The Notorious Soilborne Pathogenic Fungus Sclerotinia sclerotiorum: An Update on Genes Studied with Mutant Analysis



Abstract
1. Introduction
2. The Features of the S. sclerotiorum Genome
2.1. Genome Sequences
2.2. Transcriptomic and Secretomic Analysis
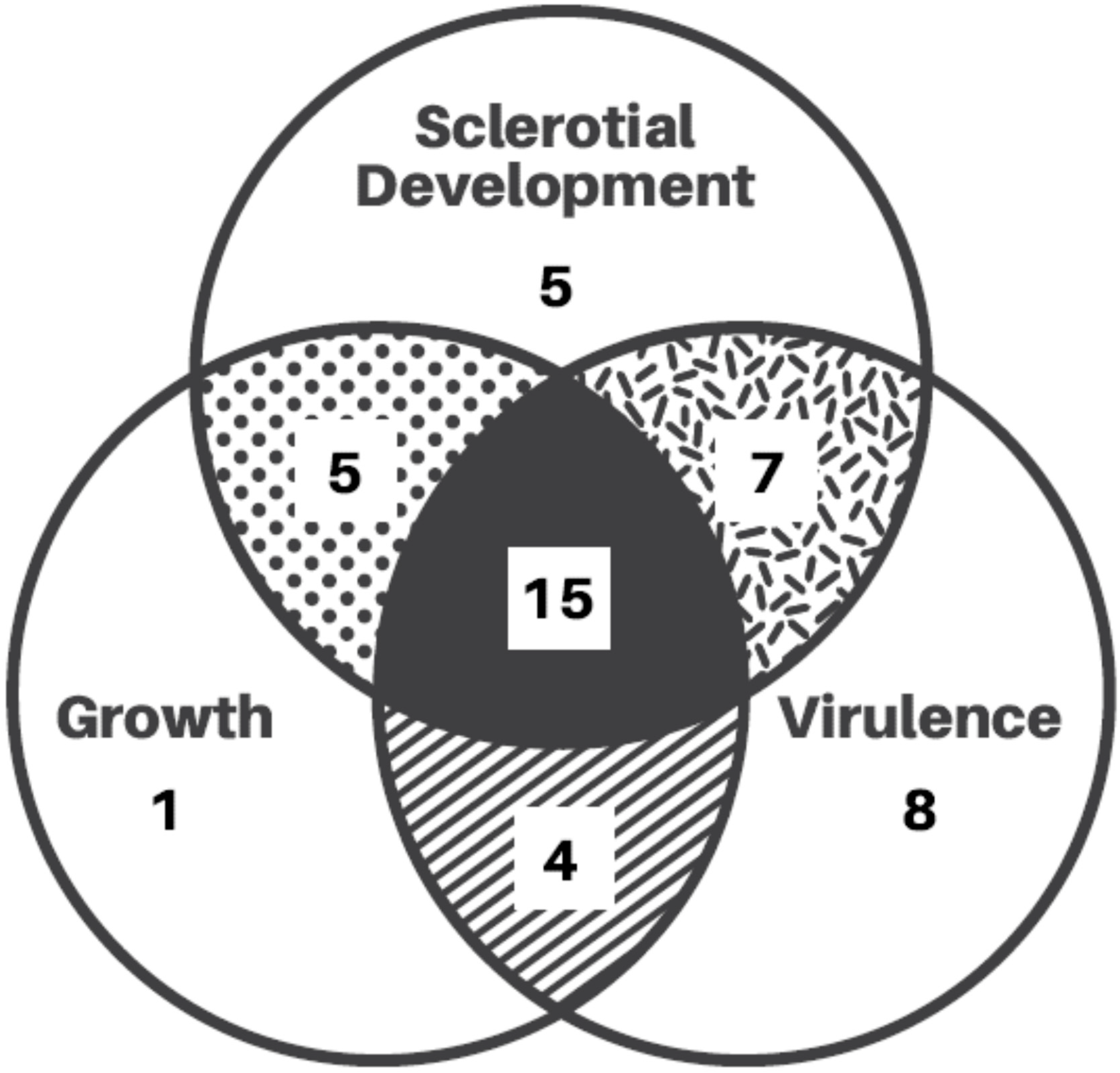
3. Molecular Dissection of S. sclerotiorum Biology
3.1. Regulation of Mycelial Growth and Virulence
3.2. Signaling Events Leading to Sclerotial Formation
3.3. Control of Apothecial and Ascospore Development
3.4. Regulation of Fungal Pathogenesis of the Fungus
4. Summary and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Libert, M.A. Plante Crytogamicae Arduennae (Exsiccati); 1837; No. 326; Published by the Author. [Google Scholar]
- Whetzel, H.H. A synopsis of the genera and species of the Sclerotiniaceae, a family of stromatic inoperculate discomycetes. Mycologia 1945, 37, 648–714. [Google Scholar] [CrossRef]
- Buchwald, N.F.; Neergaard, P.J. A plea for the retention of Sclerotinia sclerotiorum as type species for the genus Sclerotinia Fckl. emend. Friesia 1972, 10, 96–99. [Google Scholar]
- Buchwald, N.F.; Neergaard, P.J. Proposal to conserve Sclerotinia Fckl, with S. Sclerotiorum (Lib.) de bary as type species. Taxon 1976, 25, 199–200. [Google Scholar] [CrossRef]
- Kohn, L.M. Delimitation of the economically important plant pathogenic Sclerotinia species. Phytopathology 1979, 69, 881–886. [Google Scholar] [CrossRef]
- Kabbage, M.; Yarden, O.; Dickman, M.B. Pathogenic attributes of Sclerotinia sclerotiorum: Switching from a biotrophic to necrotrophic Lifestyle. Plant Sci. 2015, 233, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Bao, L.L.; Zhao, F.Y.; Tang, M.Q.; Chen, T.; Li, Y.; Wang, B.X.; Fu, B.; Fang, H.; Li, G.Y.; et al. BnaMPK3 is a key regulator of defense responses to the devastating plant pathogen Sclerotinia sclerotiorum in oilseed rape. Front. Plant Sci. 2019, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Willetts, H.; Wong, J.J. The biology of Sclerotinia sclerotiorum, S. Trifoliorum, and S. Minor with emphasis on specific nomenclature. Bot. Rev. 1980, 46, 101–165. [Google Scholar] [CrossRef]
- Harper, G.E.; Frampton, C.M.; Stewart, A. Factors influencing survival of sclerotia of Sclerotium cepivorum in New Zealand soils. N. Z. J. Crop Hortic. Sci. 2002, 30, 29–35. [Google Scholar] [CrossRef]
- Boland, G.J.; Hall, R. Index of plant hosts of Sclerotinia sclerotiorum. Plant Sci. 1994, 16, 93–108. [Google Scholar] [CrossRef]
- Liang, X.; Rollins, J.A. Mechanisms of broad host range necrotrophic pathogenesis in Sclerotinia sclerotiorum. Phytopathology 2018, 108, 1128–1140. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P.H.J.; Nelson, B.D. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Li, J.; Yu, B.; Liu, L.; Zhang, X.; Liu, J.; Montes, H.M. Transcription factor SsSte12 was involved in mycelium growth and development in Sclerotinia sclerotiorum. Front. Microbiol. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Qian, L.; Disi, J.O.; Yang, X.; Li, Q.; Li, J.; Frauen, M.; Cai, D.; Qian, W. Identification of resistant sources against Sclerotinia sclerotiorum in brassica species with emphasis on B. Oleracea. Euphytica 2011, 177, 393–399. [Google Scholar] [CrossRef]
- Li, G.Q.; Huang, H.C.; Miao, H.J.; Erickson, R.S.; Jiang, D.H.; Xiao, Y.N. Biological control of Sclerotinia diseases of rapeseed by aerial applications of the mycoparasite Coniothyrium minitans. Eur. J. Plant Pathol. 2006, 114, 345–355. [Google Scholar] [CrossRef]
- Yang, S.J. An investigation on the host range and some ecological aspects on the Sclerotinia disease of the Rape plant. Acta Phytopathol. Sin. 1959, 5, 111–122. [Google Scholar]
- Moore, W.J. Flooding as a means of destroying the sclerotia of Sclerotinia sclerotiorum. Phytopathology 1949, 39, 920–927. [Google Scholar]
- Schwartz, H.; Steadman, J.J. Factors affecting sclerotium populations of, and apothecium production by, Sclerotinia sclerotiorum. Phytopathology 1978, 68, 383–388. [Google Scholar] [CrossRef]
- Stoner, W.N.; Moore, W.D. Lowland rice farming, a possible cultural control for Sclerotinia sclerotiorum in the Everglades. Plant Dis. Rep. 1953, 37, 181–186. [Google Scholar]
- Coyne, D.; Steadman, J.; Anderson, F.J. Effect of modified plant architecture of great northern dry bean varieties (Phaseolus vulgaris) on white mold severity, and components of yield. Plant Dis. Rep. 1974, 58, 379–382. [Google Scholar]
- Amselem, J.; Cuomo, C.A.; Van Kan, J.A.; Viaud, M.; Benito, E.P.; Couloux, A.; Coutinho, P.M.; De Vries, R.P.; Dyer, P.S.; Fillinger, S.; et al. Genomic analysis of the necrotrophic fungal pathogens Sclerotinia sclerotiorum and Botrytis cinerea. PLoS Genet. 2011, 7, e1002230. [Google Scholar] [CrossRef]
- Charles, M.; Id, D.; Denton-giles, M.; Id, J.K.H.; Chang, S. A whole genome scan of SNP data suggests a lack of abundant hard selective sweeps in the genome of the broad host range plant pathogenic fungus Sclerotinia sclerotiorum. PLoS ONE 2019, 14, e0214201. [Google Scholar]
- Derbyshire, M.; Denton-giles, M.; Hegedus, D.; Seifbarghi, S.; Rollins, J.; Van, J.; Seidl, M.F.; Faino, L.; Mbengue, M.; Navaud, O.; et al. The complete genome sequence of the phytopathogenic fungus Sclerotinia sclerotiorum reveals insights into the genome architecture of broad host range pathogens. Genome Biol. Evol. 2017, 9, 593–618. [Google Scholar] [CrossRef] [PubMed]
- Hermisson, J.; Pennings, P.S. Soft sweeps: Molecular population genetics of adaptation from standing genetic variation. Genetics 2005, 169, 2335–2352. [Google Scholar] [CrossRef] [PubMed]
- Messer, P.W.; Petrov, D.A. Population genomics of rapid adaptation by soft selective sweeps. Trends Ecol. Evol. 2013, 28, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Del Ponte, E.M.; Eduardo, S.; Lehner, M.S.; Ju, T.J.D.P.; Mizubuti, G.; Pethybridge, S.J. Independently founded populations of Sclerotinia sclerotiorum from a tropical and a temperate region have similar genetic structure. PLoS ONE 2017, 12, e0173915. [Google Scholar]
- Seifbarghi, S.; Borhan, M.H.; Wei, Y.; Coutu, C.; Robinson, S.J.; Hegedus, D.D. Changes in the Sclerotinia sclerotiorum transcriptome during infection of Brassica napus. BMC Genom. 2017, 18, 266. [Google Scholar] [CrossRef]
- Heard, S.; Brown, N.A.; Hammond-kosack, K. An interspecies comparative analysis of the predicted secretomes of the necrotrophic plant pathogens Sclerotinia sclerotiorum and Botrytis cinerea. PLoS ONE 2015, 10, e0130534. [Google Scholar] [CrossRef]
- Doughan, B.; Rollins, J.A.; Avery, S.V. Characterization of MAT gene functions in the life cycle of Sclerotinia sclerotiorum reveals a lineage-specific MAT gene functioning in apothecium morphogenesis. Fungal Biol. 2016, 120, 1105–1117. [Google Scholar] [CrossRef]
- Lyu, X.; Shen, C.; Fu, Y.; Xie, J.; Jiang, D.; Li, G.; Cheng, J. A small secreted virulence-related protein is essential for the necrotrophic interactions of Sclerotinia sclerotiorum with its host plants. PLoS Pathog. 2016, 12, e1005435. [Google Scholar] [CrossRef]
- Yu, Y.; Du, J.; Wang, Y.; Zhang, M.; Huang, Z.; Cai, J.; Fang, A.; Yang, Y.; Qing, L.; Bi, C.; et al. Survival factor 1 contributes to the oxidative stress response and is required for full virulence of Sclerotinia sclerotiorum. Mol. Plant Pathol. 2019, 20, 895–906. [Google Scholar] [CrossRef]
- Harel, A.; Bercovich, S.; Yarden, O. Calcineurin is required for sclerotial development and pathogenicity of Sclerotinia sclerotiorum in an oxalic acid–independent manner. Mol. Plant Microbe Interact. 2006, 19, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Lyu, X.; Shen, C.; Fu, Y.; Xie, J.; Jiang, D.; Li, G. The microbial opsin homolog sop1 is involved in Sclerotinia sclerotiorum development and environmental stress response. Front. Microbiol. 2016, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, Q.; Sun, Y.; Zhang, Y.; Zhang, X.; Liu, J. Sssfh1, a gene encoding a putative component of the RSC chromatin remodeling complex, is involved in hyphal growth, reactive oxygen species accumulation, and pathogenicity in Sclerotinia sclerotiorum. Front. Microbiol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Ge, C.; Liu, S.; Wang, J.; Zhou, M. A two-component histidine kinase shk1 controls stress response, sclerotial formation and fungicide resistance in Sclerotinia sclerotiorum. Mol. Plant Pathol. 2013, 14, 708–718. [Google Scholar] [CrossRef]
- Xu, L.; Chen, W. Random T-DNA mutagenesis identifies a Cu/Zn superoxide dismutase gene as a virulence factor of Sclerotinia sclerotiorum. Mol. Plant Microbe Interact. 2013, 26, 431–441. [Google Scholar] [CrossRef]
- Veluchamy, S.; Williams, B.; Kim, K.; Dickman, M.B. Physiological and molecular plant pathology the CuZn superoxide dismutase from Sclerotinia sclerotiorum is involved with oxidative stress tolerance, virulence, and oxalate production. Physiol. Mol. Plant Pathol. 2012, 78, 14–23. [Google Scholar] [CrossRef]
- Liang, Y.; Yajima, W.; Davis, M.R.; Kav, N.N.V.; Stephen, E. Disruption of a gene encoding a hypothetical secreted protein from Sclerotinia sclerotiorum reduces its virulence on canola (Brassica Napus) disruption of a gene encoding a hypothetical secreted protein from Sclerotinia sclerotiorum reduces its virulence on Canola (Brassica napus). Can. J. Plant Pathol. 2013, 35, 46–55. [Google Scholar]
- Liang, Y.U.E.; Xiong, W.E.I.; Steinkellner, S.; Feng, J.I.E. Deficiency of the melanin biosynthesis genes SCD1 and THR1 affects sclerotial development and vegetative growth, but not pathogenicity, in Sclerotinia sclerotiorum. Mol. Plant Pathol. 2017, 19, 1–10. [Google Scholar] [CrossRef]
- Li, J.T.; Zhang, Y.H.; Zhang, Y.C.; Yu, P.L.; Pan, H.Y.; Rollins, J.A. Introduction of large sequence inserts by CRISPR-Cas9 to create pathogenicity mutants in the multinucleate filamentous pathogen Sclerotinia sclerotiorum. MBio 2018, 9, e00567-18. [Google Scholar] [CrossRef]
- Liberti, D.; Rollins, J.A.; Dobinson, K.F. Peroxysomal carnitine acetyl transferase influences host colonization capacity in Sclerotinia sclerotiorum. Mol. Plant Microbe Interact. 2013, 26, 768–780. [Google Scholar] [CrossRef]
- Yajima, W.; Liang, Y.; Kav, N.N.V.; Sclerotinia, A.; Bary, D. Gene disruption of an arabinofuranosidase/β-xylosidase precursor decreases Sclerotinia sclerotiorum virulence on canola tissue. Mol. Plant Microbe Interact. 2009, 22, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Xie, J.; Cheng, J.; Li, G.; Yi, X.; Jiang, D.; Fu, Y. Novel secretory protein Ss-Caf1 of the plant-pathogenic fungus Sclerotinia sclerotiorum is required for host penetration and normal sclerotial development. Mol. Plant Microbe Interact. 2014, 27, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Yarden, O.; Veluchamy, S.; Dickman, M.B.; Kabbage, M. Physiological and molecular plant pathology Sclerotinia sclerotiorum catalase SCAT1 affects oxidative stress tolerance, regulates ergosterol levels and controls pathogenic development. Physiol. Mol. Plant Pathol. 2014, 85, 34–41. [Google Scholar] [CrossRef]
- Lyu, X.; Shen, C.; Fu, Y.; Xie, J.; Jiang, D.; Li, G.; Cheng, J. Comparative genomic and transcriptional analyses of the carbohydrate-active enzymes and secretomes of phytopathogenic fungi reveal their significant roles during infection and development. Sci. Rep. 2015, 5, 15565. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Jiang, D.; Xie, J.; Cheng, J.; Li, G.; Yi, X.; Fu, Y. Ss-Sl2, a novel cell wall protein with PAN modules, is essential for sclerotial development and cellular integrity of Sclerotinia sclerotiorum. PLoS ONE 2012, 7, e34962. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.; Du, J.; Huang, Z.; Fang, A.; Yang, Y. Sclerotinia sclerotiorum thioredoxin reductase is required for oxidative stress tolerance, virulence, and sclerotial development. Front. Microbiol. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Yu, Y.; Xiao, J.; Yang, Y.; Bi, C.; Qing, L.; Tan, W. Physiological and molecular plant pathology Ss-Bi1 encodes a putative BAX Inhibitor-1 protein that is required for full virulence of Sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2015, 90, 115–122. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y.; Liu, J.; Zhang, Y.; Zhang, X.; Pan, H. The Sclerotinia sclerotiorum FoxE2 gene is required for apothecial development. Mycology 2016, 106, 484–490. [Google Scholar] [CrossRef]
- Kim, H.; Chen, C.; Kabbage, M.; Dickman, M.B. Identification and characterization of Sclerotinia sclerotiorum NADPH oxidases. Appl. Environ. Microbiol. 2011, 77, 7721–7729. [Google Scholar] [CrossRef]
- Yu, Y.; Xiao, J.; Zhu, W.; Yang, Y.; Mei, J.; Bi, C.; Qian, W.E.I.; Qing, L.; Tan, W. Ss-Rhs1, a secretory Rhs repeat-containing protein, is required for the virulence of Sclerotinia sclerotiorum. Mol. Plant Pathol. 2017, 18, 1052–1061. [Google Scholar] [CrossRef]
- Fan, H.; Yu, G.; Liu, Y.; Zhang, X.; Liu, J.; Zhang, Y.; Rollins, J.A.; Sun, F.; Pan, H. An atypical forkhead-containing transcription factor SsFKH1 is involved in sclerotial formation and is essential for pathogenicity in Sclerotinia sclerotiorum. Mol. Plant Pathol. 2017, 18, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Rollins, J.A.; Pathology, P.; Hall, F. The Sclerotinia sclerotiorum pac1 gene is required for sclerotial development and virulence. Mol. Plant Microbe Interact. 2003, 16, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Xu, Y.; Li, X.; Yao, C.; Gao, Z. Physiological and molecular plant pathology SsPemG1 encodes an elicitor-homologous protein and regulates pathogenicity in Sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2015, 92, 70–78. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Q.; Zhang, X.; Liu, J.; Zhang, Y. Ssams2, a gene encoding GATA transcription factor, is required for appressoria formation and chromosome segregation in Sclerotinia sclerotiorum. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jurick, W.M.; Dickman, M.B.; Rollins, J.A. Characterization and functional analysis of a CAMP-dependent protein kinase a catalytic subunit gene (Pka1) in Sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2004, 64, 155–163. [Google Scholar] [CrossRef]
- Chen, J.; Ullah, C.; Reichelt, M.; Gershenzon, J.; Hammerbacher, A. Sclerotinia sclerotiorum circumvents flavonoid defenses by catabolizing flavonol glycosides and aglycones. Plant Physiol. 2019, 180, 1975–1987. [Google Scholar] [CrossRef]
- Veluchamy, S.; Rollins, J.A. A CRY-DASH-type photolyase/cryptochrome from Sclerotinia sclerotiorum mediates minor UV-A-specific effects on development. Fungal Genet. Biol. 2008, 45, 1265–1276. [Google Scholar] [CrossRef]
- Li, X.L.; Guo, M.; Xu, D.F.; Chen, F.X.; Zhang, H.J.; Pan, Y.; Li, M.M.; Gao, Z. The nascent-polypeptide-associated complex alpha subunit regulates the polygalacturonases expression negatively and influences the pathogenicity of Sclerotinia sclerotiorum. Mycologia 2015, 107, 1130–1137. [Google Scholar] [CrossRef]
- Bashi, Z.D.; Gyawali, S.; Bekkaoui, D.; Coutu, C.; Lee, L.; Poon, J.; Rimmer, S.R.; Khachatourians, G.G.; Hegedus, D.D. The Sclerotinia sclerotiorum Slt2 mitogen-activated protein kinase ortholog, SMK3, is required for infection initiation but not lesion expansion. Can. J. Microbiol. 2016, 850, 836–850. [Google Scholar] [CrossRef]
- Qu, X.; Yu, B.; Liu, J.; Zhang, X.; Li, G.; Zhang, D. MADS-Box transcription factor SsMADS is involved in regulating growth and virulence in Sclerotinia sclerotiorum. Int. J. Mol. Sci. 2014, 15, 8049–8062. [Google Scholar] [CrossRef]
- Zhu, W.; Wei, W.; Fu, Y.; Cheng, J.; Xie, J.; Li, G.; Yi, X. A secretory protein of necrotrophic fungus Sclerotinia sclerotiorum that suppresses host resistance. PLoS ONE 2013, 8, e53901. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liang, X.; Rollins, J.A. Sclerotinia sclerotiorum-glutamyl transpeptidase (Ss-Ggt1) is required for regulating glutathione accumulation and development of sclerotia and compound appressoria. Mol. Plant Microbe Interact. 2012, 25, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Moomaw, E.W.; Rollins, J.A. Fungal oxalate decarboxylase activity contributes to Sclerotinia sclerotiorum early infection by affecting both compound appressoria development and function. Mol. Plant Pathol. 2015, 16, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Xiang, M.; White, D.; Chen, W. pH dependency of sclerotial development and pathogenicity revealed by using genetically defined oxalate-minus mutants of Sclerotinia sclerotiorum. Environ. Microbiol. 2015, 17, 2896–2909. [Google Scholar] [CrossRef]
- Liang, X.; Liberti, D.; Li, M.; Kim, Y.; Hutchens, A.; Wilson, R.O.N.; Rollins, J.A. Oxaloacetate acetylhydrolase gene mutants of Sclerotinia sclerotiorum do not accumulate oxalic acid, but do produce limited lesions on host plants. Mol. Plant Pathol. 2015, 16, 559–571. [Google Scholar] [CrossRef]
- Erental, A.; Harel, A.; Yarden, O. Type 2A phosphoprotein phosphatase is required for asexual development and pathogenesis of Sclerotinia sclerotiorum. Mol. Plant Microbe Interact. 2007, 20, 944–954. [Google Scholar] [CrossRef]
- Li, M.; Rollins, J.A. The development-specific Ssp1 and Ssp2 genes of Sclerotinia sclerotiorum encode lectins with distinct yet compensatory regulation. Fungal Genet. Biol. 2010, 47, 531–538. [Google Scholar] [CrossRef]
- Yu, Y.; Xiao, J.; Du, J.; Yang, Y.; Bi, C.; Qing, L. Disruption of the gene encoding endo-β-1, 4-xylanase affects the growth and virulence of Sclerotinia sclerotiorum. Front. Microbiol. 2016, 7, 1–9. [Google Scholar] [CrossRef]
- Ii, W.M.J.; Rollins, J.V.A. Deletion of the adenylate cyclase (Sac1) geneaffects multiple developmental pathways and pathogenicity in Sclerotinia sclerotiorum. Fungal Genet. Biol. 2007, 44, 521–530. [Google Scholar]
- Chen, C.; Harel, A.; Gorovoits, R.; Yarden, O.; Dickman, M.B. MAPK regulation of sclerotial development in Sclerotinia sclerotiorum is linked with pH and CAMP sensing. Mol. Plant Microbe Interact. 2004, 17, 404–413. [Google Scholar] [CrossRef]
- Yang, G.; Tang, L.; Gong, Y.; Xie, J.; Fu, Y.; Jiang, D.; Li, G.; Collinge, D.B.; Chen, W.; Cheng, J. A cerato-platanin protein SsCP1 targets plant PR1 and contributes to virulence of Sclerotinia sclerotiorum. New Phytol. 2018, 217, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Wei, J.; Yao, C.; Reng, H.; Gao, Z. Plant science SsSm1, a cerato-platanin family protein, is involved in the hyphal development and pathogenic process of Sclerotinia sclerotiorum. Plant Sci. 2018, 270, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.L.; Yu, G.; Zhang, Y.H.; Rollins, J.A.; Li, J.; Pan, H. The formaldehyde dehydrogenase SsFdh1 is regulated by and functionally cooperates with the GATA transcription. MSystems 2019, 4, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Rollins, J.A. The GATA-Type IVb zinc-finger transcription factor SsNsd1 regulates asexual—Sexual development and appressoria formation in Sclerotinia sclerotiorum. Mol. Plant Pathol. 2017, 9, 1679–1689. [Google Scholar]
- Godoy, G.; Steadman, J.R.; Dickman, M.B.; Dam, R. Use of mutants to demonstrate the role of oxalic acid in pathogenicity of Sclerotinia sclerotiorum on phaseolus vulgaris. Physiol. Mol. Plant Pathol. 1990, 37, 179–191. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
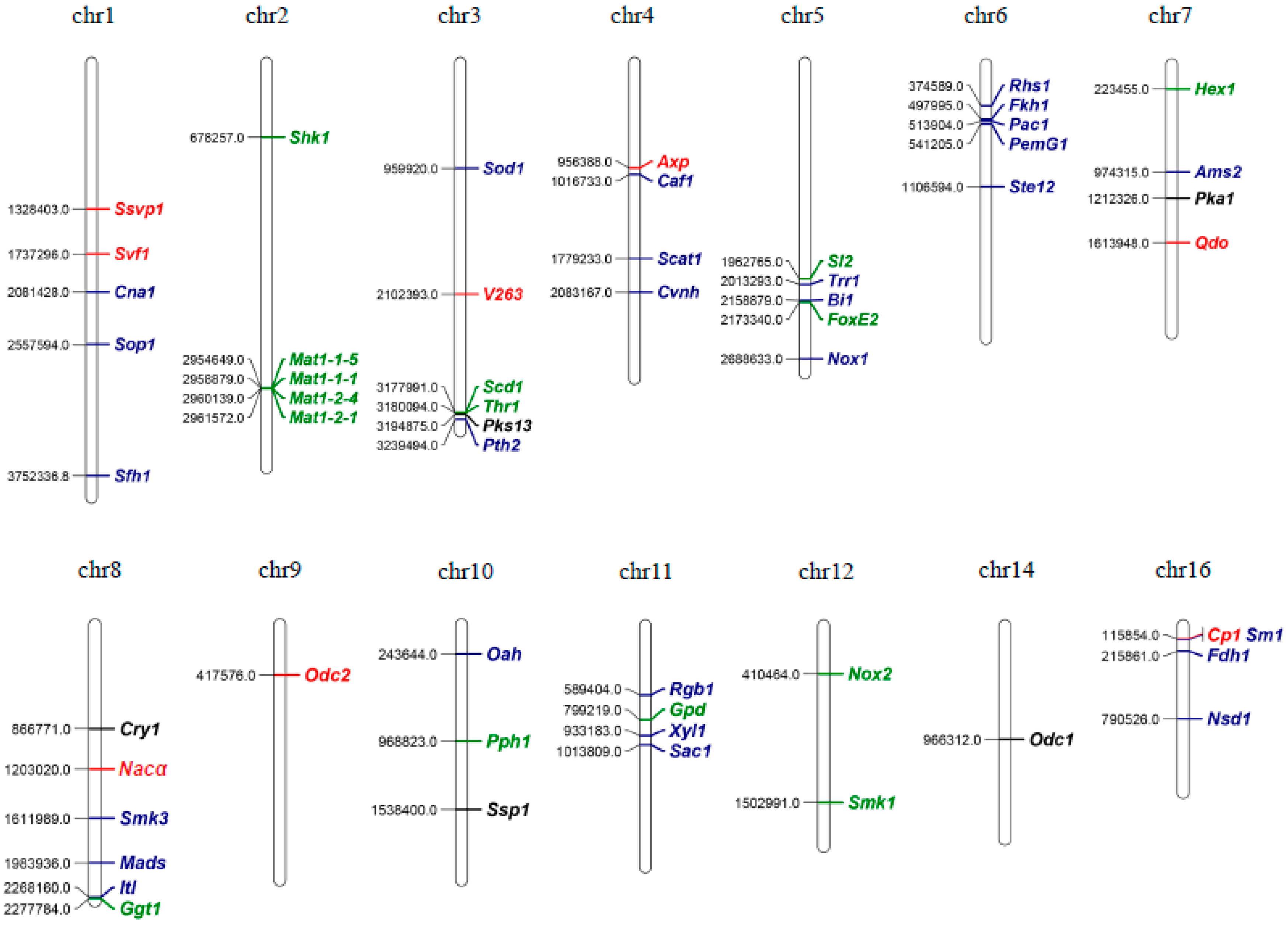
| Gene Code (New) | Gene Code (Old) | Strains | Mutant Name | Gene Full Name | Mutant Type | Mutant Phenotypes | Secretion Signal | Tested Hosts | Other Functions | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hyphal Growth | Sclerotial Formation | Oxalate Production | Virulence | Appressoria Formation | Induce Host HR/Resistance | ||||||||||
| sscle_01g003850 | SS1G_02068 | Ep-1PNA367, Heilongjiang, China | ssvp1 | Small secreted virulence-related protein | RNAi | - | NA | NA | + | NA | + | Yes | Canola (Zhongyou 821) | HR induction | [30] |
| sscle_01g004990 | SS1G_01919 | 1980, Nebraska, USA | svf1 | Survival factor 1 homologue | RNAi | - | NA | - | + | + | NA | No | Arabidopsis and canola (Zhongyou 821) | Cell wall integrity, ROS production | [31] |
| sscle_01g006030 | SS1G_01788 | 1980, Nebraska, USA | cna1 | Catalytic subunit calcineurin-encoding gene | RNAi | + | + | - | + | - | NA | No | Arabiposis and tomato (Bonny Best) | Hyphal elongation | [32] |
| sscle_01g007450 | SS1G_01614 | Ep-1PNA367, Heilongjiang, China | sop1 | Microbial opsin homolog gene | RNAi | + | + | NA | + | NA | NA | No | Arabidopsis | Stress responses | [33] |
| sscle_01g011030 | SS1G_01151 | 1980, Nebraska, USA | sfh1 | GATA-box and SNF5 domains containing transcription factor | RNAi | + | + | NA | + | + | NA | No | Soybean, common bean and tomato | ROS accumulation | [34] |
| sscle_02g013550 | SS1G_12694 | HA61, Jiangsu, China | shk1 | Histidine kinases | Deletion | + | + | NA | - | NA | NA | No | Rapeseed, strawberry, tomato and cucumber | Oxidative stresses, glycerol accumulation | [35] |
| sscle_02g020240 | SS1G_04003 | 1980, Nebraska, USA | mat1-1-5 | Mating-type gene | Deletion | - | - | NA | - | NA | NA | No | Tomato | Apothecial development | [29] |
| sscle_02g020250 | SS1G_04004 | 1980, Nebraska, USA | mat1-1-1 | Mating-type gene | Deletion | - | - | NA | - | NA | NA | No | Tomato | Apothecial development | [29] |
| sscle_02g020260 | SS1G_04005 | 1980, Nebraska, USA | mat1-2-4 | Mating-type gene | Deletion | - | - | NA | - | NA | NA | No | Tomato | Apothecial development, ascospore production | [29] |
| sscle_02g020270 | SS1G_04006 | 1980, Nebraska, USA | mat1-2-1 | Mating-type gene | Deletion | + | - | NA | - | NA | NA | No | Tomato | Apothecial development | [29] |
| sscle_03g025030 | SS1G_00699 | WMA1, Washington, USA | sod1 | Cu/Zn superoxide dismutase | T-DNA | - | - | - | + | NA | NA | No | Pea (Guido) | Detoxification of host ROS | [36] |
| sscle_03g025030 | SS1G_00699 | 1980, Nebraska, USA | sod1 | Cu/Zn superoxide dismutase | Deletion | + | + | + | + | NA | NA | No | Tomato (Garden Peach) and tobacco | Oxidative stress tolerance, repression of host ROS | [37] |
| sscle_03g028510 | SS1G_00263 | Isolated from an infected canola stem, Alberta, Canada | v263 | Hypothetical secreted protein | Deletion | - | NA | NA | + | NA | NA | Yes | Canola | [38] | |
| sscle_03g031470 | SS1G_13314 | Isolated from diseased rapeseed, Alberta, Canada | scd1 | Scytalone dehydratase | Deletion | + | + | NA | - | NA | NA | No | Rapeseed (Westar) | Hyphal branching | [39] |
| sscle_03g031480 | SS1G_13315 | Isolated from diseased rapeseed, Alberta, Canada | thr1 | Trihydroxynaphthalene reductase | Deletion | + | + | NA | - | NA | NA | No | Rapeseed (Westar) | Hyphal branching | [39] |
| sscle_03g031520 | SS1G_13322 | 1980, Nebraska, U.S.A.; UF1, Florida; WMA, Washington, USA | pks13 | Polyketide synthase | Deletion by CRISPR | - | - | NA | - | + | NA | No | Soybean, canola, tomato (Better Boy), faba bean (Windsor) and pea (Sugar Daddy) | Pigmentation of compound appressoria | [40] |
| sscle_03g031670 | SS1G_13339 | 274, Nebraska, USA | pth2 | Peroxysomal carnitine acetyl transferase | Deletion | + | + | + | + | + | NA | No | Soybean | Oxalic acid accumulation | [41] |
| sscle_04g034810 | SS1G_02462 | Isolated from an infected canola stem, Alberta, Canada | axp | Arabinofuranosidase/β-xylosidase | Deletion | NA | NA | NA | + | NA | NA | Yes | Canola | [42] | |
| sscle_04g034960 | SS1G_02486 | Sunf-M, Inner Mongolia, China | caf1 | Secreted protein with a putative Ca2+-binding EF-hand motif | RNAi, T-DNA | + | + | - | + | + | + | Yes | Arabidopsis, rapeseed, pak choi cabbage, hot pepper, tomato, cucumber and soybean | [43] | |
| sscle_04g037170 | SS1G_02784 | 1980, Nebraska, USA | scat1 | Type A catalase | Deletion | + | + | NA | + | NA | NA | No | Tomato (Garden Peach) | Modulation of ROS | [44] |
| sscle_04g038020 | SS1G_02904 | Ep-1PNA367, Heilongjiang, China | cvnh | Secreted protein | RNAi | + | + | NA | + | NA | NA | Yes | Arabidopsis | [45] | |
| sscle_05g046240 | SS1G_05917 | SUN-F-M, Inner Mongolia, China | sl2 | Cell wall protein | RNAi | - | + | NA | - | NA | NA | Yes | Rapeseed and Arabidopsis | Cellular integrity | [46] |
| sscle_05g046390 | SS1G_05899 | 1980, Nebraska, USA | trr1 | Thioredoxin reductase | RNAi | NA | + | NA | + | NA | NA | No | Arabidopsis and tobacco | Oxidative stress tolerance | [47] |
| sscle_05g046790 | SS1G_05839 | 1980, Nebraska, USA | bi1 | Bax inhibitor-1 protein | RNAi | + | - | - | + | NA | NA | No | Arabidopsis and tomato | Stress responses, hyphal tip branching | [48] |
| sscle_05g046830 | SS1G_05834 | JRUF1, Florida, USA | foxe2 | Forkhead-box transcription factor family gene | Deletion, T-DNA | - | - | NA | - | - | NA | No | Tomato | Apothecial development | [49] |
| sscle_05g048220 | SS1G_05661 | 1980, Nebraska, USA | nox1 | NADPH oxidase | RNAi | - | + | + | + | NA | NA | No | Tomato (Rutger) | ROS regulation | [50] |
| sscle_06g049430 | SS1G_07404 | 1980, Nebraska, USA | rhs1 | Rearrangement hot spot repeat-containing protein | RNAi | + | + | - | + | + | NA | Yes | Arabidopsis and canola (Zhongshuang 9) | [51] | |
| sscle_06g049780 | SS1G_07360 | 1980, Nebraska, USA | fkh1 | Atypical forkhead (FKH)-box-containing protein | RNAi | + | + | NA | + | NA | NA | No | Tomato | Cellular integrity | [52] |
| sscle_06g049830 | SS1G_07355 | 1980, Nebraska, USA | pac1 | pH-Responsive transcription factor | Deletion | - | + | + | + | NA | NA | No | Arabidopsis and tomato (Bonnie Best) | [53] | |
| sscle_06g049890 | SS1G_07345 | NGA4, Anhui, China | pemg1 | Elicitor-homologous protein | RNAi | + | NA | - | + | + | NA | No | Oilseed rape and soybean | Negative regulator of growth and virulence | [54] |
| sscle_06g051560 | SS1G_07136 | UF1, Florida, USA | ste12 | Downstream transcription factor of MAPK pathway | RNAi | + | + | NA | + | + | NA | No | Bush bean and tomato | [13] | |
| sscle_07g055970 | SS1G_03527 | SUN-F-M, Inner Mongolia, China | hex1 | Woronin body major protein | RNAi | - | + | NA | NA | NA | NA | No | Cellular integrity | [46] | |
| sscle_07g058030 | SS1G_03252 | UF1, Florida, USA | ams2 | Cell-cycle-regulated GATA transcription factor | RNAi | + | - | NA | + | + | NA | No | Common bean | Chromosome segregation, number and distribution of sclerotia | [55] |
| sscle_07g058620 | SS1G_03171 | 1980, Nebraska, USA | pka1 | Protein kinase A | Deletion | - | - | - | NA | NA | NA | No | PKA activity | [56] | |
| sscle_07g059700 | MK992913 | 1980, Nebraska, USA | qdo | Quercetin dioxygenase gene | Deletion | - | NA | NA | + | NA | NA | Yes | Arabidopsis | Flavonol degradation | [57] |
| sscle_08g064670 | SS1G_05163 | 1980, Nebraska, USA | cry1 | Cryptochrome family CRY-DASH ortholog | Deletion | - | - | NA | - | NA | NA | No | Arabidopsis and tomato | Sclerotial mass, response to UV | [58] |
| sscle_08g065550 | SS1G_05284 | NGA4, Anhui, China | nacα | Nascent polypeptide-associated complex α-subunit | RNAi | - | - | - | + | NA | NA | No | Oilseed rape and tobacco | [59] | |
| sscle_08g066770 | SS1G_05445 | 1980, Nebraska, USA | smk3 | Slt2 ortholog | Deletion | + | + | NA | + | + | NA | No | Canola (Westar) | Cuticle penetration, cell wall integrity | [60] |
| sscle_08g067830 | SS1G_05588 | 1980, Nebraska, U.S.A. | mads | MADS-box proteins | RNAi | + | - | NA | + | NA | NA | No | Tomato | [61] | |
| sscle_08g068500 | SS1G_14133 | Ep-1PNA367, Heilongjiang, China | itl | Integrin-like protein | RNAi | + | + | NA | + | NA | + | Yes | Arabidopsis and canola | Hyphal branching, suppression of host defense | [62] |
| sscle_08g068530 | SS1G_14127 | 1980, Nebraska, USA | ggt1 | γ-Glutamyl transpeptidase | Deletion | - | + | NA | - | + | NA | Yes | Tomato | [63] | |
| sscle_09g069850 | SS1G_10796 | 1980, Nebraska, USA | odc2 | Oxalate decarboxylases | Deletion | - | - | + | + | + | NA | Yes | Common bean (Bush Blue Lake 47), soybean (Harosoy), tomato (Bonnie Best) and celery | [64] | |
| sscle_10g075560 | SS1G_08218 | WMA1, Washington, USA | oah | Oxaloacetate acetylhydrolase | T-DNA | - | - | + | + | NA | NA | No | Faba bean (Broad Windsor), pea (Guido), green bean (Great North Tara) and soybean (Skylla) | [65] | |
| sscle_10g075560 | SS1G_08218 | 1980, Nebraska, USA | oah1 | Oxaloacetate acetylhydrolase | Deletion | - | + | + | + | + | NA | No | Tomato (Bonnie Best), common bean (Bush Blue Lake 47), soybean (Harosoy), canola, Arabidopsis and sunflower | [66] | |
| sscle_10g075560 | SS1G_08218 | 1980, Nebraska, U.S.A.; UF1, Florida; WMA, Washington, USA | oah1 | Oxaloacetate acetylhydrolase | Deletion by CRISPR | - | + | + | + | + | NA | No | Soybean, canola, tomato (Better Boy), faba bean (Windsor), and pea (Sugar Daddy) | [40] | |
| sscle_10g077630 | SS1G_08489 | 1980, Nebraska, USA | pph1 | Type 2A Ser/Thr phosphatase catalytic subunit PP2Ac | RNAi | + | + | NA | NA | NA | NA | No | [67] | ||
| sscle_10g079320 | SS1G_14065 | 1980, Nebraska, USA | ssp1 | Development-specific protein | Deletion | - | - | NA | NA | NA | NA | No | Resistance to glycoside-containing antibiotics | [68] | |
| sscle_11g082700 | SS1G_07871 | 1980, Nebraska, USA | rgb1 | Type 2A Ser/Thr phosphatase B subunit | RNAi | - | + | NA | + | + | NA | No | Arabidopsis and tomato (Bonny Best) | MAPK pathway | [67] |
| sscle_11g083230 | SS1G_07798 | SUN-F-M, Inner Mongolia, China | gpd | glyceraldehyde-3-phosphate dehydrogenase | RNAi | NA | + | NA | NA | NA | NA | No | [46] | ||
| sscle_11g083680 | SS1G_07749 | 1980, Nebraska, USA | xyl1 | Endo-β-1,4-xylanase | Deletion | + | + | NA | + | NA | NA | Yes | Canola and Arabidopsis | Apothecia formation | [69] |
| sscle_11g083950 | SS1G_07715 | 1980, Nebraska, USA | sac1 | Adenylate cyclase | Deletion | + | + | - | + | NA | NA | No | Tomato (Bonnie Best) | Apothecia production, cAMP-signaling | [70] |
| sscle_12g087830 | SS1G_11172 | 1980, Nebraska, USA | nox2 | NADPH oxidase | RNAi | - | + | - | - | NA | NA | No | Tomato (Rutger) | ROS regulation | [50] |
| sscle_12g090900 | SS1G_11866 | 1980, Nebraska, USA | smk1 | ERK -type MAP kinase | RNAi | + | + | NA | NA | NA | NA | No | pH-dependent regulation | [71] | |
| sscle_14g099710 | SS1G_08814 | 1980, Nebraska, USA | odc1 | Oxalate decarboxylases | Deletion | - | NA | NA | - | NA | NA | Yes | Common bean (Bush Blue Lake 47), soybean (Harosoy) and tomato (Bonnie Best) | [64] | |
| sscle_16g107670 | SS1G_10096 | 1980, Nebraska, USA | cp1 | Cerato-platanin protein | Deletion | - | - | NA | + | NA | + | Yes | Arabidopsis and tobacco | Interaction with PR1 | [72] |
| sscle_16g107670 | SS1G_10096 | FXGD2, Anhui, China | sm1 | Cerato-platanin protein | RNAi | + | + | - | + | NA | NA | Yes | Tobacco, oilseed rape and soybean | [73] | |
| sscle_16g107930 | SS1G_10135 | 1980, Nebraska, USA | fdh1 | Formaldehyde dehydrogenase | Deletion | - | + | NA | + | + | NA | No | Common bean and tomato | Osmotic oxidative stress resistance | [74] |
| sscle_16g109570 | ANQ80447 | 1980, Nebraska, USA | nsd1 | GATA-type Ivb zinc-finger transcription factor | Deletion | - | - | NA | + | + | NA | No | Tomato (Bonnie Best), celery and tomato | Ascogonia formation, apothecium development | [75] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, S.; Xu, Y.; Hoy, R.; Zhang, J.; Qin, L.; Li, X. The Notorious Soilborne Pathogenic Fungus Sclerotinia sclerotiorum: An Update on Genes Studied with Mutant Analysis. Pathogens 2020, 9, 27. https://doi.org/10.3390/pathogens9010027
Xia S, Xu Y, Hoy R, Zhang J, Qin L, Li X. The Notorious Soilborne Pathogenic Fungus Sclerotinia sclerotiorum: An Update on Genes Studied with Mutant Analysis. Pathogens. 2020; 9(1):27. https://doi.org/10.3390/pathogens9010027
Chicago/Turabian StyleXia, Shitou, Yan Xu, Ryan Hoy, Julia Zhang, Lei Qin, and Xin Li. 2020. "The Notorious Soilborne Pathogenic Fungus Sclerotinia sclerotiorum: An Update on Genes Studied with Mutant Analysis" Pathogens 9, no. 1: 27. https://doi.org/10.3390/pathogens9010027
APA StyleXia, S., Xu, Y., Hoy, R., Zhang, J., Qin, L., & Li, X. (2020). The Notorious Soilborne Pathogenic Fungus Sclerotinia sclerotiorum: An Update on Genes Studied with Mutant Analysis. Pathogens, 9(1), 27. https://doi.org/10.3390/pathogens9010027