Impact of a Live Attenuated Classical Swine Fever Virus Introduced to Jeju Island, a CSF-Free Area

Abstract
1. Introduction
2. Results
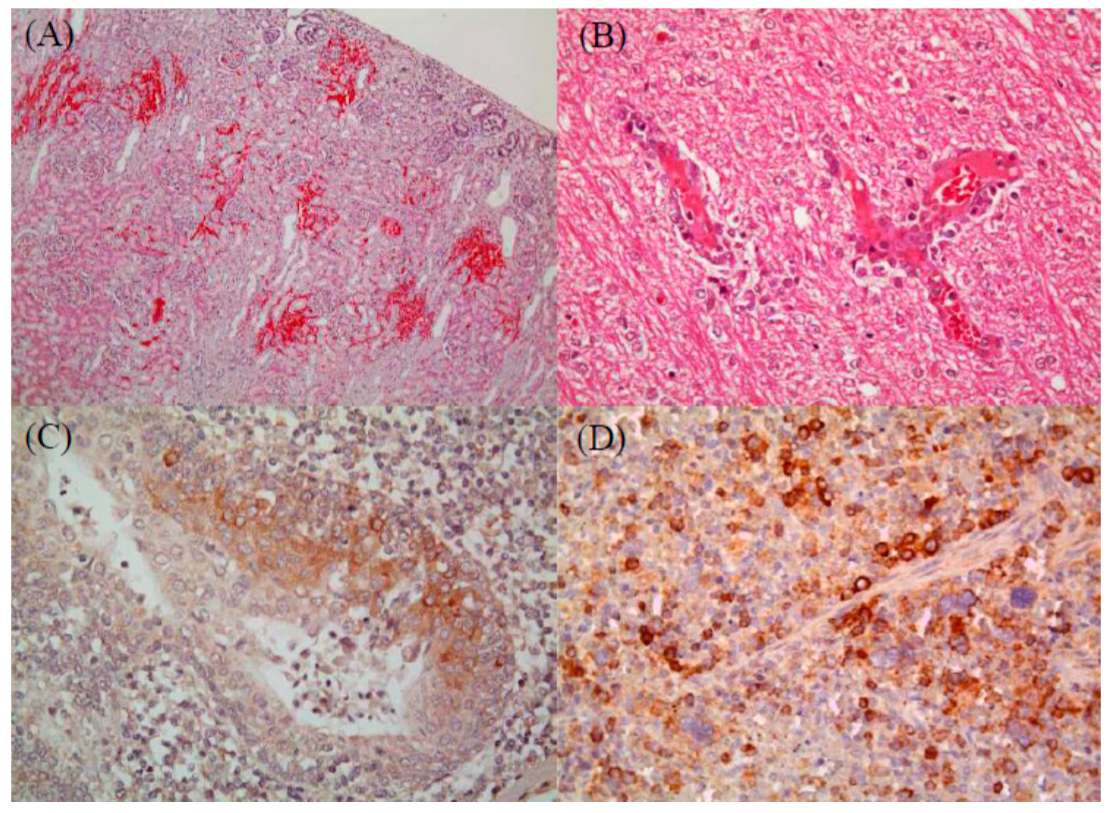
2.1. Histopathological Analysis to Detect Infection by CSFV (Jeju LOM) Alone or Co-Infection with CSFV (Jeju LOM) and Other Pathogens
2.2. Detection of Antibodies on Pig Farms Exposed to the Jeju LOM Strain
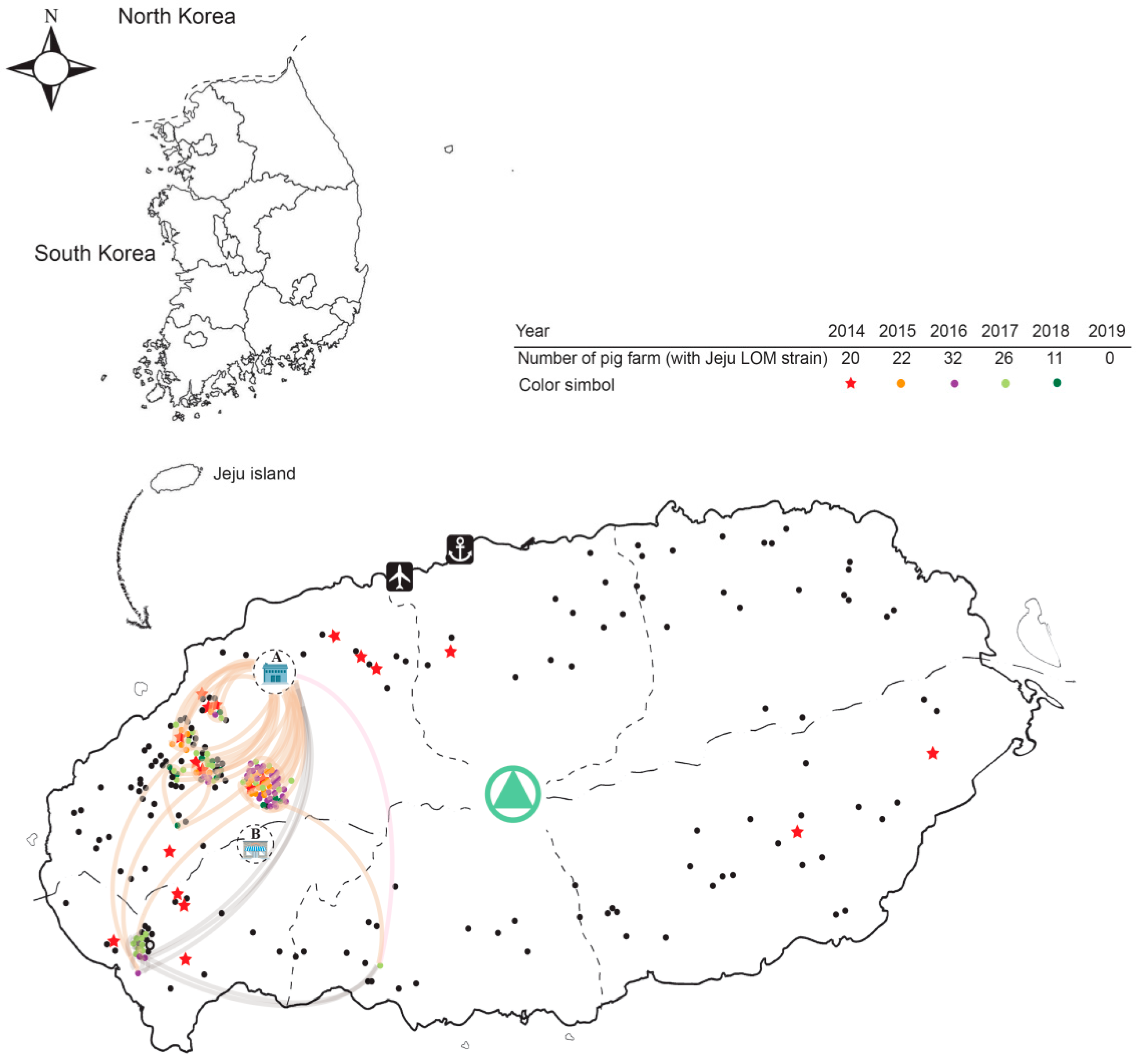
2.3. Farm-Slaughterhouse-Farm Transmission
2.4. Pig-to-Pig Transmission and Reproduction Rate (R)
2.5. Comparison of LOM Strain Genome Sequences
2.6. Root-to-Top Divergence and Positive Selection Analyses
2.7. Geographic Distance and MCC Tree Analysis
3. Discussion
4. Materials and Methods
4.1. Detection of Histopathologic Lesions and Immunohistochemical Straining Analysis
4.2. Detection of Anti-CSF Antibodies
4.3. Environmental Samples Taken from a Slaughterhouse
4.4. Horizontal Transmission between Pigs
4.5. Genomic Analysis of Commercial LOM Vaccine Strains and Jeju LOM Strains
4.6. Maximum Clade Credibility Tree
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Ethical approval and consent to participate
References
- Becher, P.; Avalos Ramirez, R.; Orlich, M.; Cedillo Rosales, S.; Konig, M.; Schweizer, M.; Stalder, H.; Schirrmeier, H.; Thiel, H.J. Genetic and antigenic characterization of novel pestivirus genotypes: Implications for classification. Virology 2003, 311, 96–104. [Google Scholar] [CrossRef]
- Rice, C.M. Flaviviridae: The viruses and their replication. In Fundamental Virology, 3rd ed.; Knipe, D.M., Howley, P., Eds.; Lippincott Raven: Philadelphia, PA, USA, 1996; pp. 931–959. [Google Scholar]
- Paton, D.J.; McGoldrick, A.; Greiser-Wilke, I.; Parchariyanon, S.; Song, J.Y.; Liou, P.P.; Stadejek, T.; Lowings, J.P.; Björklund, H.; Belák, S. Genetic typing of classical swine fever virus. Vet. Microbiol. 2000, 73, 137–157. [Google Scholar] [CrossRef]
- Edwards, S.; Fukusho, A.; Lefevre, P.C.; Lipowski, A.; Pejsak, Z.; Roehe, P.; Westergaard, J. Classical swine fever: The global situation. Vet. Microbiol. 2000, 73, 103–119. [Google Scholar] [CrossRef]
- Morilla, A.; Carvajal, M.A. Experiences with classical swine fever vaccination in Mexico. In Trends in Emerging Viral Infections of Swine; Morilla, A., Yoon, K.J., Zimmerman, J.J., Eds.; Iowa State Press, Blackwell Publishing Company: Ames, IA, USA, 2002; pp. 159–164. [Google Scholar]
- Pijoan, C.; Campos, M.; Ochoa, G. Effect of a hog cholera vaccine strain on the bactericidal activity of porcine alveolar macrophages. Rev. Latinoam. Microbiol. 1980, 22, 69–71. [Google Scholar] [PubMed]
- Lim, S.I.; Song, J.Y.; Kim, J.; Hyun, B.H.; Kim, H.Y.; Cho, I.S.; Kim, B.; Woo, G.H.; Lee, J.B.; An, D.J. Safety of classical swine fever virus vaccine strain LOM in pregnant sows and their offspring. Vaccine 2016, 34, 2021–2026. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.I.; Jeoung, H.Y.; Kim, B.; Song, J.Y.; Kim, J.; Kim, H.Y.; Cho, I.S.; Woo, G.H.; Lee, J.B.; An, D.J. Impact of porcine reproductive and respiratory syndrome virus and porcine circovirus-2 infection on the potency of the classical swine fever vaccine (LOM strain). Vet. Microbiol. 2016, 193, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Song, J.Y.; Tark, D.S.; Lim, S.I.; Choi, E.J.; Park, C.K.; Lee, B.Y.; Wee, S.H.; Bae, Y.C.; Lee, O.S.; et al. Feed comtaminated with classical swine fever vaccine virus (LOM strain) can induce antibodies to the virus in pigs. Vet. Rec. 2008, 162, 12–17. [Google Scholar] [CrossRef]
- Wieringa-Jelsma, T.; Quak, S.; Loeffen, W.L. Limited BVDV transmission and full protection against CSFV transmission in pigs experimentally infected with BVDV type 1b. Vet. Microbiol. 2006, 118, 26–36. [Google Scholar] [CrossRef]
- Kang, M.I. Epidemiological Survey Report on the Detection of CSF Vaccine (LOM Strain) in Jeju Island; National Veterinary Research Quarantine Service (NVRQS) Publishing: Anyang, Korea, 2005; pp. 1–137. [Google Scholar]
- Tamura, T.; Sakoda, Y.; Yoshino, F.; Nomura, T.; Yamamoto, N.; Sato, Y.; Okamatsu, M.; Ruggli, N.; Kida, H. Selection of classical swine fever virus with enhanced pathogenicity reveals synergistic virulence determinants in E2 and NS4B. J. Virol. 2012, 86, 8602–8613. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Nagashima, N.; Ruggli, N.; Summerfield, A.; Kida, H.; Sakoda, Y. Npro of classical swine fever virus contributes to pathogenicity in pigs by preventing type I interferon induction at local replication sites. Vet. Res. 2014, 45, 47. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Li, L.; Zhao, Y.; Tu, J.; Pan, Z. Identification of two amino acids within E2 important for the pathogenicity of chimeric classical swine fever virus. Virus. Res. 2016, 211, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Holinka, L.G.; Fernandez-Sainz, I.; Sanford, B.; O'Donnell, V.; Gladue, D.P.; Carison, J.; Lu, Z.; Risatti, G.R.; Borca, M.V. Development of an improved live attenuated antigenic marker CSF vaccine strain candidate with an increased genetic stability. Virology 2014, 471, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Risatti, G.R.; Holinka, L.G.; Fernandez Sainz, I.; Carrillo, C.; Lu, Z.; Borca, M.V. N-linked glycosylation status of classical swine fever virus strain Brescia E2 glycoprotein influences virulence in swine. J. Virol. 2007, 81, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.; Nagai, H.; Katayama, K.; Tsutsui, M.; Tanabayashi, K.; Takeuchi, K.; Hishiyama, M.; Saitoh, A.; Takagi, M.; Gotoh, K.; et al. Comparison of the entire nucleotide and deduced amino acid sequences of the attenuated hog cholera vaccine strain GPE- and the wild-type parental strain ALD. Arch. Virol. 1995, 140, 1385–1391. [Google Scholar] [CrossRef] [PubMed]
- Larochelle, R.; Antaya, M.; Morin, M.; Magar, R. Typing of porcine circovirus in clinical specimens by multiplex PCR. J. Virol. Methods 1999, 80, 69–75. [Google Scholar] [CrossRef]
- Drew, T. Classical swine fever (hog cholera). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals: Mammals, Birds and Bees, 6th ed.; Office International des Epizooties (OIE) , Ed.; OIE: Paris, France, 2008; pp. 1092–1106. [Google Scholar]
- Dietz, K. The estimation of the basic reproduction number for infectious diseases. Stat. Methods Med. Res. 1993, 2, 23–41. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Lam, T.T.; Carvalho, L.M.; Pybus, O.G. Exploring the temporal structure of heterochronous sequences using TempEst. Virus Evol. 2016, 2, vew007. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J. Tracer v1.5. 2009. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 20 November 2019).
- Rambaut, A.; Drummond, A.J. TreeAnnotator v1.7.0. 2012. Available online: http://beast.bio.ed.ac.uk (accessed on 20 November 2019).
- Rambaut, A. FigFree v.1.4.0. 2012. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 20 November 2019).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infection Pattern with CSFV (Jeju LOM Strain) | Pathogens (or Diseases) | Year | |||||
|---|---|---|---|---|---|---|---|
| 2014 | 2015 | 2016 | 2017 | 2018 | |||
| Suckling piglets | Co-infection with other pathogens | Porcine epidemic diarrhea (PED) | 1 | 2 | |||
| Rotavirus enteritis | 2 | 2 | |||||
| Rota viral enteritis + Strep or Staphylo pneumonia | 1 | 1 | |||||
| Colibacillosis | 4 | 2 | |||||
| Clostridium difficile associated disease | 2 | 1 | |||||
| Clostridium enteritis (+ exudative epidermitis ) | 1 | 2(1) | |||||
| Porcine reproductive and respiratory syndrome (PRRS) | 2 | ||||||
| PRRS + Pasteurella pneumonia | 1 | ||||||
| Streptococcal infection (abscess etc.) | 1 | 3 | 3 | ||||
| Staphylococcal infection | 2 | ||||||
| Viral encephalitis suspect or bacterial meningitis | 1 | 2 | |||||
| Infection (only Jeju LOM) | 3 | 6 | 5 | ||||
| Weaning piglets | Co-infection with other pathogens | PRRS | 1 | ||||
| PRRS + Streptococcal pneumonia | 1 | ||||||
| PRRS + PCV-2 + APP | 1 | ||||||
| Actinobacillus pleuropneumoniae (APP) | 1 | 2 | |||||
| APP + PCV-2 | 1 | ||||||
| PCV-2 (+ Salmonellosis/Staphylo) | 3 | ||||||
| Pasteurella (or E. coli) pneumonia | 1 | 1 | |||||
| Streptococcal infection | 4 | ||||||
| Rotavirus enteritis | 2 | ||||||
| Salmonellosis (Colibacillosis) | 2(1) | ||||||
| Total | 2 | 2 | 14 | 28(2) | 24 | ||
| Infection Pattern with CSFV (Jeju LOM Strain) | Internal Organs (No. of Positive Pigs/No. Tested) | Total | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Tonsil | Lymph Node | Spleen | Lung | Heart | Kidney | Liver | GI Tract | CNS | |||
| Suckling piglets | Co-infection | 11/24 | 3/30 | 8/34 | 4/29 | 0/32 | 2/34 | 1/34 | 2/33 | 0/26 | 17/34 |
| Infection (only Jeju LOM) | 3/11 | 3/10 | 3/14 | 2/12 | 0/13 | 0/14 | 0/13 | 0/13 | 0/12 | 8/14 | |
| Subtotal | 14/35 | 6/40 | 11/48 | 6/41 | 0/45 | 2/48 | 1/47 | 2/46 | 0/38 | 25/48 | |
| Weaning pigs | Co-infection | 1/20 | 4/20 | 3/18 | 3/20 | 0/20 | 0/20 | 0/19 | 0/21 | 0/15 | 6/21 |
| Aborted fetuses | Infection (only Jeju LOM) | NT | NT | 0/8 | 0/8 | 0/8 | 1/8 | 0/8 | NT | 0/8 | 1/8 |
| Pig Farm | Antibody Positive Ratio (%) against CSFV (Jeju LOM Strain) | Average Antibody Titers (Log 2) for CSFV (Jeju LOM Strain) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sow | Piglet (10–20 days) | Pig (40–60 days) | Pig (90–120 days) | Pig (150–180 days) | Sow | Piglet (10–20 days) | Pig (40–60 days) | Pig (90–120 days) | Pig (150–180 days) | |
| A | 80 | 40 | 30 | 0 | 0 | 9.5 | 7.0 | 9.1 | - | - |
| B | 90 | 95 | 100 | 80 | 0 | 10.0 | 10.0 | 7.1 | 5.2 | - |
| C | 100 | 100 | 40 | 40 | 90 | 8.1 | 6.1 | 5.5 | 5.5 | 8.0 |
| D | 100 | 90 | 0 | 10 | 70 | 9.1 | 6.3 | - | 5.6 | 8.5 |
| E | 90 | 95 | 0 | 0 | 40 | 9.5 | 6.2 | - | - | 8.9 |
| F | 80 | 65 | 0 | 0 | 10 | 9.6 | 6.0 | - | - | 7.1 |
| G | 70 | 60 | 0 | 20 | 60 | 10.2 | 7.7 | - | 5.8 | 4.7 |
| qRT-PCR | Immuno Histo Chemistry (IHC) Staining | |||||
|---|---|---|---|---|---|---|
| Ct Value (Range) | Jeju LOM Strain * TCID50 (Log 10) | Sample no. | Negative Samples | Weak Positive Samples | Strong Positive Samples | ** No Test |
| >40 | <1.0 | 91 | 43 | 48 | ||
| 36.1–39.9 | 1.0–2.0 | 26 | 9 | 1 | 16 | |
| 30.5–36.0 | 2.0–3.0 | 21 | 4 | 8 | 9 | |
| 26.7–30.4 | 3.0–4.0 | 92 | 12 | 22 | 3 | 55 |
| 24.4–26.6 | 4.0–5.0 | 11 | 1 | 9 | 1 | |
| <24.3 | >5.0 | 1 | 1 | |||
| Group | Number of Pigs Inoculated with the Jeju LOM Strain | Non-Inoculated Pigs Exposed to Jeju LOM Virus Inoculated Pigs (1 DPI a) | Pigs Detected with Jeju LOM Strain Antigens and/or Antibodies in Non-Inoculated Pigs (45 DPI) | Transmission Probability Estimate | |
|---|---|---|---|---|---|
| R0 b | 95% CI c | ||||
| 1 | 6 | 6 | 2 | 1.22 | 0.980–1.765 |
| 2 | 6 | 6 | 0 | 0.00 | NA d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choe, S.; Kim, J.-H.; Kim, K.-S.; Song, S.; Kang, W.-C.; Kim, H.-J.; Park, G.-N.; Cha, R.M.; Cho, I.-S.; Hyun, B.-H.; et al. Impact of a Live Attenuated Classical Swine Fever Virus Introduced to Jeju Island, a CSF-Free Area. Pathogens 2019, 8, 251. https://doi.org/10.3390/pathogens8040251
Choe S, Kim J-H, Kim K-S, Song S, Kang W-C, Kim H-J, Park G-N, Cha RM, Cho I-S, Hyun B-H, et al. Impact of a Live Attenuated Classical Swine Fever Virus Introduced to Jeju Island, a CSF-Free Area. Pathogens. 2019; 8(4):251. https://doi.org/10.3390/pathogens8040251
Chicago/Turabian StyleChoe, SeEun, Jae-Hoon Kim, Ki-Sun Kim, Sok Song, Wan-Choul Kang, Hyeon-Ju Kim, Gyu-Nam Park, Ra Mi Cha, In-Soo Cho, Bang-Hun Hyun, and et al. 2019. "Impact of a Live Attenuated Classical Swine Fever Virus Introduced to Jeju Island, a CSF-Free Area" Pathogens 8, no. 4: 251. https://doi.org/10.3390/pathogens8040251
APA StyleChoe, S., Kim, J.-H., Kim, K.-S., Song, S., Kang, W.-C., Kim, H.-J., Park, G.-N., Cha, R. M., Cho, I.-S., Hyun, B.-H., Park, B.-K., & An, D.-J. (2019). Impact of a Live Attenuated Classical Swine Fever Virus Introduced to Jeju Island, a CSF-Free Area. Pathogens, 8(4), 251. https://doi.org/10.3390/pathogens8040251