Reduced Antioxidant Response of the Fan Mussel Pinna nobilis Related to the Presence of Haplosporidium pinnae



 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
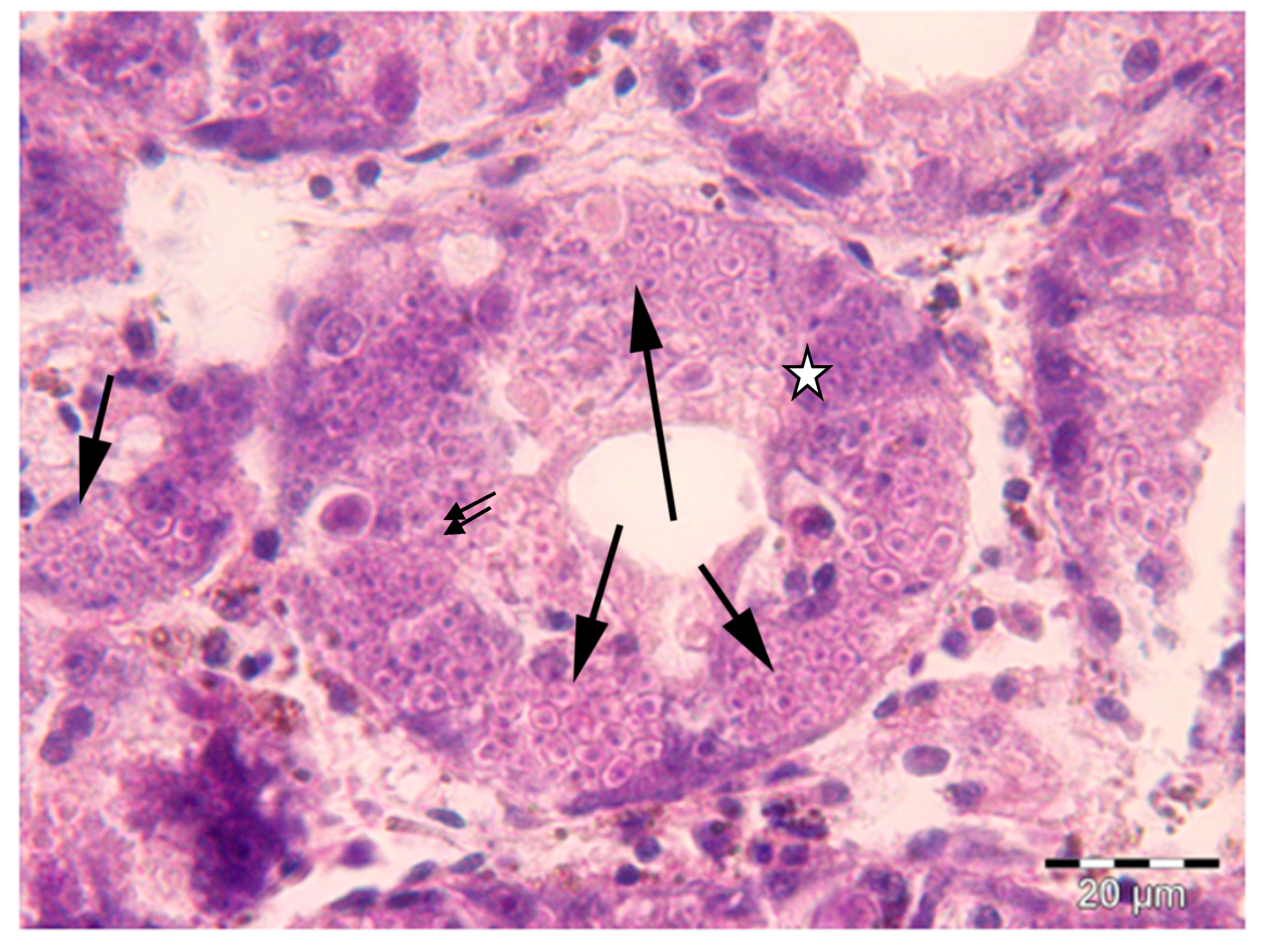
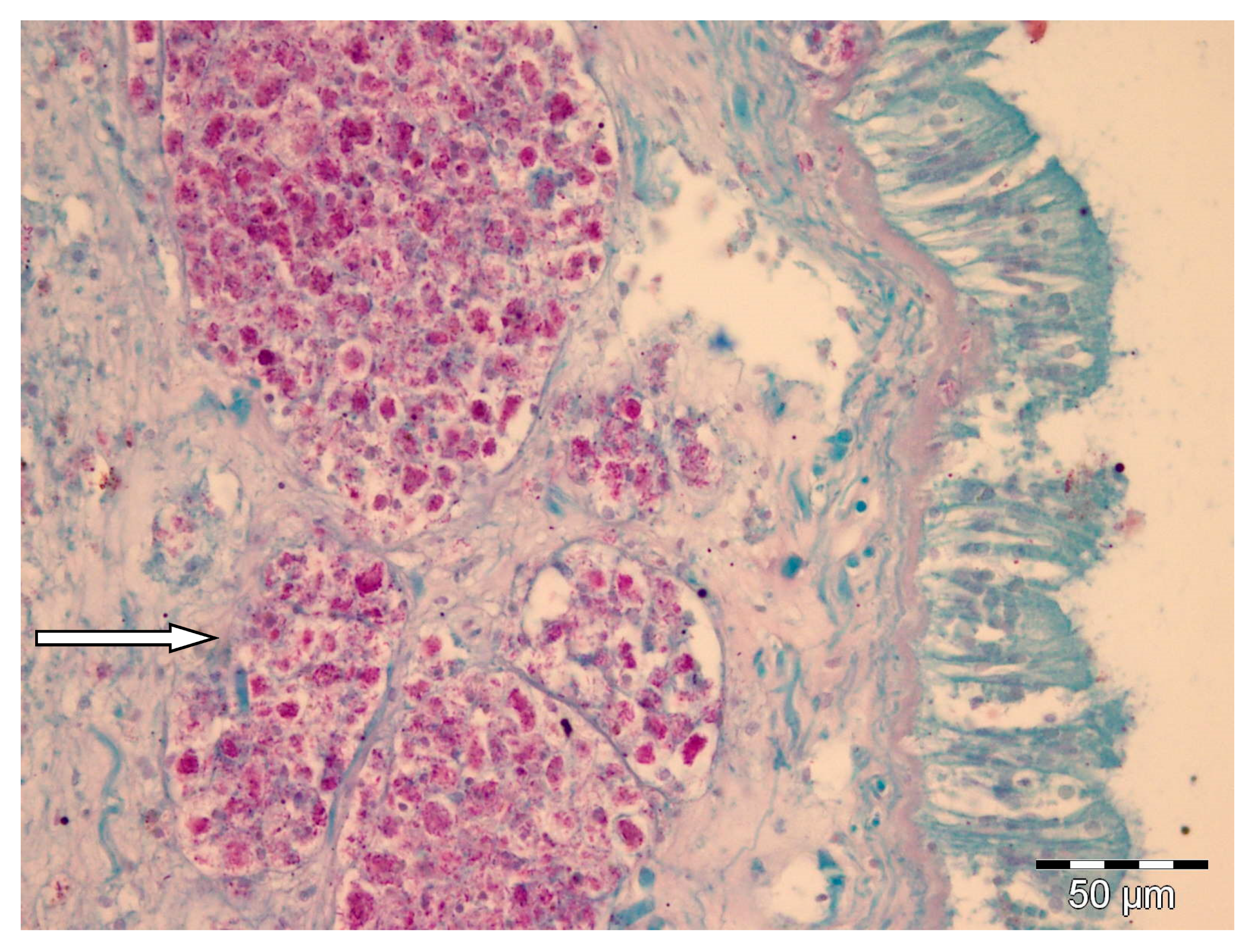
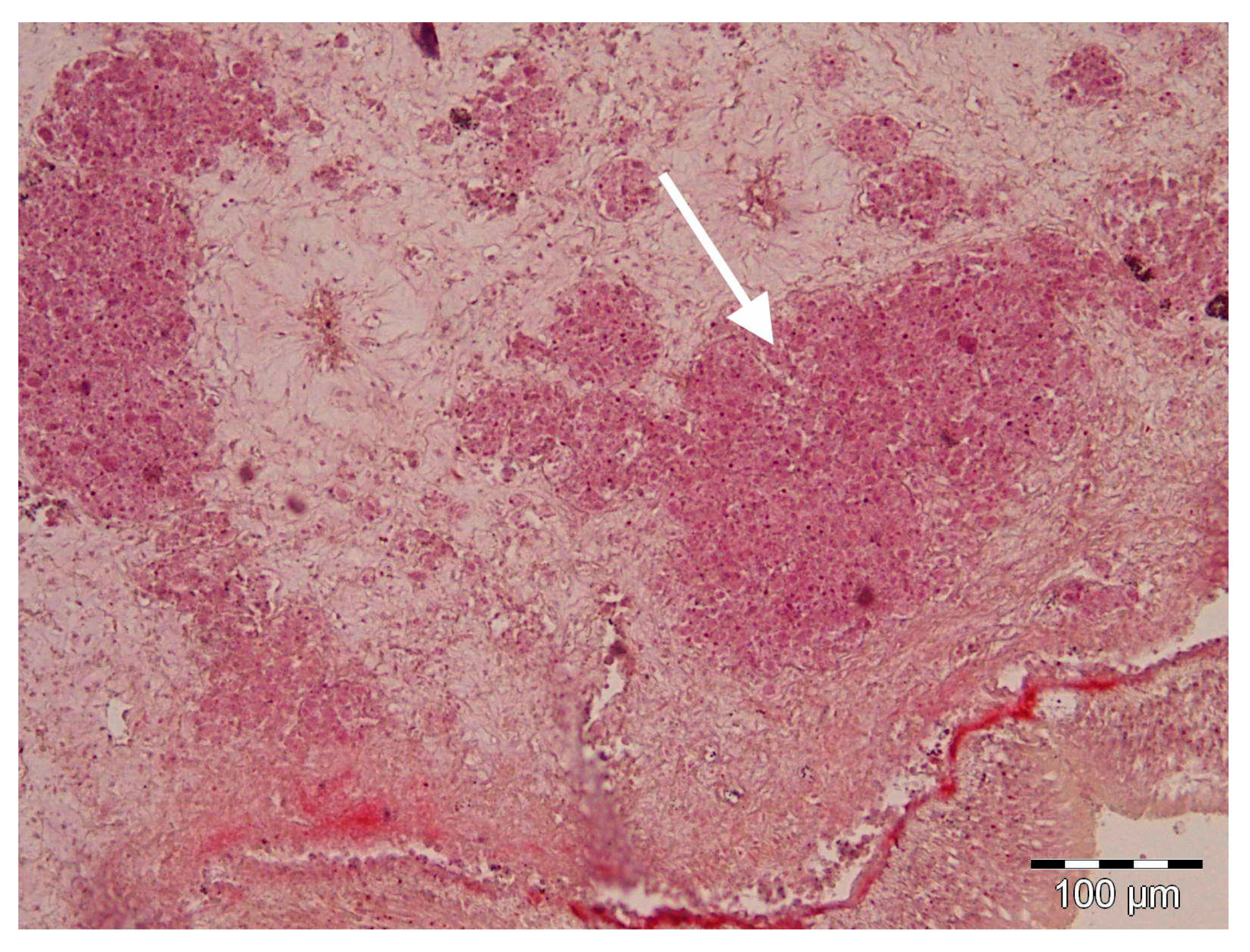
2.1. Histology and Molecular Biology Analyses
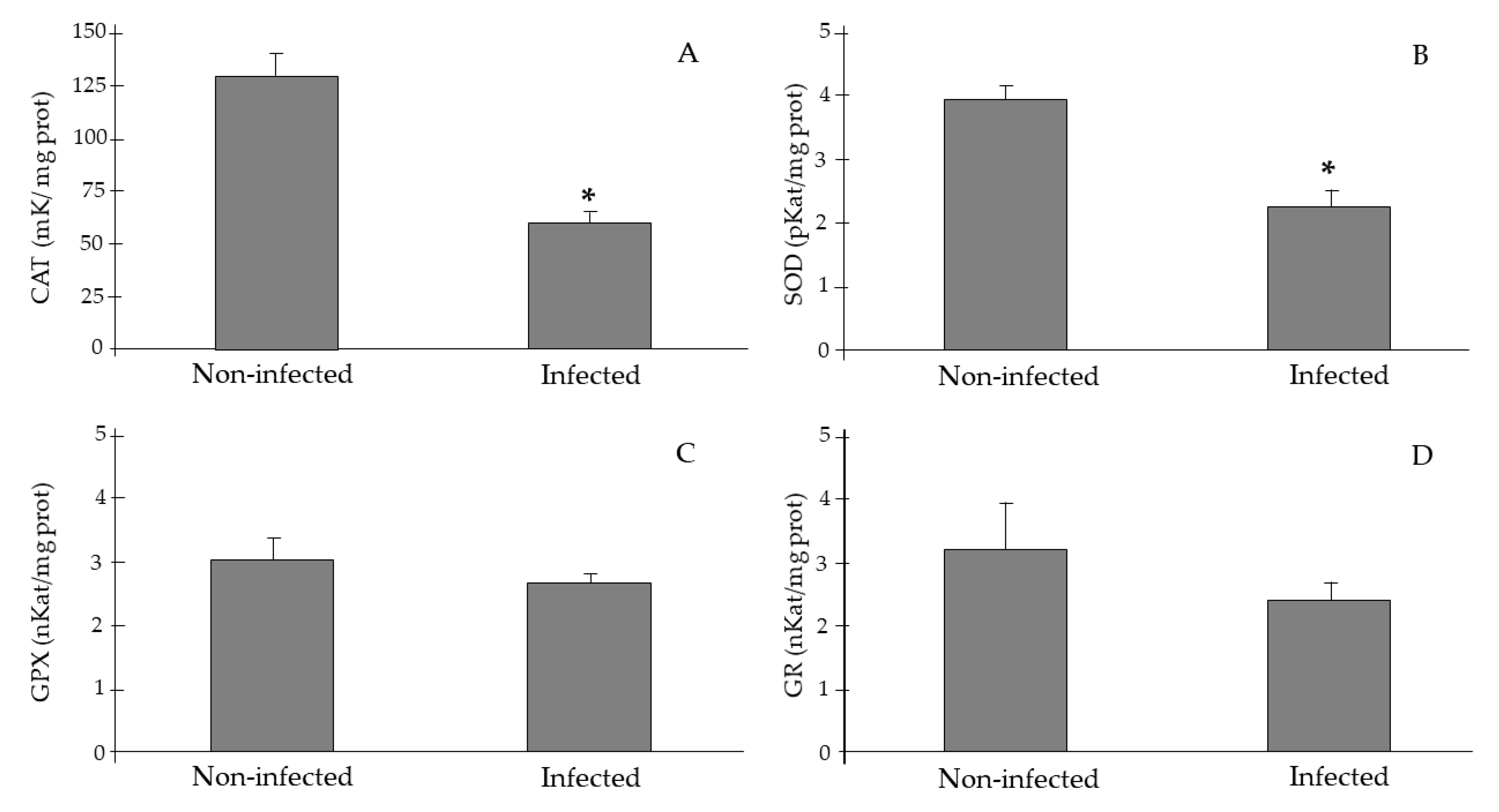
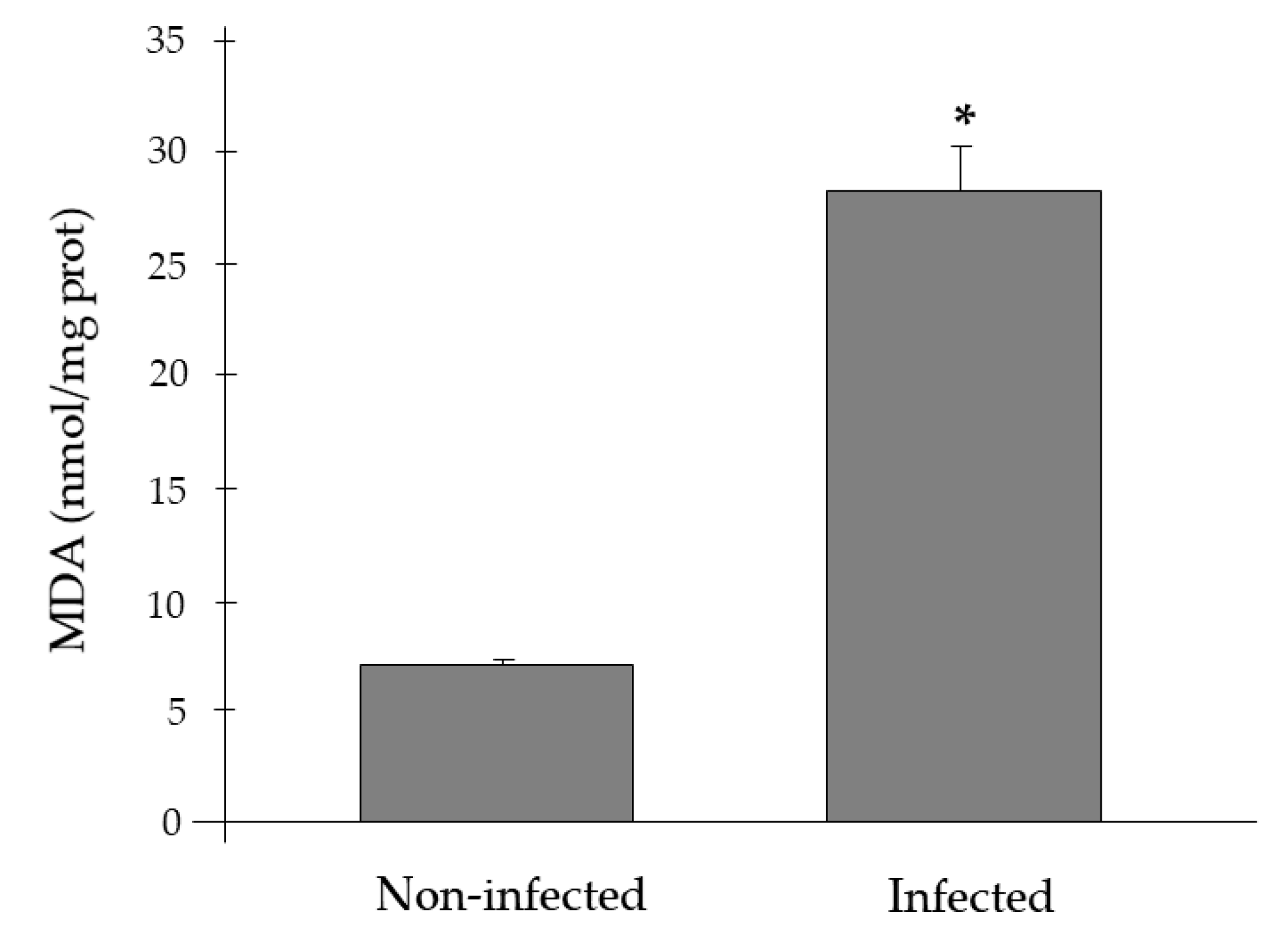
2.2. Antioxidant Activities and Lipid Peroxidation Biomarkers
3. Discussion
4. Conclusions
5. Materials and Methods
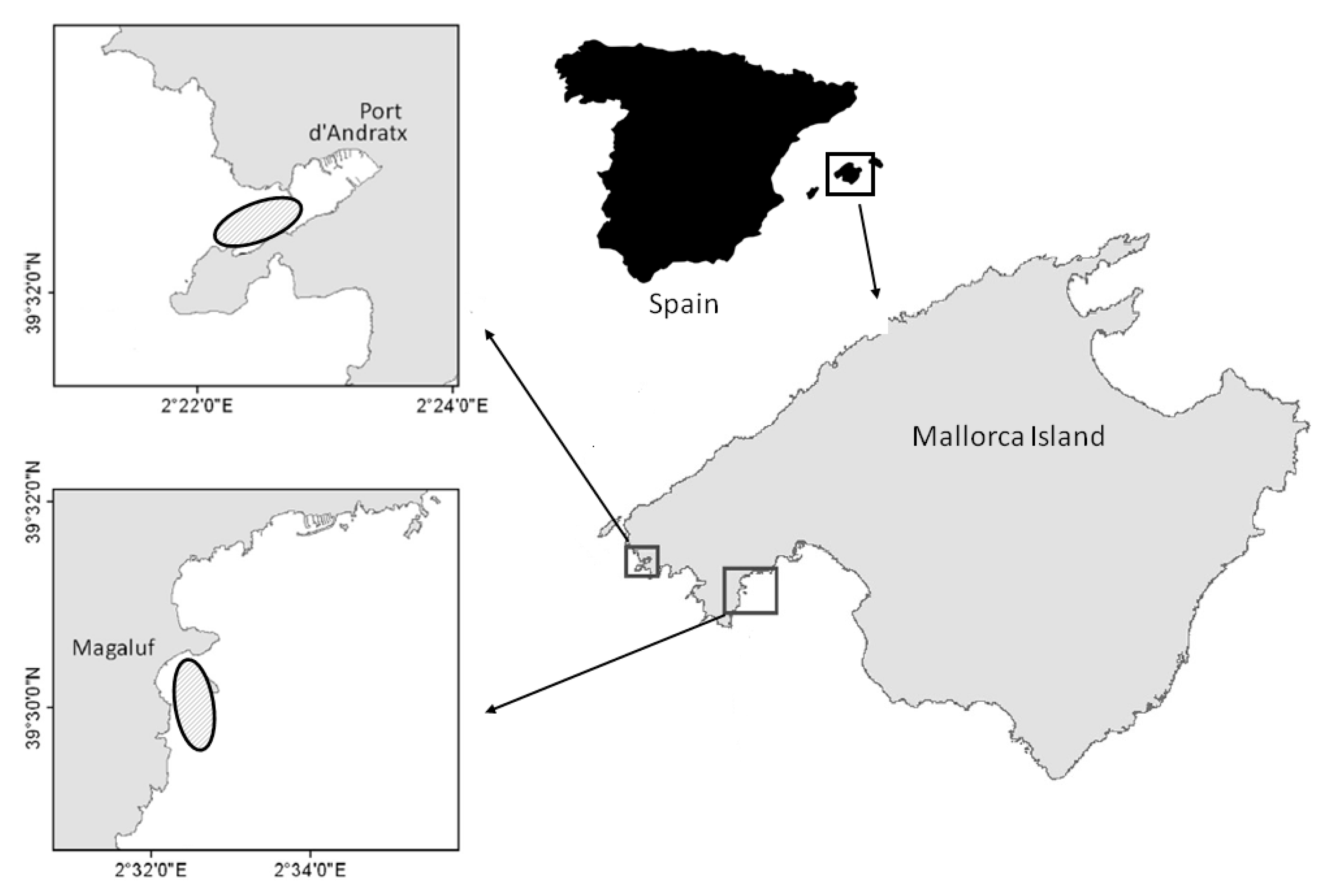
5.1. Sampling and Description of Collection Area
5.2. Histological and Molecular Analyses
5.3. Sample Homogenization for Biochemical Analyses
5.4. Antioxidant Enzyme Activities
5.5. Malondialdehyde (MDA) Assay
5.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Richardson, C.A.; Kennedy, H.; Duarte, C.M.; Kennedy, D.P.; Proud, S.V. Age and growth of the fan mussel Pinna nobilis from southeast Spanish Mediterranean seagrass (Posidonia oceanica) meadows. Mar. Biol. 1999, 133, 205–212. [Google Scholar] [CrossRef]
- García-March, J.R.; García-Carrascosa, A.M.; Cantero, A.P.; Wang, Y.G. Population structure, mortality and growth of Pinna nobilis Linnaeus, 1758 (Mollusca, Bivalvia) at different depths in Moraira bay (Alicante, Western Mediterranean). Mar. Biol. 2007, 150, 861–871. [Google Scholar] [CrossRef]
- Sanna, D.; Cossu, P.; Dedola, G.L.; Scarpa, F.; Maltagliati, F.; Castelli, A.; Franzoi, P.; Lai, T.; Cristo, B.; Curini-Galletti, M.; et al. Mitochondrial DNA reveals genetic structuring of Pinna nobilis across the Mediterranean Sea. PLoS ONE 2013, 8, e67372. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Luis, M.; Álvarez, E.; Barrajón, A.; García-March, J.R.; Grau, A.; Hendriks, I.E.; Jiménez, S.; Kersting, D.; Moreno, D.; Pérez, M.; et al. SOS Pinna nobilis: A mass mortality event in western Mediterranean Sea. Front. Mar. Sci. 2017, 4, 220. [Google Scholar] [CrossRef]
- European Commission. Interpretation Manual of European Union Habitats; EUR 27; DG Environment: Brussels, Belgium, 2007. [Google Scholar]
- Spanish government. Royal Decree 139/2011, of February 4, for the development of the List of Wild Species under the Special Protection Regime and the Spanish Catalog of Threatened Species. BOE 2011, 46, 20912–20951. [Google Scholar]
- Kersting, D.; Benabdi, M.; Čižmek, H.; Grau, A.; Jimenez, C.; Katsanevakis, S.; Öztürk, B.; Tuncer, S.; Tunesi, L.; Vázquez-Luis, M.; et al. Pinna nobilis. IUCN Red List Threat. Species 2019. [Google Scholar] [CrossRef]
- Catanese, G.; Grau, A.; Valencia, J.M.; Garcia-March, J.R.; Vázquez-Luis, M.; Alvarez, E.; Deudero, S.; Darriba, S.; Carballal, M.J.; Villalba, A. Haplosporidium pinnae sp. nov., a haplosporidan parasite associated with mass mortalities of the fan mussel, Pinna nobilis, in the Western Mediterranean Sea. J. Invertebr. Pathol. 2018, 157, 9–24. [Google Scholar] [CrossRef]
- Carella, F.; Aceto, S.; Pollaro, F.; Miccio, A.; Iaria, C.; Carrasco, N.; Prado, P.; De Vico, G. 2019. A mycobacterial disease is associated with the silent mass mortality of the pen shell Pinna nobilis along the Tyrrhenian coastline of Italy. Sci. Rep. 2019, 9, 2725. [Google Scholar] [CrossRef]
- Carella, F.; Elisabetta, A.; Simone, F.; Fulvio, S.; Daniela, M.; Prado, P.; Panarese, R.; Marino, F.; Fiocchi, E.; Pretto, T.; et al. In the Wake of the Ongoing Mass Mortality Events: Co-occurrence of Mycobacterium, Haplosporidium and Other Pathogens in Pinna nobilis Collected in Italy and Spain (Mediterranean Sea). Front. Mar. Sci. 2020, 7, 48. [Google Scholar] [CrossRef]
- Prado, P.; Carrasco, N.; Catanese, G.; Grau, A.; Cabanes, P.; Carella, F.M.; Garcia-March, J.R.; Tena, J.; Roque, A.; Bertomeu, E.; et al. Presence of Vibrio mediterranei associated to major mortality in stabled individuals of Pinna nobilis L. Aquaculture 2020, 519, 734899. [Google Scholar] [CrossRef]
- García-March, J.R.; Tena, J.; Henandis, S.; Vázquez-Luis, M.; López, D.; Téllez, C.; Prado, P.; Navas, J.I.; Bernal, J.; Catanese, G.; et al. Can we save a marine species affected by a highly infective, highly lethal, waterborne disease from extinction? Biol. Conserv. 2020, 243, 108498. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Tsirintanis, K.; Tsaparis, D.; Doukas, D.; Sini, M.; Athanassopoulou, F.; Κolygas, M.N.; Tontis, D.; Koutsoubas, D.; Bakopoulos, V. The cryptogenic parasite Haplosporidium pinnae invades the Aegean Sea and causes the collapse of Pinna nobilis populations. Aquat. Invasions 2019, 14, 150–164. [Google Scholar] [CrossRef]
- Čižmek, H.; Čolić, B.; Gračan, R.; Grau, A.; Catanese, G. An emergency situation for pen shells in the Mediterranean: The Adriatic Sea, one of the last Pinna nobilis shelters, is now affected by a mass mortality event. J. Invertebr. Pathol. 2020, 173, 107388. [Google Scholar] [CrossRef] [PubMed]
- Šarić, T.; Župan, I.; Aceto, S.; Villari, G.; Palić, D.; De Vico, G.; Carella, F. Epidemiology of noble pen shell (Pinna nobilis L. 1758) mass mortality events in Adriatic Sea is characterized with rapid spreading and acute disease progression. Pathogens 2020, 9, 776. [Google Scholar] [CrossRef] [PubMed]
- Ramilo, A.; Carrasco, N.; Reece, K.S.; Valencia, J.M.; Grau, A.; Aceituno, P.; Rojas, M.; Gairin, I.; Furones, M.D.; Abollo, E.; et al. Update of information on perkinsosis in NW Mediterranean coast: Identification of Perkinsus spp. (Protista) in new locations and hosts. J. Invertebr. Pathol. 2015, 125, 37–41. [Google Scholar] [CrossRef]
- Robledo, J.A.F.; Vasta, G.R.; Record, N.R. Protozoan parasites of bivalve molluscs: Literature follows culture. PLoS ONE 2014, 9, e100872. [Google Scholar]
- Bower, S.M.; McGladdery, S.E.; Price, I.M. Synopsis of infectious diseases and parasites of commercially exploited shellfish. Annu. Rev. Fish Dis. 1994, 4, 1–199. [Google Scholar] [CrossRef]
- Comps, M. Observations relatives à l’infection branchiale des huîtres portugaises (Crassostrea angulata Lmk). Rev. Trav. Inst. Pêches Marit. 1970, 33, 151–160. [Google Scholar]
- Burreson, E.M.; Stokes, N.A.; Carnegie, R.B.; Bishop, M.J. Bonamia sp. (Haplosporidia) found in nonnative oysters Crassostrea ariakensis in Bogue Sound, North Carolina. J. Aquat. Anim. Health 2004, 16, 1–9. [Google Scholar] [CrossRef]
- Burreson, E.M.; Ford, S.E. A review of recent information on the Haplosporidia, with special reference to Haplosporidium nelsoni (MSX disease). Aquat. Living Resour. 2004, 17, 499–517. [Google Scholar] [CrossRef]
- Waki, T.; Shimokawa, J.; Watanabe, S.; Yoshinaga, T.; Ogawa, K. Experimental challenges of wild Manila clams with Perkinsus species isolated from naturally infected wild Manila clams. J. Invertebr. Pathol. 2012, 111, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Bottari, T.; Scarfì, G.; Giacobbe, S. Mass Mortality in Noah’S Ark Arca noae (Linnaeus, 1758): A Case Study from the Strait of Messina (Mediterranean Sea). J. Shellfish Res. 2017, 36, 749–753. [Google Scholar] [CrossRef]
- Kersting, D.K.; García-March, J.R.; Templado, J. Evaluation of Spondylus gaederopus Linné, 1758, mass mortality event in the Columbretes Islands Marine Reserve (western Mediterranean, Spain). Org. Divers. Evol. 2006, 6. Available online: http://www.senckenbergmuseum.de/odes/06-16/Kersting_et_al_Spondylus-MortalityMediterran.pdf (accessed on 10 November 2020).
- Ertl, N.G.; O’Connor, W.A.; Papanicolaou, A.; Wiegand, A.N.; Elizur, A. Transcriptome analysis of the Sydney rock oyster, Saccostrea glomerata: Insights into molluscan immunity. PLoS ONE 2016, 11, e0156649. [Google Scholar] [CrossRef] [PubMed]
- Box, A.; Capó, X.; Tejada, S.; Sureda, A.; Mejías, L.; Valencia, J.M. Perkinsus mediterraneus infection induces oxidative stress in the mollusc Mimachlamys varia. J. Fish Dis. 2020, 43, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Morga, B.; Arzul, I.; Chollet, B.; Renault, T. Infection with the protozoan parasite Bonamia ostreae modifies in vitro haemocyte activities of flat oyster Ostrea edulis. Fish Shellfish Immunol. 2009, 26, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef]
- Sureda, A.; Box, A.; Tejada, S.; Blanco, A.; Caixach, J.; Deudero, S. Biochemical responses of Mytilus galloprovincialis as biomarkers of acute environmental pollution caused by the Don Pedro oil spill (Eivissa Island, Spain). Aquat. Toxicol. 2011, 101, 540–549. [Google Scholar] [CrossRef]
- Capó, X.; Tejada, S.; Box, A.; Deudero, S.; Sureda, A. Oxidative status assessment of the endemic bivalve Pinna nobilis affected by the oil spill from the sinking of the Don Pedro. Mar. Environ. Res. 2015, 110, 19–24. [Google Scholar] [CrossRef]
- Sureda, A.; Capó, X.; Busquets-Cortés, C.; Tejada, S. Acute exposure to sunscreen containing titanium induces an adaptive response and oxidative stress in Mytillus galloprovincialis. Ecotoxicol. Environ. Saf. 2018, 149, 58–63. [Google Scholar] [CrossRef]
- Box, A.; Sureda, A.; Galgani, F.; Pons, A.; Deudero, S. Assessment of environmental pollution at Balearic Islands applying oxidative stress biomarkers in the mussel Mytilus galloprovincialis. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Sillero-Ríos, J.; Sureda, A.; Capó, X.; Oliver-Codorniú, M.; Arechavala-Lopez, P. Biomarkers of physiological responses of Octopus vulgaris to different coastal environments in the western Mediterranean Sea. Mar. Pollut. Bull. 2018, 128, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Panarese, R.; Tedesco, P.; Chimienti, G.; Latrofa, M.S.; Quaglio, F.; Passantino, G.; Buonavoglia, C.; Gustinelli, A.; Tursi, A.; Otranto, D. Haplosporidium pinnae associated with mass mortality in endangered Pinna nobilis (Linnaeus 1758) fan mussels. J. Invertebr. Pathol. 2019, 164, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Cabanellas-Reboredo, M.; Vázquez-Luis, M.; Mourre, B.; Álvarez, E.; Deudero, S.; Amores, Á.; Addis, P.; Ballesteros, E.; Barrajón, A.; Coppa, S.; et al. Tracking a mass mortality outbreak of pen shell Pinna nobilis populations: A collaborative effort of scientists and citizens. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Scarpa, F.; Sanna, D.; Azzena, I.; Mugetti, D.; Cerruti, F.; Hosseini, S.; Cossu, P.; Pinna, S.; Grech, D.; Cabana, D.; et al. Multiple non-species-specific pathogens possibly triggered the mass mortality in Pinna nobilis. Life 2020, 10, 238. [Google Scholar] [CrossRef]
- Zannella, C.; Mosca, F.; Mariani, F.; Franci, G.; Folliero, V.; Galdiero, M.; Tiscar, P.G.; Galdiero, M. Microbial Diseases of Bivalve Mollusks: Infections, Immunology and Antimicrobial Defense. Mar. Drugs 2017, 15, 182. [Google Scholar] [CrossRef]
- Cheng, T.C.; Bui, M.N.; Howland, K.H.; Schoenberg, D.A.; Sullivan, J.T. Effect of preinjection of Crassostrea virginica with bacteria on subsequent chemotactic response by its hemocytes. J. Invertebr. Pathol. 1981, 38, 122–126. [Google Scholar] [CrossRef]
- Darriba, S. First haplosporidan parasite reported infecting a member of the Superfamily Pinnoidea (Pinna nobilis) during a mortality event in Alicante (Spain, Western Mediterranean). J. Invertebr. Pathol. 2017, 148, 14–19. [Google Scholar] [CrossRef]
- Hine, P.M.; Thorne, T. Haplosporidium sp. (Alveolata: Haplosporidia) associated with mortalities among rock oysters Saccostrea cuccullata in north Western Australia. Dis. Aquat. Org. 2002, 51, 123–133. [Google Scholar] [CrossRef]
- López-Sanmartín, M.; Catanese, G.; Grau, A.; Valencia, J.M.; García-March, J.R.; Navas, J.I. Correction: Real-Time PCR based test for the early diagnosis of Haplosporidium pinnae affecting fan mussel Pinna nobilis. PLoS ONE 2020, 15, e0229548. [Google Scholar] [CrossRef]
- Deudero, S.; Vázquez-Luis, M.; Álvarez, E. Human stressors are driving coastal benthic long-lived sessile fan mussel Pinna nobilis population structure more than environmental stressors. PLoS ONE 2015, 10, e0134530. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Luis, M.; Banach-Esteve, G.; Alvarez, E.; Deudero, S. Colonization on Pinna nobilis at a marine protected area: Extent of the spread of two invasive seaweeds. J. Mar. Biolog. Assoc. UK 2014, 94, 857. [Google Scholar] [CrossRef]
- Tiscar, P.G.; Mosca, F. Defense mechanisms in farmed marine mollusks. Vet. Res. Commun. 2004, 28, 57. [Google Scholar] [CrossRef]
- Matozzo, V.; Pagano, M.; Spinelli, A.; Caicci, F.; Faggio, C. Pinna nobilis: A big bivalve with big haemocytes? Fish Shellfish Immunol. 2016, 55, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Soudant, P.; Chu, F.L.E.; Volety, A. Host–parasite interactions: Marine bivalve molluscs and protozoan parasites, Perkinsus species. J. Invertebr. Pathol. 2013, 114, 196–216. [Google Scholar] [CrossRef] [PubMed]
- Morga, B.; Renault, T.; Faury, N.; Chollet, B.; Arzul, I. Cellular and molecular responses of haemocytes from Ostrea edulis during in vitro infection by the parasite Bonamia ostreae. Int. J. Parasitol. 2011, 41, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Barber, B.J.; Ford, S.E.; Haskin, H.H. Effects of the parasite MSX (Haplosporidium nelsoni) on oyster (Crassostrea virginica) energy metabolism. I. Condition index and relative fecundity. J. Shellfish Res. 1988, 7, 25–31. [Google Scholar]
- Ford, S.E. Comparison of hemolymph proteins from resistant and susceptible oysters, Crassostrea virginica, exposed to the parasite Haplosporidium nelsoni (MSX). J. Invertebr. Pathol. 1986, 47, 283–294. [Google Scholar] [CrossRef]
- Bonnes-Taourel, D.; Guérin, M.C.; Torreilles, J. Is malonaldehyde a valuable indicator of lipid peroxidation? Biochem. Pharmacol. 1992, 44, 985–988. [Google Scholar] [CrossRef]
- Gutteridge, J.M. Lipid peroxidation and antioxidants as biomarkers of tissue damage. Clin. Chem. 1995, 41, 1819–1828. [Google Scholar] [CrossRef]
- Moore, M.N.; Shaw, J.P.; Pascoe, C.; Beesley, A.; Viarengo, A.; Lowe, D.M. Anti-oxidative hormetic effects of cellular autophagy induced by nutrient deprivation in a molluscan animal model. Mar. Environ. Res. 2020, 156, 104903. [Google Scholar] [CrossRef] [PubMed]
- Genard, B.; Pernet, F.; Lemarchand, K.; Boudry, P.; Moraga, D.; Tremblay, R. Physiological and biochemical changes associated with massive mortality events occurring in larvae of American oyster (Crassostrea virginica). Aquat. Living Resour. 2011, 24, 247–260. [Google Scholar] [CrossRef]
- Lassudrie, M.; Soudant, P.; Richard, G.; Henry, N.; Medhioub, W.; da Silva, P.M.; Donval, A.; Bunel, M.; Le Goïc, N.; Lambert, C.; et al. Physiological responses of Manila clams Venerupis (=Ruditapes) philippinarum with varying parasite Perkinsus olseni burden to toxic algal Alexandrium ostenfeldii exposure. Aquat. Toxicol. 2014, 154, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Neves, C.A.; Santos, E.A.; Bainy, A.C.D. Reduced superoxide dismutase activity in Palaemonetes argentinus (Decapoda, Palemonidae) infected by Probopyrus ringueleti (Isopoda, Bopyridae). Dis. Aquat. Org. 2000, 39, 155–158. [Google Scholar] [CrossRef]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Howard, D.W. Histological Techniques for Marine Bivalve Mollusks and Crustaceans; NOAA, National Ocean Service, National Centers for Coastal Ocean Service, Center for Coastal Environmental Health and Biomolecular Research: Charleston, SC, USA; Cooperative Oxford Laboratory: Annapolis MD, USA, 2004; Volume 5. [Google Scholar]
- Böddinghaus, B.; Rogall, T.; Flohr, T.; Blöcker, H.; Böttger, E.C. Detection and identification of mycobacteria by amplification of rRNA. J. Clin. Microbiol. 1990, 28, 1751–1759. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Sympos. Ser. 1999, 41, 95–98. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Flohé, L.; Gunzler, W.A. Assays of Glutathione Peroxidase. Methods Enzymol. 1984, 105, 114–120. [Google Scholar]
- Goldberg, D.M.; Spooner, R.J. Assay of Glutathione Reductase. In Methods of Enzymatic Analysis, 3rd ed.; Bergmeyen, H.V., Ed.; Verlag Chemie: Deerfiled Beach, FL, USA, 1983; Volume 3, pp. 258–265. [Google Scholar]
- Flohé, L.; Ötting, F. Superoxide Dismutase Assay. Methods Enzymol. 1984, 105, 93–104. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | H. pinnae | Mycobacterium sp. | Other Gram-Negative Bacteria | Date | H. pinnae | Mycobacterium sp. | Other Gram-Negative Bacteria |
|---|---|---|---|---|---|---|---|
| 03/06/2011 | − | + | + | 24/11/2016 | + | − | − |
| 03/06/2011 | − | − | + | 24/11/2016 | + | + | + |
| 03/06/2011 | − | − | − | 24/11/2016 | + | + | + |
| 21/06/2011 | − | + | + | 03/03/2017 | + | + | − |
| 21/06/2011 | − | + | − | 03/03/2017 | + | − | + |
| 21/09/2011 | − | + | − | 03/03/2017 | + | + | + |
| 11/10/2011 | − | − | − | 03/03/2017 | + | + | + |
| 25/11/2011 | − | − | − | 03/03/2017 | + | − | + |
| 10/02/2012 | − | + | − | 26/06/2017 | + | − | − |
| 15/03/2012 | − | + | + | 26/06/2017 | + | − | − |
| n | Catalase | SOD | GPx | GRd | MDA | |
|---|---|---|---|---|---|---|
| H. pinnae (−) Mycobacterium sp. (−) | 4 | 115 ± 7 | 4.16 ± 0.43 | 3.24 ± 0.63 | 3.58 ± 1.25 | 7.10 ± 0.61 |
| H. pinnae (−) Mycobacterium sp. (+) | 6 | 138 ± 15 | 3.86 ± 0.10 | 2.98 ± 0.27 | 3.04 ± 0.72 | 7.13 ± 0.27 |
| H. pinnae (+) Mycobacterium sp. (−) | 5 | 62.0 ± 3.2 * | 2.06 ± 0.29 * | 2.68 ± 0.19 | 2.22 ± 0.19 | 28.3 ± 3.1 * |
| H. pinnae (+) Mycobacterium sp. (+) | 5 | 61.8 ± 6.8 * | 2.52 ± 0.32 * | 2.73 ± 0.19 | 2.60 ± 0.48 | 28.2 ± 2.2 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Box, A.; Capó, X.; Tejada, S.; Catanese, G.; Grau, A.; Deudero, S.; Sureda, A.; Valencia, J.M. Reduced Antioxidant Response of the Fan Mussel Pinna nobilis Related to the Presence of Haplosporidium pinnae. Pathogens 2020, 9, 932. https://doi.org/10.3390/pathogens9110932
Box A, Capó X, Tejada S, Catanese G, Grau A, Deudero S, Sureda A, Valencia JM. Reduced Antioxidant Response of the Fan Mussel Pinna nobilis Related to the Presence of Haplosporidium pinnae. Pathogens. 2020; 9(11):932. https://doi.org/10.3390/pathogens9110932
Chicago/Turabian StyleBox, Antonio, Xavier Capó, Silvia Tejada, Gaetano Catanese, Amalia Grau, Salud Deudero, Antoni Sureda, and José María Valencia. 2020. "Reduced Antioxidant Response of the Fan Mussel Pinna nobilis Related to the Presence of Haplosporidium pinnae" Pathogens 9, no. 11: 932. https://doi.org/10.3390/pathogens9110932
APA StyleBox, A., Capó, X., Tejada, S., Catanese, G., Grau, A., Deudero, S., Sureda, A., & Valencia, J. M. (2020). Reduced Antioxidant Response of the Fan Mussel Pinna nobilis Related to the Presence of Haplosporidium pinnae. Pathogens, 9(11), 932. https://doi.org/10.3390/pathogens9110932