In Vitro Incorporation of Helicobacter pylori into Candida albicans Caused by Acidic pH Stress


 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
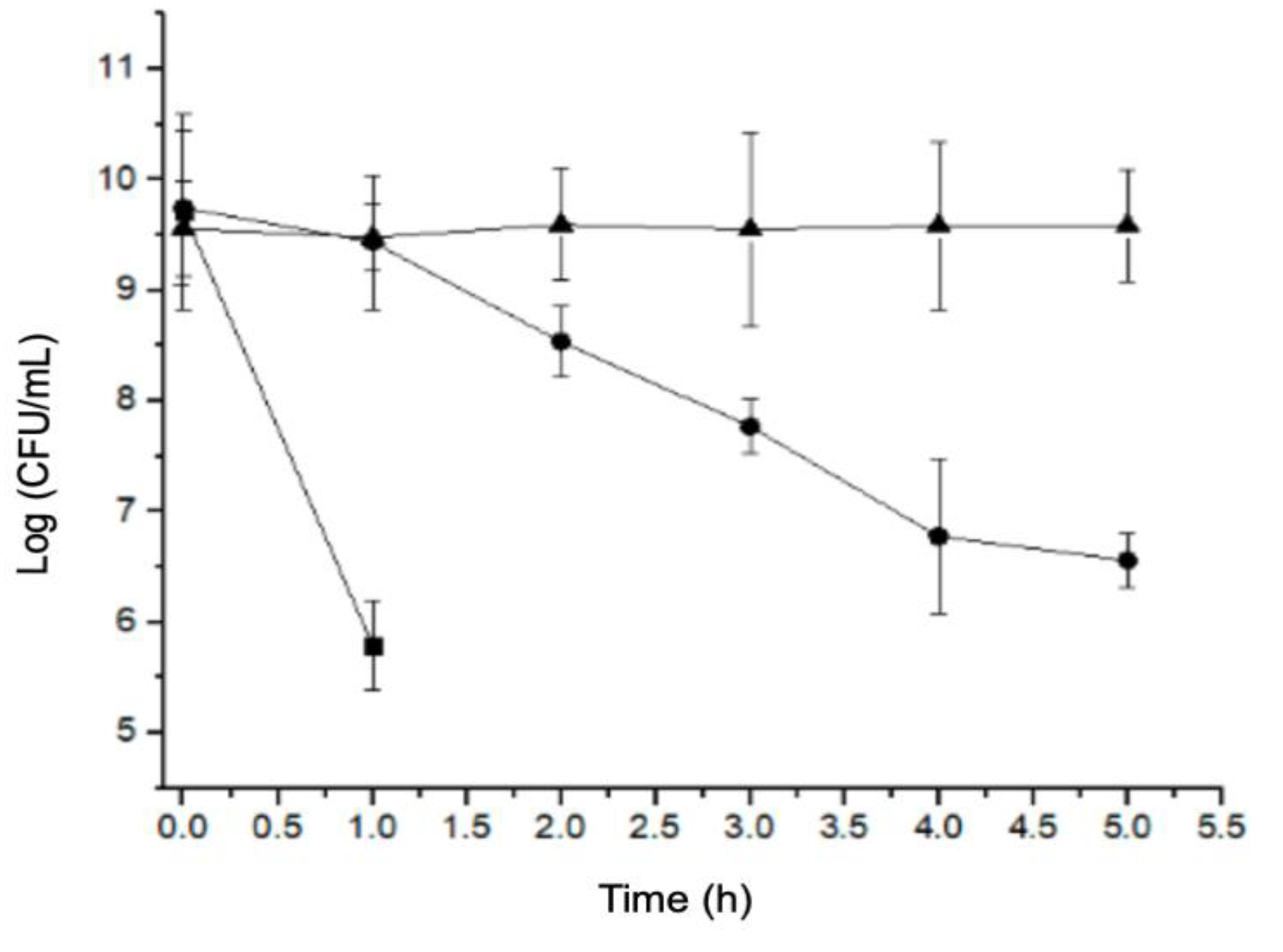
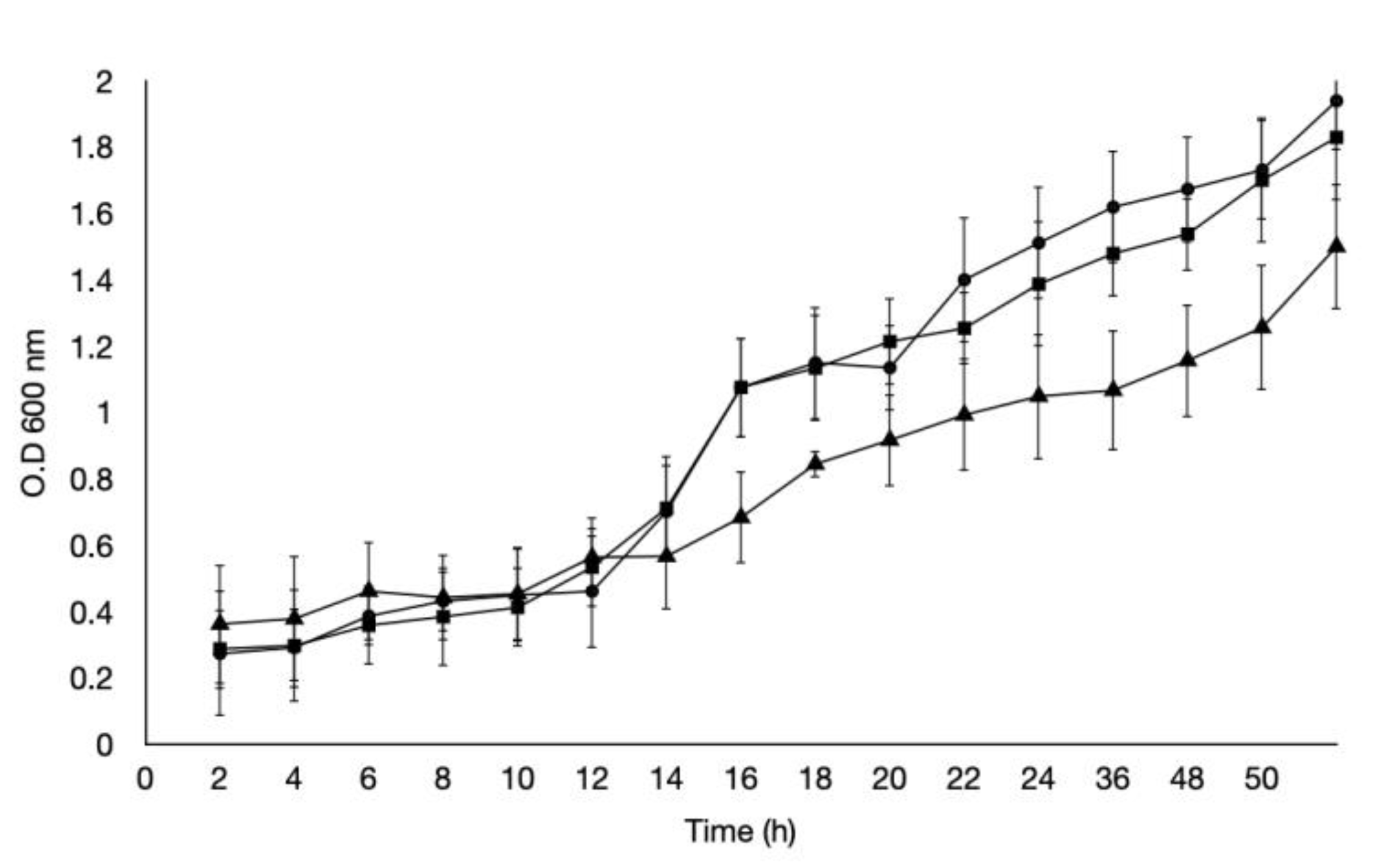
2.1. Acidic pH Stress Factor Assay
2.2. In Vitro Entry of H. pylori J99 into C. albicans ATCC 90028
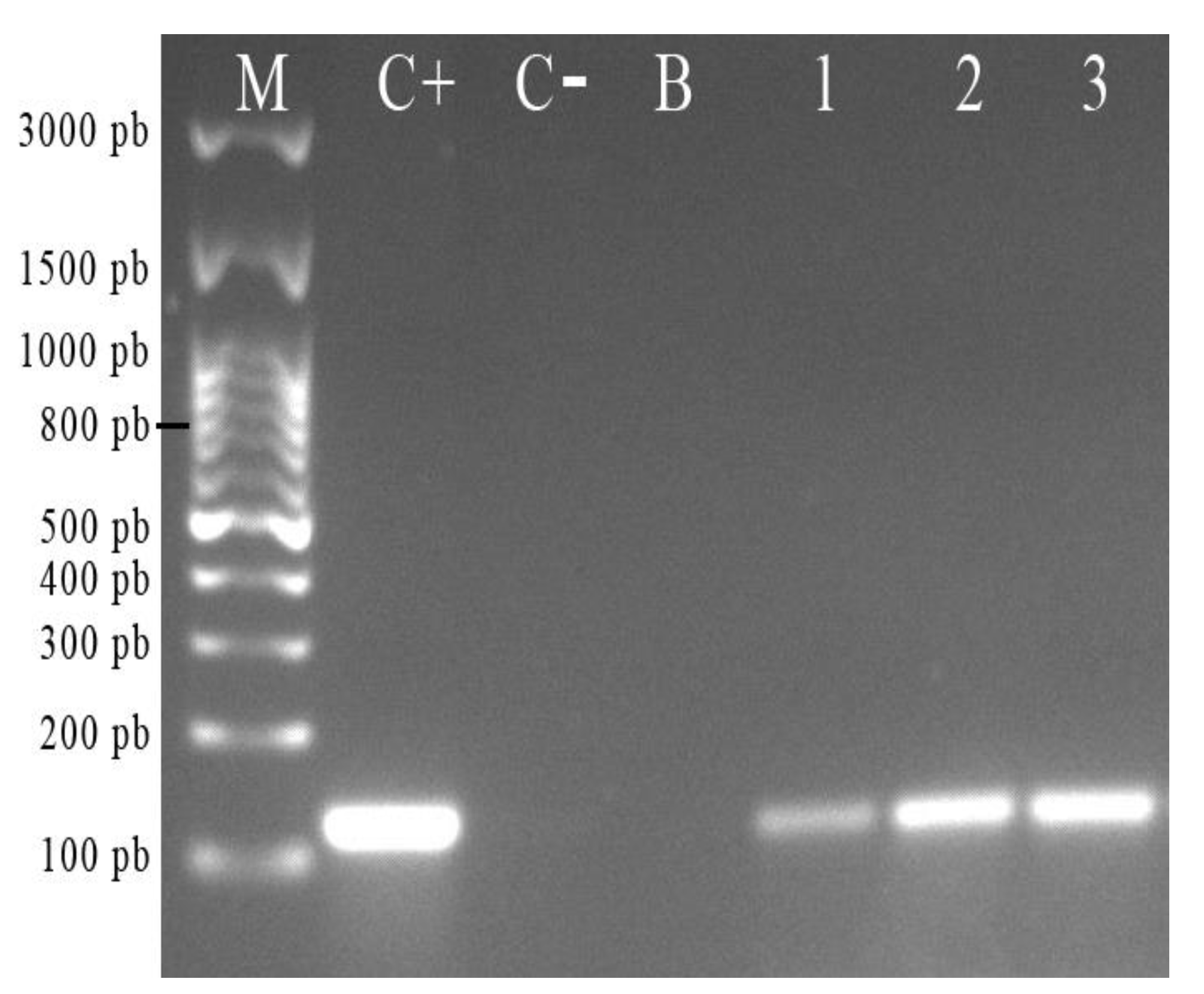
2.3. Amplification of The H. pylori J99 16S rDNA Gene from the Total DNA Extracted from C. albicans ATCC 90028
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Exposure to Acidic pH Stress Assay
4.3. In Vitro Assessment of H. pylori J99 Entry into C. albicans ATCC 90028 Caused by Acidic pH
4.4. Detection of H. pylori J99 Entry into C. albicans ATCC 90028 by Immunofluorescence
4.5. Amplification of The H. pylori 16S rDNA Gene from The Total DNA Extracted from C. albicans ATCC 90028
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Berk, S.G.; Ting, R.S.; Turner, G.W.; Ashburn, R.J. Production of respirable vesicles containing live Legionella pneumophila cells by two Acanthamoeba spp. Appl. Environ. Microbiol. 1998, 64, 279–286. [Google Scholar] [CrossRef] [PubMed]
- McFall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.G.; Carey, H.V.; Domazet-Lošo, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F.; et al. Animals in a bacterial world, a new imperative for the life sciences. Proc. Nat. Acad. Sci. USA 2013, 110, 3229–3236. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M.; Place, D.E.; Kuriakose, T.; Kanneganti, T.D. Interferon-inducible guanylate-binding proteins at the interface of cell-autonomous immunity and inflammasome activation. J. Leukoc. Biol. 2017, 101, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Siavoshi, F.; Heydari, S.; Shafiee, M.; Ahmadi, S.; Saniee, P.; Sarrafnejad, A.; Kolahdoozan, S. Sequestration inside the yeast vacuole may enhance Helicobacter pylori survival against stressful condition. Infect. Genet. Evol. 2019, 69, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Gasch, A.P.; Werner-Washburne, M. The genomics of yeast responses to environmental stress and starvation. Funct. Integr. Genomics 2002, 2, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Heydari, S.; Siavoshi, F.; Ebrahimi, H.; Sarrafnejad, A.; Sharifi, A.H. Excision of endosymbiotic bacteria from yeast under aging and starvation stresses. Infect. Genet. Evol. 2020, 78, 104141. [Google Scholar] [CrossRef] [PubMed]
- Siavoshi, F.; Salmanian, A.H.; Akbari, F.; Malekzadeh, R.; Massarrat, S. Detection of Helicobacter pylori-specific genes in the oral yeast. Helicobacter 2005, 10, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Salmanian, A.H.; Siavoshi, F.; Akbari, F.; Afshari, A.; Malekzadeh, R. Yeast of the oral cavity is the reservoir of Heliobacter pylori. J. Oral Pathol. Med 2008, 37, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Saniee, P.; Siavoshi, F.; Nikbakht Broujeni, G.; Khormali, M.; Sarrafnejad, A.; Malekzadeh, R. Immunodetection of Helicobacter pylori-specific proteins in oral and gastric Candida yeasts. Arch. Iran. Med. 2013, 16, 624–630. [Google Scholar] [PubMed]
- Saniee, P.; Siavoshi, F.; Nikbakht Broujeni, G.; Khormali, M.; Sarrafnejad, A.; Malekzadeh, R. Localization of H.pylori within the vacuole of Candida yeast by direct immunofluorescence technique. Arch. Iran. Med. 2013, 16, 705–710. [Google Scholar] [PubMed]
- Nett, J.E. Special Issue: Candida and Candidiasis. J. Fungi (Basel) 2018, 4, 74. [Google Scholar] [CrossRef] [PubMed]
- Zisova, L.G.; Chokoeva, A.A.; Amaliev, G.I.; Petleshkova, P.V.; Miteva-Katrandzhieva, T.; Krasteva, M.B.; Uchikova, E.H.; Kouzmanov, A.H.; Ivanova, Z.V. Vulvovaginal Candidiasis in Pregnant Women and its Importance for Candida Colonization of Newborns. Folia. Med. (Plovdiv) 2016, 58, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.; Sobel, J.D.; Nyirjesy, P.; Sobel, R.; Williams, V.L.; Yu, Q.; Noverr, M.C.; Fidel, P.L., Jr. Current patient perspectives of vulvovaginal candidiasis: Incidence, symptoms, management and post-treatment outcomes. BMC Womens Health 2019, 19, 48. [Google Scholar] [CrossRef] [PubMed]
- Miramón, P.; Lorenz, M.C. A feast for Candida: Metabolic plasticity confers an edge for virulence. PLoS Pathog. 2017, 13, e1006144. [Google Scholar] [CrossRef] [PubMed]
- Sherrington, S.L.; Sorsby, E.; Mahtey, N.; Kumwenda, P.; Lenardon, M.D.; Brown, I.; Ballou, E.R.; MacCallum, D.M.; Hall, R.A. Adaptation of Candida albicans to environmental pH induces cell wall remodelling and enhances innate immune recognition. PLoS Pathog. 2017, 13, e1006403. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C. Candida infections: An overview. Crit. Rev. Microbiol. 1987, 15, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Soll, D.R.; Galask, R.; Schmid, J.; Hanna, C.; Mac, K.; Morrow, B. Genetic dissimilarity of commensal strains of Candida spp. carried in different anatomical locations of the same healthy women. J. Clin. Microbiol. 1991, 29, 1702–1710. [Google Scholar] [CrossRef]
- Soll, D.R. Candida commensalism and virulence: The evolution of phenotypic plasticity. Acta. Trop. 2002, 81, 101–110. [Google Scholar] [CrossRef]
- Moreno-Mesonero, L.; Moreno, Y.; Alonso, J.L.; Ferrús, M.A. DVC-FISH and PMA-qPCR techniques to assess the survival of Helicobacter pylori inside Acanthamoeba castellanii. Res. Microbiol. 2016, 167, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Siavoshi, F.; Nourali-Ahari, F. Yeast protects Helicobacter pylori against the environmental stress. Arch. Iran. Med. 1998, 1, 2–821. [Google Scholar]
- Salmanian, A.-H.; Siavoshi, F.; Beyrami, Z.; Latifi-Navid, S.; Tavakolian, A.; Sadjadi, A. Foodborne yeasts serve as reservoirs of Helicobacter pylori. J. Food Safety 2012, 32, 152–160. [Google Scholar] [CrossRef]
- Matamala-Valdés, L.; Sánchez-Alonzo, K.; Parra, C.; Sáez, K.; Aguayo-Reyes, A.; García, A. Detection of intracellular Helicobacter pylori in Candida. SPP from neonate oral swabs. Rev. Assoc. Med. Bras. 2018, 64, 928–935. [Google Scholar] [CrossRef]
- Ince, A.T.; Kocaman, O.; Ismailova, M.; Tozlu, M.; Gücin, Z.; Iraz, M. A rare co-existence of Helicobacter pylori, Candida albicans and Candida keyfir in a giant gastric ulcer. Turk. J. Gastroenterol. 2014, 25, 435–436. [Google Scholar] [CrossRef] [PubMed]
- Pittet, D.; Li, N.; Wenzel, R.P. Association of secondary and polymicrobial nosocomial bloodstream infections with higher mortality. Eur. J. Clin. Microbiol. Infect. Dis. 1993, 12, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.; Salas-Jara, M.J.; Herrera, C.; Gonzalez, C. Biofilm and Helicobacter pylori: From environment to human host. World J. Gastroenterol. 2014, 20, 5632–5638. [Google Scholar] [CrossRef] [PubMed]
- Mladenova, I.; Durazzo, M. Transmission of Helicobacter pylori. Minerva. Gastroenterol. Dietol. 2018, 64, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Burucoa, C.; Axon, A. Epidemiology of Helicobacter pylori infection. Helicobacter 2017, 22 (Suppl. 1). [Google Scholar] [CrossRef] [PubMed]
- Mamishi, S.; Eshaghi, H.; Mahmoudi, S.; Bahador, A.; Hosseinpour Sadeghi, R.; Najafi, M.; Farahmand, F.; Khodadad, A.; Pourakbari, B. Intrafamilial transmission of Helicobacter pylori: Genotyping of faecal samples. Br. J. Biomed. Sci. 2016, 73, 38–43. [Google Scholar] [CrossRef] [PubMed]
- She, F.F.; Lin, J.Y.; Liu, J.Y.; Huang, C.; Su, D.H. Virulence of water-induced coccoid Helicobacter pylori and its experimental infection in mice. World J. Gastroenterol. 2003, 9, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, N.F.; Almeida, C.; Cerqueira, L.; Dias, S.; Keevil, C.W.; Vieira, M.J. Coccoid form of Helicobacter pylori as a morphological manifestation of cell adaptation to the environment. Appl. Environ. Microbiol. 2007, 73, 3423–3427. [Google Scholar] [CrossRef] [PubMed]
- Ansari, S.; Yamaoka, Y. Survival of Helicobacter pylori in gastric acidic territory. Helicobacter 2017, 22. [Google Scholar] [CrossRef]
- Suzuki, N.; Yoneda, M.; Naito, T.; Iwamoto, T.; Masuo, Y.; Yamada, K.; Hisama, K.; Okada, I.; Hirofuji, T. Detection of Helicobacter pylori DNA in the saliva of patients complaining of halitosis. J. Med. Microbiol. 2008, 57, 1553–1559. [Google Scholar] [CrossRef]
- Agarwal, S.; Jithendra, K.D. Presence of Helicobacter pylori in subgingival plaque of periodontitis patients with and without dyspepsia, detected by polymerase chain reaction and culture. J. Indian Soc. Periodontol. 2012, 16, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Siavoshi, F.; Sahraee, M.; Ebrahimi, H.; Sarrafnejad, A.; Saniee, P. Natural fruits, flowers, honey, and honeybees harbor Helicobacter pylori-positive yeasts. Helicobacter 2018, 23, e12471. [Google Scholar] [CrossRef] [PubMed]
- Vylkova, S. Environmental pH modulation by pathogenic fungi as a strategy to conquer the host. PLoS Pathogens. 2017, 13, e1006149. [Google Scholar] [CrossRef]
- Bahrami, A.R.; Rahimi, E.; Ghasemian Safaei, H. Detection of Helicobacter pylori in city water, dental units’ water, and bottled mineral water in Isfahan, Iran. Sci. World J. 2013, 2013, 280510. [Google Scholar] [CrossRef] [PubMed]
- Ahumada-Rudolph, R.; Cajas-Madriaga, D.; Rudolph, A.; Reinoso, R.; Torres, C.; Silva, M.; Becerra, J. Variation of sterols and fatty acids as an adaptive response to changes in temperature, salinity and pH of a marine fungus Epicoccum nigrum isolated from the Patagonian Fjords. Rev. Biol. Mar. Oceanogr. 2014, 49, 293–305. [Google Scholar] [CrossRef]
- Valle-González, E.R.; Jackman, J.A.; Yoon, B.K.; Park, S.; Sut, T.N.; Cho, N.J. Characterizing How Acidic pH Conditions Affect the Membrane-Disruptive Activities of Lauric Acid and Glycerol Monolaurate. Langmuir 2018, 34, 13745–13753. [Google Scholar] [CrossRef] [PubMed]
- Amieva, M.R.; Salama, N.R.; Tompkins, L.S.; Falkow, S. Helicobacter pylori enter and survive within multivesicular vacuoles of epithelial cells. Cell. Microbiol. 2002, 4, 677–690. [Google Scholar] [CrossRef]
- Tournu, H.; Carroll, J.; Latimer, B.; Dragoi, A.-M.; Dykes, S.; Cardelli, J.; Peters, T.L.; Eberle, K.E.; Palmer, G.E. Identification of small molecules that disrupt vacuolar function in the pathogen Candida albicans. PLoS ONE 2017, 12, e0171145. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Herman, P.K.; Emr, S.D. The fungal vacuole: Composition, function, and biogenesis. Microbiol. Rev. 1990, 54, 266–292. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (h) | pH 3 | pH 4 | pH 7 | p-Value |
|---|---|---|---|---|
| 0 | - | - | - | |
| 1 | 11.7% | 13.7% | 6.3% | 0.0559 |
| 3 | 3.0% | 3.3% | 2.7% | 0.9540 |
| 6 | 4.2% | 3.3% | 2.2% | 0.5773 |
| 12 | 7.1% | 6.4% | 2.2% | 0.0963 |
| 24 | 8.7% | 12.5% | 3.3% | 0.0031 |
| 48 | 11.1% | 18.7% | 4.1% | 0.0309 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Alonzo, K.; Parra-Sepúlveda, C.; Vega, S.; Bernasconi, H.; Campos, V.L.; Smith, C.T.; Sáez, K.; García-Cancino, A. In Vitro Incorporation of Helicobacter pylori into Candida albicans Caused by Acidic pH Stress. Pathogens 2020, 9, 489. https://doi.org/10.3390/pathogens9060489
Sánchez-Alonzo K, Parra-Sepúlveda C, Vega S, Bernasconi H, Campos VL, Smith CT, Sáez K, García-Cancino A. In Vitro Incorporation of Helicobacter pylori into Candida albicans Caused by Acidic pH Stress. Pathogens. 2020; 9(6):489. https://doi.org/10.3390/pathogens9060489
Chicago/Turabian StyleSánchez-Alonzo, Kimberly, Cristian Parra-Sepúlveda, Samuel Vega, Humberto Bernasconi, Víctor L. Campos, Carlos T. Smith, Katia Sáez, and Apolinaria García-Cancino. 2020. "In Vitro Incorporation of Helicobacter pylori into Candida albicans Caused by Acidic pH Stress" Pathogens 9, no. 6: 489. https://doi.org/10.3390/pathogens9060489
APA StyleSánchez-Alonzo, K., Parra-Sepúlveda, C., Vega, S., Bernasconi, H., Campos, V. L., Smith, C. T., Sáez, K., & García-Cancino, A. (2020). In Vitro Incorporation of Helicobacter pylori into Candida albicans Caused by Acidic pH Stress. Pathogens, 9(6), 489. https://doi.org/10.3390/pathogens9060489