptococcus neoformans variety neoformans serotype D (Filobasidiella neoformans). Genetics 2004, 167, 619–631. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lengeler, K.B.; Wang, P.; Cox, G.M.; Perfect, J.R.; Heitman, J. Identification of the MATa mating-type locus of Cryptococcus neoformans reveals a serotype A MATa strain thought to have been extinct. Proc. Natl. Acad. Sci. USA 2000, 97, 14455–14460. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.; Cox, G.M.; Wang, P.; Toffaletti, D.L.; Perfect, J.R.; Heitman, J. Sexual cycle of Cryptococcus neoformans var. grubii and virulence of congenic a and α Isolates. Infect. Immun. 2003, 71, 4831–4841. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.; Cox, G.M.; Litvintseva, A.P.; Mylonakis, E.; Malliaris, S.D.; Benjamin, D.K., Jr.; Giles, S.S.; Mitchell, T.G.; Casadevall, A.; Perfect, J.R.; et al. Cryptococcus neoformans alpha strains preferentially disseminate to the central nervous system during coinfection. Infect. Immun. 2005, 73, 4922–4933. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Hull, C.M.; Heitman, J. Sexual reproduction between partners of the same mating type in Cryptococcus neoformans. Nature 2005, 434, 1017. [Google Scholar] [CrossRef]
- Fu, J.; Morris, I.R.; Wickes, B.L. The production of monokaryotic hyphae by Cryptococcus neoformans can be induced by high temperature arrest of the cell cycle and is independent of same-sex mating. PLoS Pathog. 2013, 9, e1003335. [Google Scholar] [CrossRef]
- Ren, P.; Chaturvedi, V.; Chaturvedi, S. Carbon dioxide is a powerful inducer of monokaryotic hyphae and spore development in Cryptococcus gattii and carbonic anhydrase activity is dispensable in this dimorphic transition. PLoS ONE 2014, 9, e113147. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lin, J.; Zhao, Y.; Kirkman, E.; So, Y.-S.; Bahn, Y.-S.; Lin, X. Glucosamine stimulates pheromone-independent dimorphic transition in Cryptococcus neoformans by promoting Crz1 nuclear translocation. PLoS Genet. 2017, 13, e1006982. [Google Scholar] [CrossRef]
- Tscharke, R.L.; Lazera, M.; Chang, Y.C.; Wickes, B.L.; Kwon-Chung, K.J. Haploid fruiting in Cryptococcus neoformans is not mating type alpha-specific. Fungal Genet. Biol. 2003, 39, 230–237. [Google Scholar] [CrossRef]
- Wickes, B.L.; Mayorga, M.E.; Edman, U.; Edman, J.C. Dimorphism and haploid fruiting in Cryptococcus neoformans: Association with the alpha-mating type. Proc. Natl. Acad. Sci. USA 1996, 93, 7327–7331. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Huang, J.C.; Mitchell, T.G.; Heitman, J. Virulence attributes and hyphal growth of C. neoformans are quantitative traits and the MATα allele enhances filamentation. PLoS Genet. 2006, 2, e187. [Google Scholar] [CrossRef]
- Chang, Y.C.; Penoyer, L.A.; Kwon-Chung, K.J. The second STE12 homologue of Cryptococcus neoformans is MATa-specific and plays an important role in virulence. Proc. Natl. Acad. Sci. USA 2001, 98, 3258–3263. [Google Scholar] [CrossRef]
- Wickes, B.L.; Edman, U.; Edman, J.C. The Cryptococcus neoformans STE12α gene: A putative Saccharomyces cerevisiae STE12 homologue that is mating type specific. Mol. Microbiol. 1997, 26, 951–960. [Google Scholar] [CrossRef]
- Lin, X.; Jackson, J.C.; Feretzaki, M.; Xue, C.; Heitman, J. Transcription factors Mat2 and Znf2 operate cellular circuits orchestrating opposite- and same-sex mating in Cryptococcus neoformans. PLoS Genet. 2010, 6, e1000953. [Google Scholar] [CrossRef]
- Xu, J.; Ali, R.Y.; Gregory, D.A.; Amick, D.; Lambert, S.E.; Yoell, H.J.; Vilgalys, R.J.; Mitchell, T.G. Uniparental mitochondrial transmission in sexual crosses in Cryptococcus neoformans. Curr. Microbiol. 2000, 40, 269–273. [Google Scholar] [CrossRef]
- Yan, Z.; Xu, J. Mitochondria are inherited from the MATa parent in crosses of the basidiomycete fungus Cryptococcus neoformans. Genetics 2003, 163, 1315–1325. [Google Scholar]
- Yan, Z.; Hull, C.M.; Sun, S.; Heitman, J.; Xu, J. The mating type-specific homeodomain genes SXI1alpha and SXI2a coordinately control uniparental mitochondrial inheritance in Cryptococcus neoformans. Curr. Genet. 2007, 51, 187–195. [Google Scholar] [CrossRef]
- Gyawali, R.; Lin, X. Prezygotic and postzygotic control of uniparental mitochondrial DNA inheritance in Cryptococcus neoformans. mBio 2013, 4, e00112–e00113. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Fu, C.; Ianiri, G.; Heitman, J. The pheromone and pheromone receptor mating-type locus is involved in controlling uniparental mitochondrial inheritance in Cryptococcus. Genetics 2020, 214, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Gyawali, R.; Lin, X. Mechanisms of uniparental mitochondrial DNA inheritance in Cryptococcus neoformans. Mycobiology 2011, 39, 235–242. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, Z.; Wilson, A.; Xu, J. Mitochondrial DNA inheritance in the human fungal pathogen Cryptococcus gattii. Fungal Genet. Biol. 2015, 75, 1–10. [Google Scholar] [CrossRef]
- Matha, A.R.; Lin, X. Current perspectives on uniparental mitochondrial inheritance in Cryptococcus neoformans. Pathogens 2020, 9, 743. [Google Scholar] [CrossRef]
- Hua, W.; Vogan, A.; Xu, J. Genotypic and phenotypic analyses of two “isogenic” strains of the human fungal pathogen Cryptococcus neoformans var. neoformans. Mycopathologia 2019, 184, 195–212. [Google Scholar] [CrossRef]


{kind=link}
| Pair Name | JEC20/21 | KN99 | KN3501 | KN433 | XL280 | AIR265 | ||||||
| Recipient Parent | B3502 | H99 | B3501 | NIH433 | XL280 | R265 | ||||||
| Serotype | D | A | D | D | D | B | ||||||
| Mating Type | a | α | a | α | a | α | a | α | a | α | a | α |
| Molecular Type | VNIV | VNIV | VNI | VNI | VNIV | VNIV | VNIV | VNIV | VNIV | VNIV | VGII | VGII |
| Self-filamentation | - | + | ˗ | ˗ | ˗ | ˗ | ˗ | ˗ | ++ | +++ | - | - |
| Bisexual Mating | +++ | +++ | ++ | ++ | +++ | +++ | +++ | +++ | +++ | +++ | + | + |
| Virulence (i.n.) | ˧ | ˧ | +++ | +++ | n/a | n/a | n/a | n/a | ++ | ++ | +++ | +++ |
| Virulence (i.v.) | + | +˧ | +++ | +++ | + | + | + | +˧ | +++ | +++ | +++ | +++ |
| Reference | [19] | [19] | [29] | [29] | [27] | [27] | [27] | [27] | [30] | [30] | [23] | [23] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chadwick, B.J.; Lin, X. On the History and Applications of Congenic Strains in Cryptococcus Research. Pathogens 2020, 9, 750. https://doi.org/10.3390/pathogens9090750
Chadwick BJ, Lin X. On the History and Applications of Congenic Strains in Cryptococcus Research. Pathogens. 2020; 9(9):750. https://doi.org/10.3390/pathogens9090750
Chicago/Turabian StyleChadwick, Benjamin J., and Xiaorong Lin. 2020. "On the History and Applications of Congenic Strains in Cryptococcus Research" Pathogens 9, no. 9: 750. https://doi.org/10.3390/pathogens9090750
APA StyleChadwick, B. J., & Lin, X. (2020). On the History and Applications of Congenic Strains in Cryptococcus Research. Pathogens, 9(9), 750. https://doi.org/10.3390/pathogens9090750