Naturally Acquired Humoral Immunity against Malaria Parasites in Non-Human Primates from the Brazilian Amazon, Cerrado and Atlantic Forest

 , , , , ,
, , , , ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
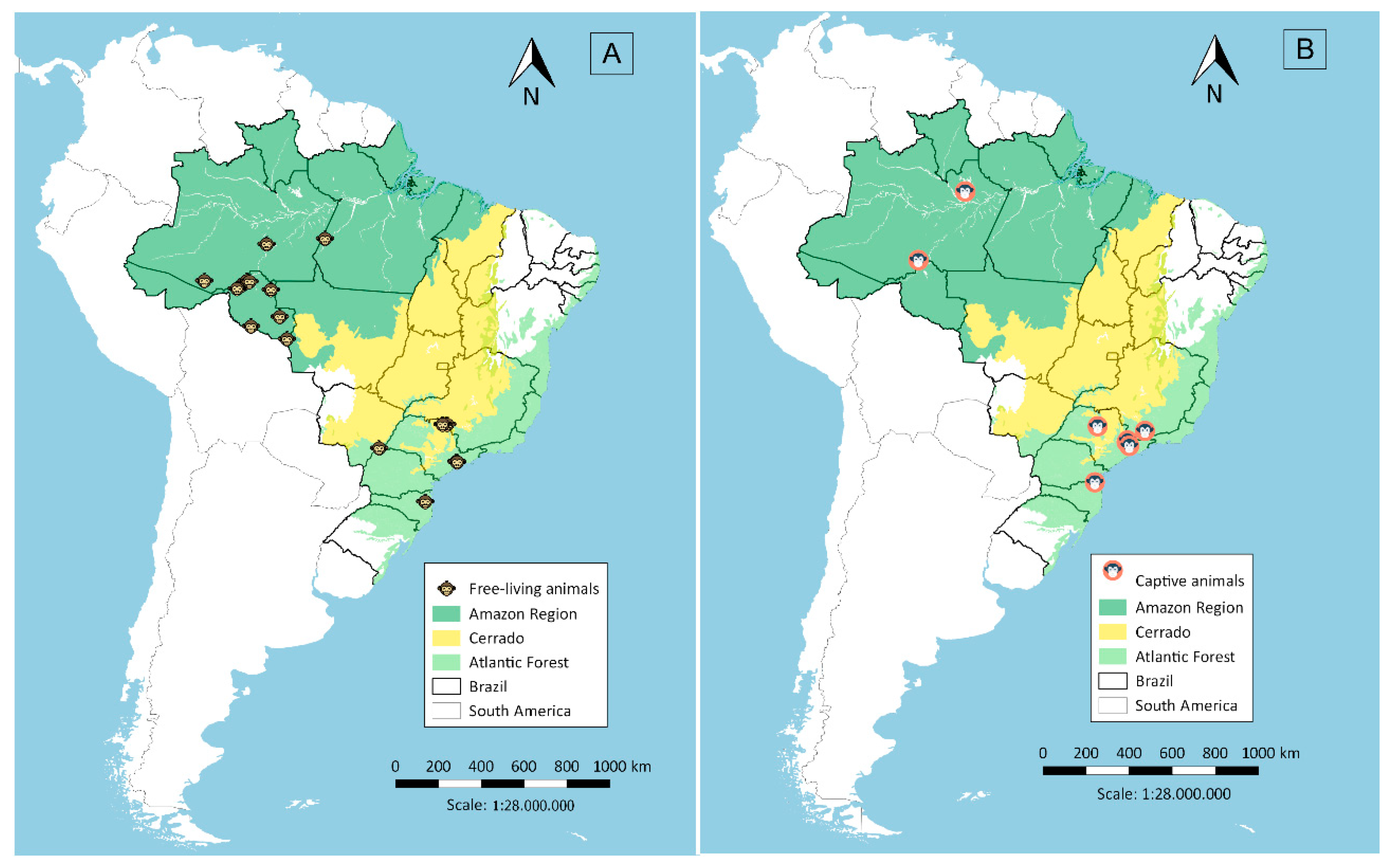
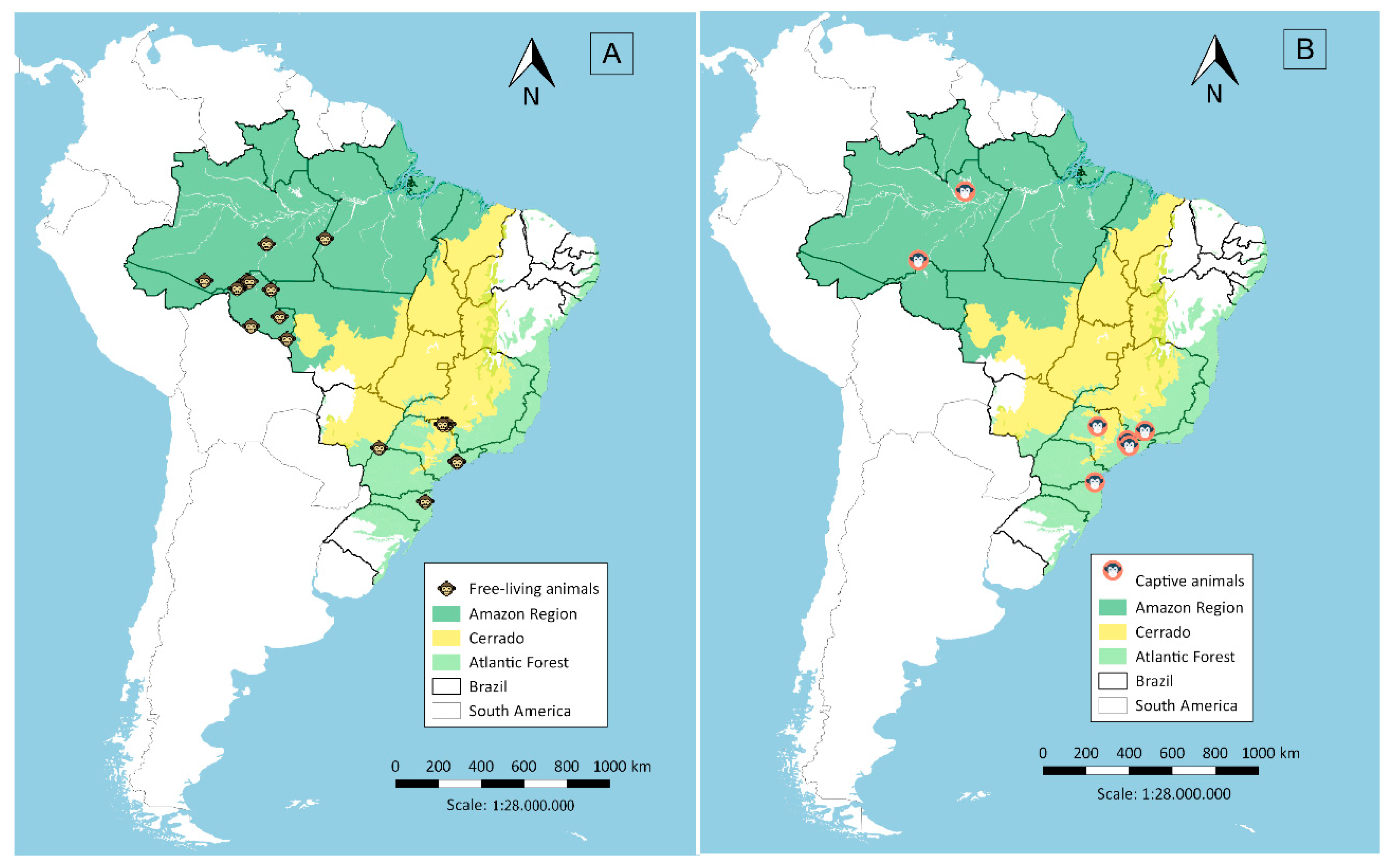
2.1. Sera Sampling from Non-Human Primates
2.2. Recombinant Antigens
2.3. Multiplexed Serological Assay
2.4. Assessment of Coupling Efficiency
3. Results
3.1. Testing of Coupling Efficiency of GST-Fusion Proteins to BioPlex Carboxylated Magnetic Beads
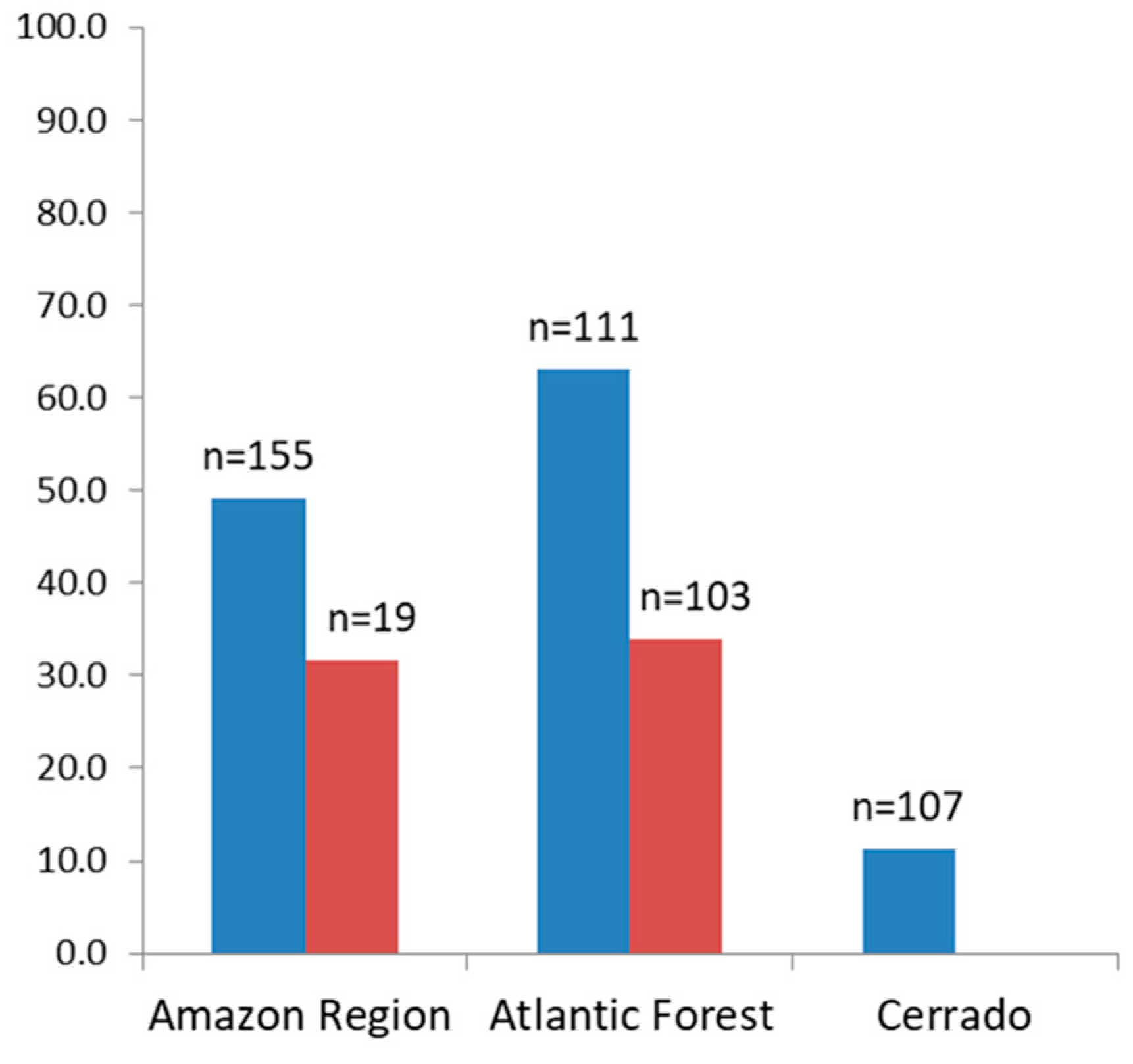
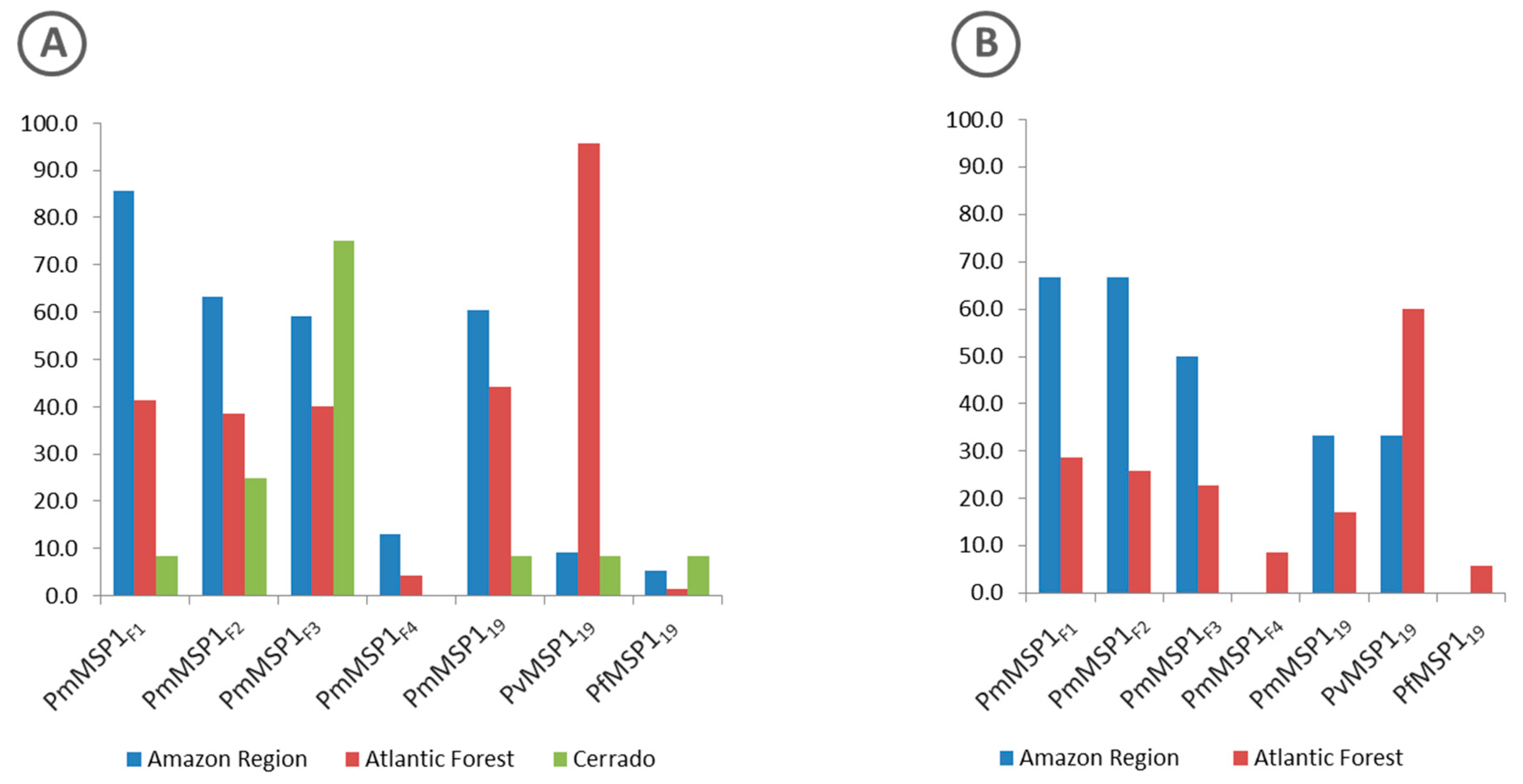
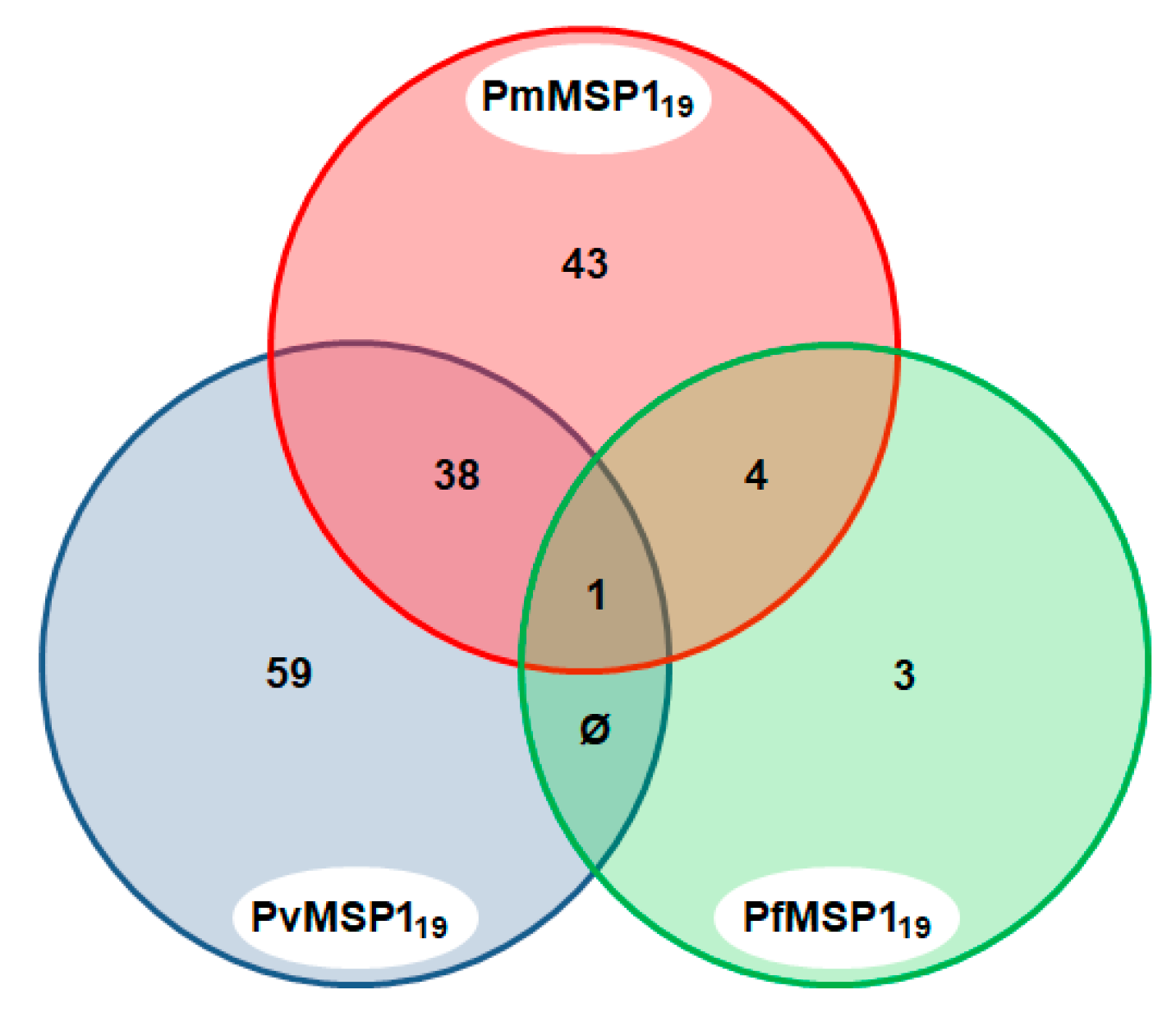
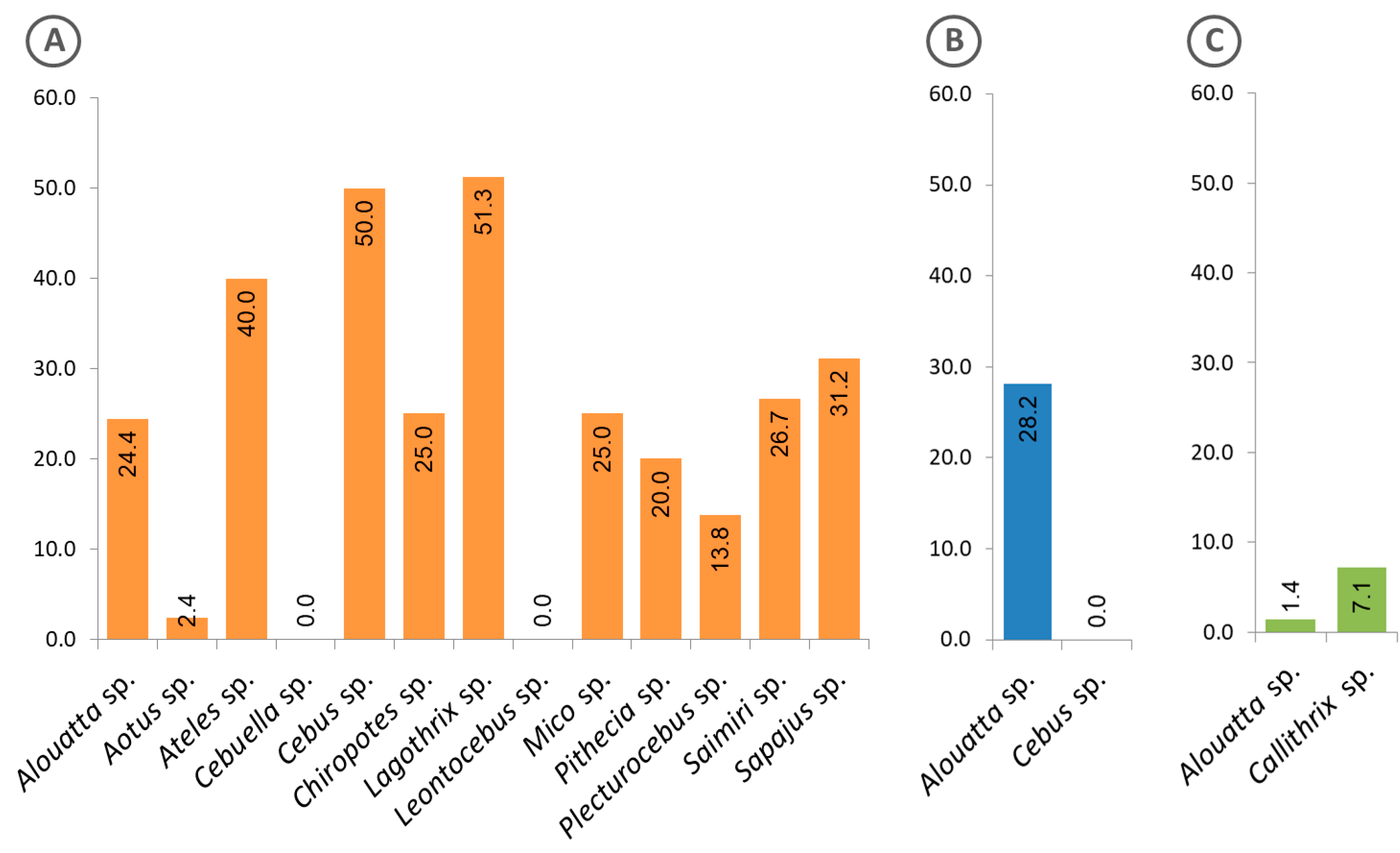
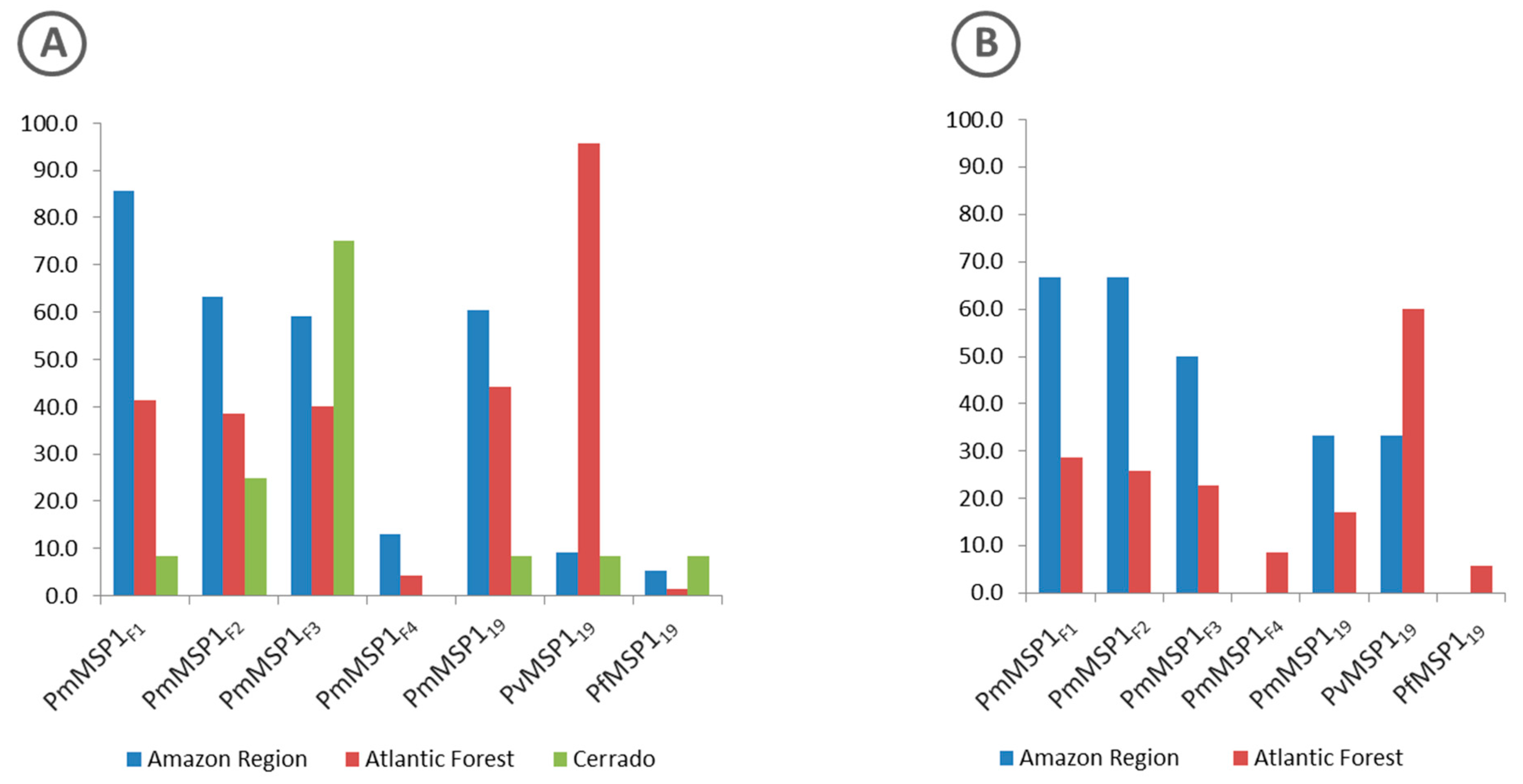
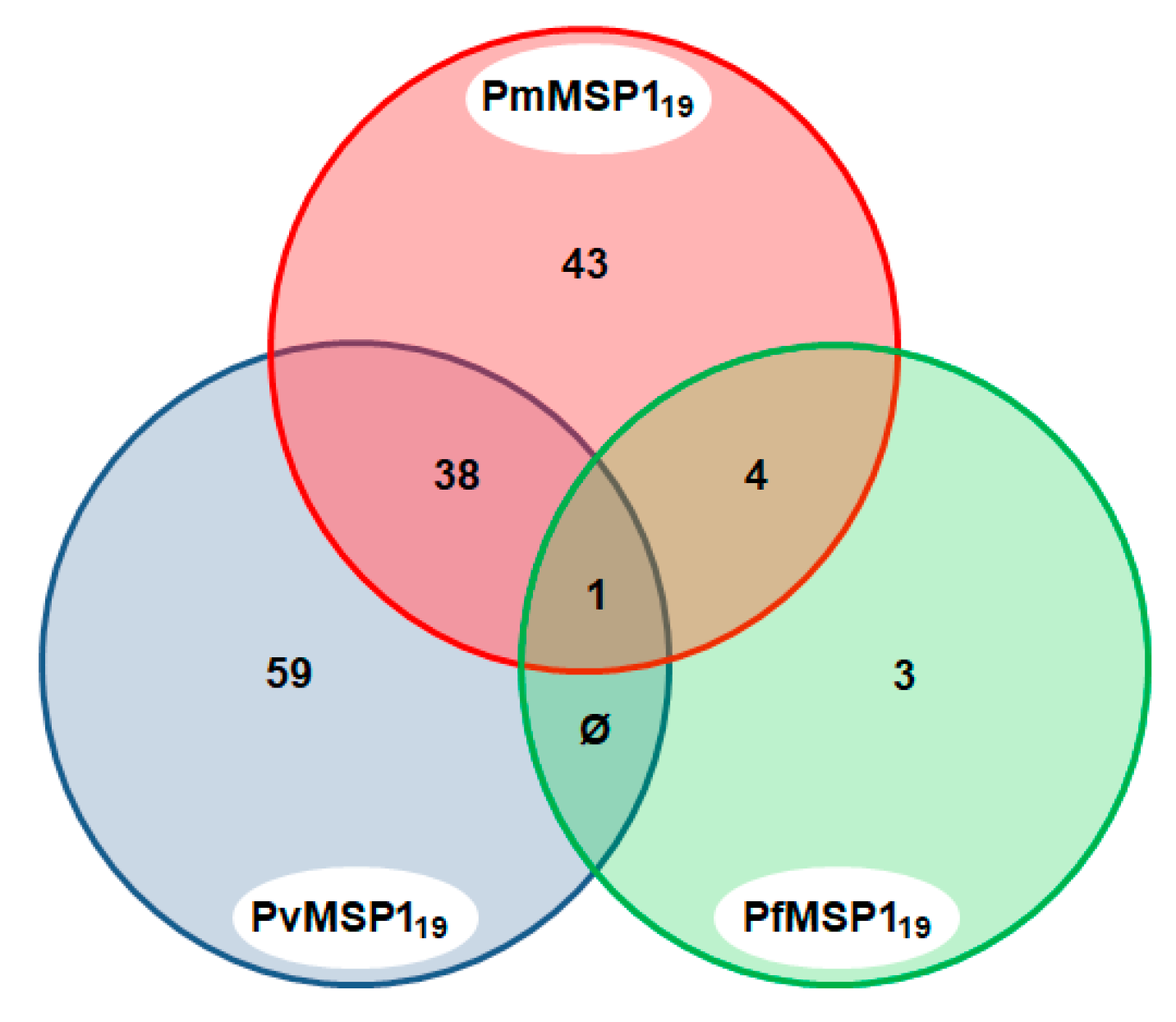
3.2. Antibodies Against MSP1 in the NHP Sera
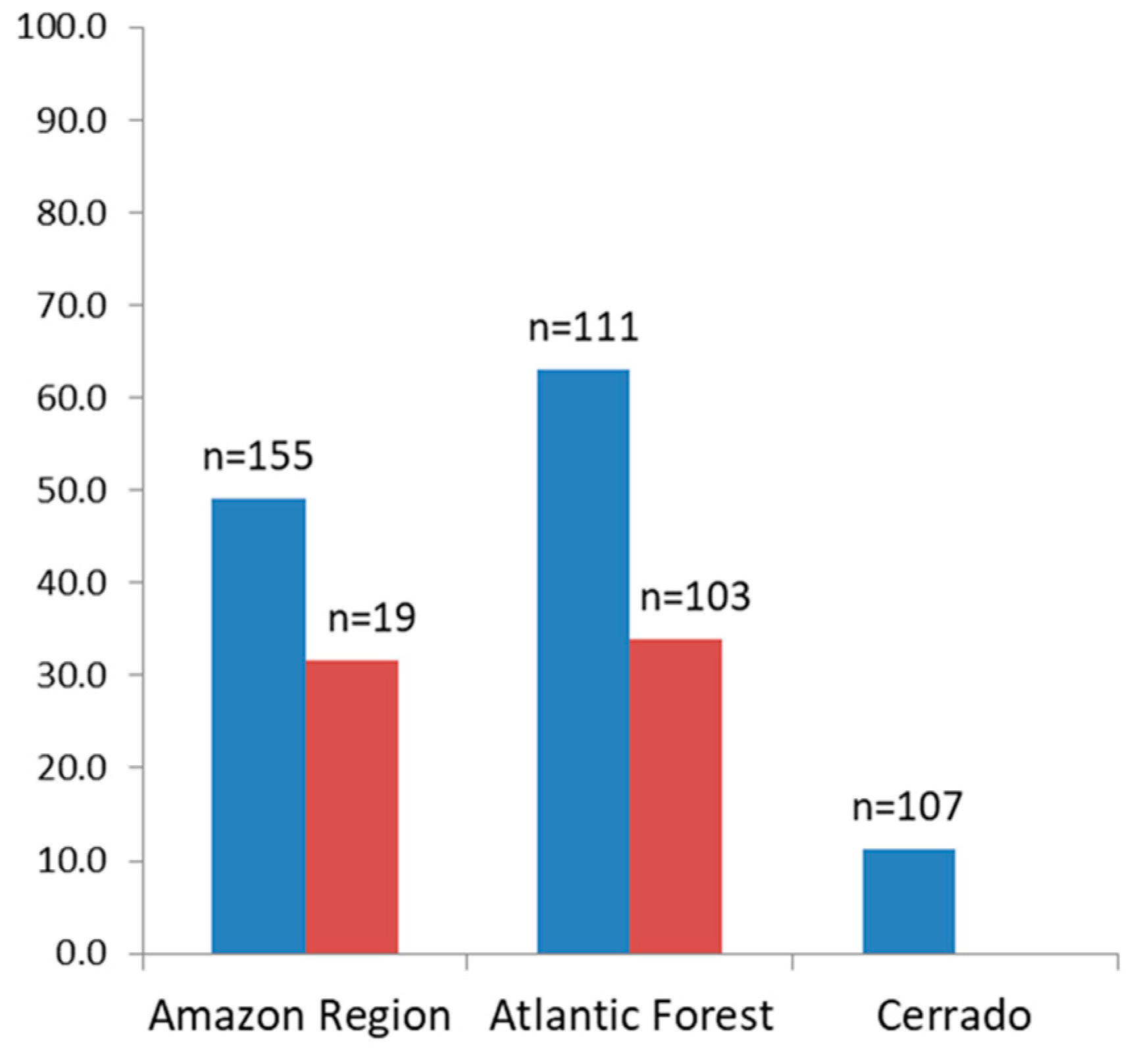
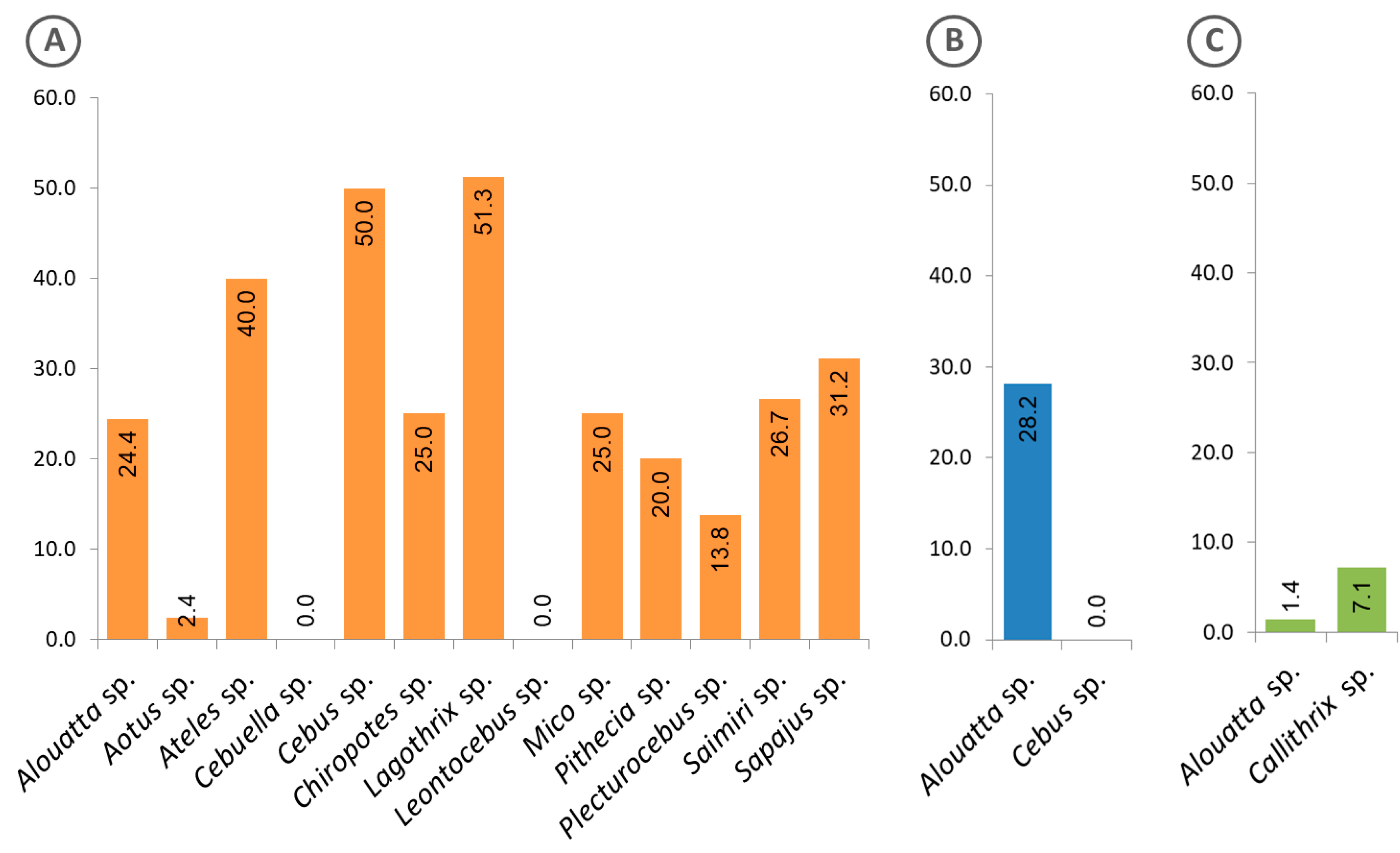
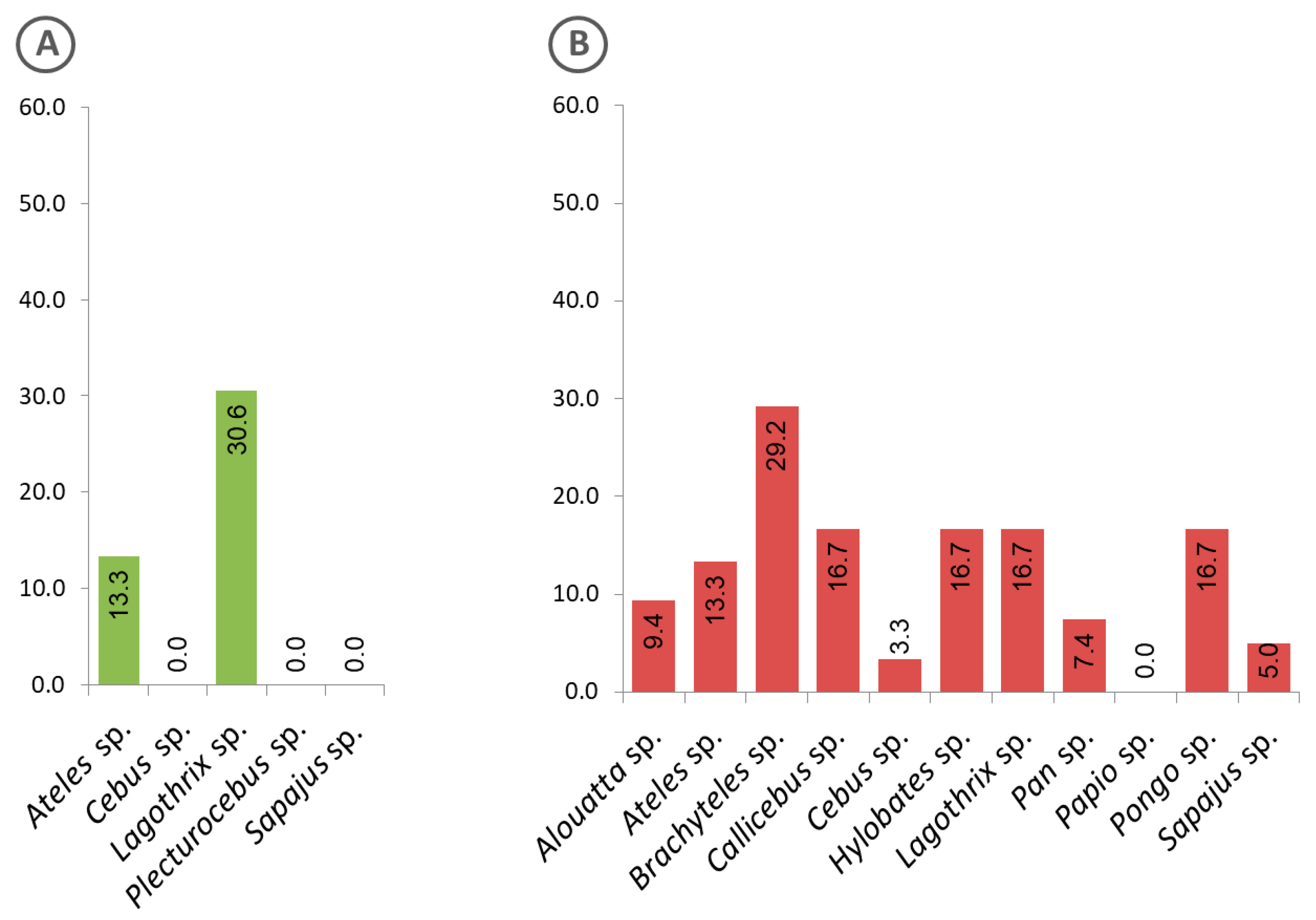
3.3. Malaria Exposure History
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holder, A.A. The carboxy-terminus of merozoite surface protein 1: Structure, specific antibodies and immunity to malaria. Parasitology 2009, 136, 1445–1456. [Google Scholar] [CrossRef] [PubMed]
- Holder, A.A.; Freeman, R.R. Biosynthesis and processing of a Plasmodium falciparum schizont antigen recognized by immune serum and a monoclonal antibody. J. Exp. Med. 1982, 156, 1528–1538. [Google Scholar] [CrossRef] [PubMed]
- Birkenmeyer, L.; Muerhoff, A.S.; Dawson, G.J.; Desai, S.M. Isolation and characterization of the MSP1 genes from Plasmodium malariae and Plasmodium ovale. Am. J. Trop. Med. Hyg. 2010, 82, 996–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, L.O.; Wunderlich, G.; Alves, J.M.; Bueno, M.G.; Röhe, F.; Catão-Dias, J.L.; Neves, A.; Malafronte, R.S.; Curado, I.; Domingues, W.; et al. Merozoite surface protein-1 genetic diversity in Plasmodium malariae and Plasmodium brasilianum from Brazil. BMC Infect. Dis. 2015, 15, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Healer, J.; Chiu, C.Y.; Hansen, D.S. Mechanisms of naturally acquired immunity to P. falciparum and approaches to identify merozoite antigen targets. Parasitology 2018, 145, 839–847. [Google Scholar] [CrossRef]
- Elizardez, Y.B.; Fotoran, W.L.; Junior, A.J.G.; Curado, I.; Junior, N.K.; Monteiro, E.F.; Romero Neto, I.; Wunderlich, G.; Kirchgatter, K. Recombinant proteins of Plasmodium malariae merozoite surface protein 1 (PmMSP1): Testing immunogenicity in the BALB/c model and potential use as diagnostic tool. PLoS ONE 2019, 14, e0219629. [Google Scholar] [CrossRef] [Green Version]
- Araújo, M.S.; Messias, M.R.; Figueiró, M.R.; Gil, L.H.; Probst, C.M.; Vidal, N.M.; Katsuragawa, T.H.; Krieger, M.A.; Silva, L.H.; Ozaki, L.S. Natural Plasmodium infection in monkeys in the state of Rondônia (Brazilian Western Amazon). Malar. J. 2013, 12, 180. [Google Scholar] [CrossRef] [Green Version]
- Bueno, M.G.; Rohe, F.; Kirchgatter, K.; Di Santi, S.M.; Guimarães, L.O.; Witte, C.L.; Costa-Nascimento, M.J.; Toniolo, C.R.; Catão-Dias, J.L. Survey of Plasmodium spp. in free-ranging neotropical primates from the Brazilian Amazon region impacted by anthropogenic actions. Ecohealth 2013, 10, 48–53. [Google Scholar] [CrossRef]
- Duarte, A.M.; Malafronte, R.; Cerutti, C., Jr.; Curado, I.; de Paiva, B.R.; Maeda, A.Y.; Yamasaki, T.; Summa, M.E.; Neves, D.; de Oliveira, S.G.; et al. Natural Plasmodium infections in Brazilian wild monkeys: Reservoirs for human infections? Acta Trop. 2008, 107, 179–185. [Google Scholar] [CrossRef]
- Duarte, A.M.; Porto, M.A.; Curado, I.; Malafronte, R.S.; Hoffmann, E.H.; de Oliveira, S.G.; da Silva, A.M.; Kloetzel, J.K.; Gomes, A. Widespread occurrence of antibodies against circumsporozoite protein and against blood forms of Plasmodium vivax, P. falciparum and P. malariae in Brazilian wild monkeys. J. Med. Primatol. 2006, 35, 87–96. [Google Scholar] [CrossRef]
- Fernandez-Becerra, C.; Sanz, S.; Brucet, M.; Stanisic, D.I.; Alves, F.P.; Camargo, E.P.; Alonso, P.L.; Mueller, I.; del Portillo, H.A. Naturally-acquired humoral immune responses against the N- and C-termini of the Plasmodium vivax MSP1 protein in endemic regions of Brazil and Papua New Guinea using a multiplex assay. Malar. J. 2010, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittermeier, R.A. Primate diversity and the tropical forest: Case studies from Brazil and Madagascar and the importance of the megadiversity countries. In Biodiversity; Wilson, E.O., Peter, F.M., Eds.; National Academy Press: Washington, DC, USA, 1988; pp. 145–154. [Google Scholar]
- Mittermeier, R.A.; Coimbra-Filho, A.F.; Kierulff, M.C.M.; Rylands, A.B.; Mendes, S.L.; Pissinatti, A.; Almeida, L.M. Monkeys of the Atlantic Forest of Eastern Brazil: Pocket Identification Guide; Conservation International Tropical Pocket Guide Series #3; Conservation International: Arlington, VA, USA, 2007. [Google Scholar]
- Gonder, R.; Von Berenberg-Gossler, H. Untersuchungen über malaria-plasmodien der affen. Malaria Intern. Archiv. (Leipzig). 1908, 1, 47–50. [Google Scholar]
- Fonseca, F. Plasmódio de primatas do Brasil. Mem. Inst. Oswaldo Cruz. 1951, 49, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, L.O.; Bajay, M.M.; Wunderlich, G.; Bueno, M.G.; Röhe, F.; Catão-Dias, J.L.; Neves, A.; Malafronte, R.S.; Curado, I.; Kirchgatter, K. The genetic diversity of Plasmodium malariae and Plasmodium brasilianum from human, simian and mosquito hosts in Brazil. Acta Trop. 2012, 124, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Bueno, M.G.; Catão-dias, J.L.; Laroque, P.O.; Vasconcellos, S.A.; Ferreira Neto, J.S.; Gennari, S.M.; Ferreira, F.; Laurenti, M.D.; Umezawa, E.S.; Kesper, N.; et al. Infectious Diseases In Free-ranging Blonde Capuchins, Sapajus Flavius, In Brazil. Int. J. Primatol. 2017, 38, 1017–1031. [Google Scholar] [CrossRef]
- Costa, D.C.; da Cunha, V.P.; de Assis, G.M.; de Souza Junior, J.C.; Hirano, Z.M.; de Arruda, M.E.; Kano, F.S.; Carvalho, L.H.; de Brito, C.F. Plasmodium simium/Plasmodium vivax infections in southern brown howler monkeys from the Atlantic Forest. Mem. Inst. Oswaldo Cruz 2014, 109, 641–653. [Google Scholar] [CrossRef]
- Alvarenga, D.A.; de Pina-Costa, A.; de Sousa, T.N.; Pissinatti, A.; Zalis, M.G.; Suaréz-Mutis, M.C.; Lourenço-de-Oliveira, R.; Brasil, P.; Daniel-Ribeiro, C.T.; de Brito, C.F. Simian malaria in the Brazilian Atlantic forest: First description of natural infection of capuchin monkeys (Cebinae subfamily) by Plasmodium simium. Malar. J. 2015, 14, 81. [Google Scholar] [CrossRef] [Green Version]
- Alvarenga, D.A.; Pina-Costa, A.; Bianco, C., Jr.; Moreira, S.B.; Brasil, P.; Pissinatti, A.; Daniel-Ribeiro, C.T.; Brito, C.F. New potential Plasmodium brasilianum hosts: Tamarin and marmoset monkeys (family Callitrichidae). Malar. J. 2017, 16, 71. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, M.A.; Di Santi, S.M.; Figueiredo, T.A.; Machado, R.Z. Natural Plasmodium infection in neotropical primates in the island of São Luís, state of Maranhão, Brazil. Rev. Bras. Parasitol. Vet. 2015, 24, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Assis, G.M.; Alvarenga, D.A.; Costa, D.C.; Souza, J.C., Jr.; Hirano, Z.M.; Kano, F.S.; Sousa, T.N.; Brito, C.F. Detection of Plasmodium in faeces of the New World primate Alouatta clamitans. Mem. Inst. Oswaldo Cruz. 2016, 111, 570–576. [Google Scholar] [CrossRef] [Green Version]
- Brasil, P.; Zalis, M.G.; de Pina-Costa, A.; Siqueira, A.M.; Júnior, C.B.; Silva, S.; Areas, A.L.L.; Pelajo-Machado, M.; de Alvarenga, D.A.M.; da Silva Santelli, A.C.F.; et al. Outbreak of human malaria caused by Plasmodium simium in the Atlantic Forest in Rio de Janeiro: A molecular epidemiological investigation. Lancet Glob. Health 2017, 5, E1038–E1046. [Google Scholar] [CrossRef] [Green Version]
- Silva, T.; Barros, F.; Bahia, M.; Sampaio Junior, F.D.; Santos, S.; Inoue, L.S.; Gonçalves, T.S.; Chiesorin Neto, L.; Faria, D.; Tochetto, C.; et al. Plasmodium vivax and Plasmodium falciparum infection in Neotropical primates in the western Amazon, Brazil. Zoonoses Public. Health 2019, 66, 798–804. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, T.; Duarte, A.M.; Curado, I.; Summa, M.E.; Neves, D.V.; Wunderlich, G.; Malafronte, R.S. Detection of etiological agents of malaria in howler monkeys from Atlantic Forests, rescued in regions of São Paulo city, Brazil. J. Med. Primatol. 2011, 40, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Soares, I.S.; Levitus, G.; Souza, J.M.; Del Portillo, H.A.; Rodrigues, M.M. Acquired immune responses to the N- and C-terminal regions of Plasmodium vivax merozoite surface protein 1 in individuals exposed to malaria. Infect. Immun. 1997, 65, 1606–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polley, S.D.; Tetteh, K.K.; Cavanagh, D.R.; Pearce, R.J.; Lloyd, J.M.; Bojang, K.A.; Okenu, D.M.; Greenwood, B.M.; McBride, J.S.; Conway, D.J. Repeat sequences in block 2 of Plasmodium falciparum merozoite surface protein 1 are targets of antibodies associated with protection from malaria. Infect. Immun. 2003, 71, 1833–1842. [Google Scholar] [CrossRef] [Green Version]
- Priest, J.W.; Plucinski, M.M.; Huber, C.S.; Rogier, E.; Mao, B.; Gregory, C.J.; Candrinho, B.; Colborn, J.; Barnwell, J.W. Specificity of the IgG antibody response to Plasmodium falciparum, Plasmodium vivax, Plasmodium malariae, and Plasmodium ovale MSP1(19) subunit proteins in multiplexed serologic assays. Malar. J. 2018, 17, 417. [Google Scholar] [CrossRef]
- Drakeley, C.J.; Corran, P.H.; Coleman, P.G.; Tongren, J.E.; McDonald, S.L.; Carneiro, I.; Malima, R.; Lusingu, J.; Manjurano, A.; Nkya, W.M.M.; et al. Estimating medium- and long-term trends in malaria transmission by using serological markers of malaria exposure. Proc. Natl. Acad. Sci. USA 2005, 102, 5108–5113. [Google Scholar] [CrossRef] [Green Version]
- Stewart, L.; Gosling, R.; Griffin, J.; Gesase, S.; Campo, J.; Hashim, R.; Masika, P.; Mosha, J.; Bousema, T.; Shekalaghe, S.; et al. Rapid assessment of malaria transmission using age-specific sero-conversion rates. PLoS ONE 2009, 4, e6083. [Google Scholar] [CrossRef] [Green Version]
- Buery, J.C.; Rodrigues, P.T.; Natal, L.; Salla, L.C.; Loss, A.C.; Vicente, C.R.; Rezende, H.R.; Duarte, A.M.R.C.; Fux, B.; Malafronte, R.D.S.; et al. Mitochondrial genome of Plasmodium vivax/simium detected in an endemic region for malaria in the Atlantic Forest of Espírito Santo state, Brazil: Do mosquitoes, simians and humans harbour the same parasite? Malar. J. 2017, 16, 437. [Google Scholar] [CrossRef] [Green Version]
- Deane, L.M. Simian malaria in Brazil. Mem. Inst. Oswaldo Cruz 1992, 87, 1–20. [Google Scholar] [CrossRef]
- Abreu, F.V.S.; Santos, E.D.; Mello, A.R.L.; Gomes, L.R.; Alvarenga, D.A.M.; Gomes, M.Q.; Vargas, W.P.; Bianco-Júnior, C.; Pina-Costa, A.; Teixeira, D.S.; et al. Howler monkeys are the reservoir of malarial parasites causing zoonotic infections in the Atlantic forest of Rio de Janeiro. PLoS Negl. Trop. Dis. 2019, 13, e0007906. [Google Scholar] [CrossRef] [PubMed]
- Gómez, A.; Aguirre, A.A. Infectious diseases and the illegal wildlife trade. Ann. N. Y. Acad. Sci. 2008, 1149, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Kock, R.A.; Woodford, M.H.; Rossiter, P.B. Disease risk associated with the translocation of wildlife. Revue Scientifique et Technique 2010, 29, 329–350. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Malaria Species | Target Antigen | Signal Cut-Off Values (MFI) * |
|---|---|---|
| P. malariae | PmMSP1F1 | 252.6 |
| P. malariae | PmMSP1F2 | 625.5 |
| P. malariae | PmMSP1F3 | 465.0 |
| P. malariae | PmMSP1F4 | 351.5 |
| P. malariae | PmMSP119 | 473.4 |
| P. vivax | PvMSP119 | 424.2 |
| P. falciparum | PfMSP119 | 471.7 |
| Recombinant Protein | Positive (%) | |
|---|---|---|
| PmMSP1F1 | 109 | (22.0) |
| PmMSP1F2 | 91 | (18.4) |
| PmMSP1F3 | 93 | (18.8) |
| PmMSP1F4 | 16 | (3.2) |
| PmMSP119 | 86 | (17.4) |
| PvMSP119 | 98 | (19.8) |
| PfMSP119 | 8 | (1.6) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, E.F.; Fernandez-Becerra, C.; Araujo, M.d.S.; Messias, M.R.; Ozaki, L.S.; Duarte, A.M.R.d.C.; Bueno, M.G.; Catao-Dias, J.L.; Chagas, C.R.F.; Mathias, B.d.S.; et al. Naturally Acquired Humoral Immunity against Malaria Parasites in Non-Human Primates from the Brazilian Amazon, Cerrado and Atlantic Forest. Pathogens 2020, 9, 525. https://doi.org/10.3390/pathogens9070525
Monteiro EF, Fernandez-Becerra C, Araujo MdS, Messias MR, Ozaki LS, Duarte AMRdC, Bueno MG, Catao-Dias JL, Chagas CRF, Mathias BdS, et al. Naturally Acquired Humoral Immunity against Malaria Parasites in Non-Human Primates from the Brazilian Amazon, Cerrado and Atlantic Forest. Pathogens. 2020; 9(7):525. https://doi.org/10.3390/pathogens9070525
Chicago/Turabian StyleMonteiro, Eliana Ferreira, Carmen Fernandez-Becerra, Maisa da Silva Araujo, Mariluce Rezende Messias, Luiz Shozo Ozaki, Ana Maria Ribeiro de Castro Duarte, Marina Galvão Bueno, Jose Luiz Catao-Dias, Carolina Romeiro Fernandes Chagas, Bruno da Silva Mathias, and et al. 2020. "Naturally Acquired Humoral Immunity against Malaria Parasites in Non-Human Primates from the Brazilian Amazon, Cerrado and Atlantic Forest" Pathogens 9, no. 7: 525. https://doi.org/10.3390/pathogens9070525
APA StyleMonteiro, E. F., Fernandez-Becerra, C., Araujo, M. d. S., Messias, M. R., Ozaki, L. S., Duarte, A. M. R. d. C., Bueno, M. G., Catao-Dias, J. L., Chagas, C. R. F., Mathias, B. d. S., Santos, M. G. d., Santos, S. V., Holcman, M. M., Souza, J. C. d., Jr., & Kirchgatter, K. (2020). Naturally Acquired Humoral Immunity against Malaria Parasites in Non-Human Primates from the Brazilian Amazon, Cerrado and Atlantic Forest. Pathogens, 9(7), 525. https://doi.org/10.3390/pathogens9070525