Zebrafish are Resistant to Staphylococcus aureus Endophthalmitis

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
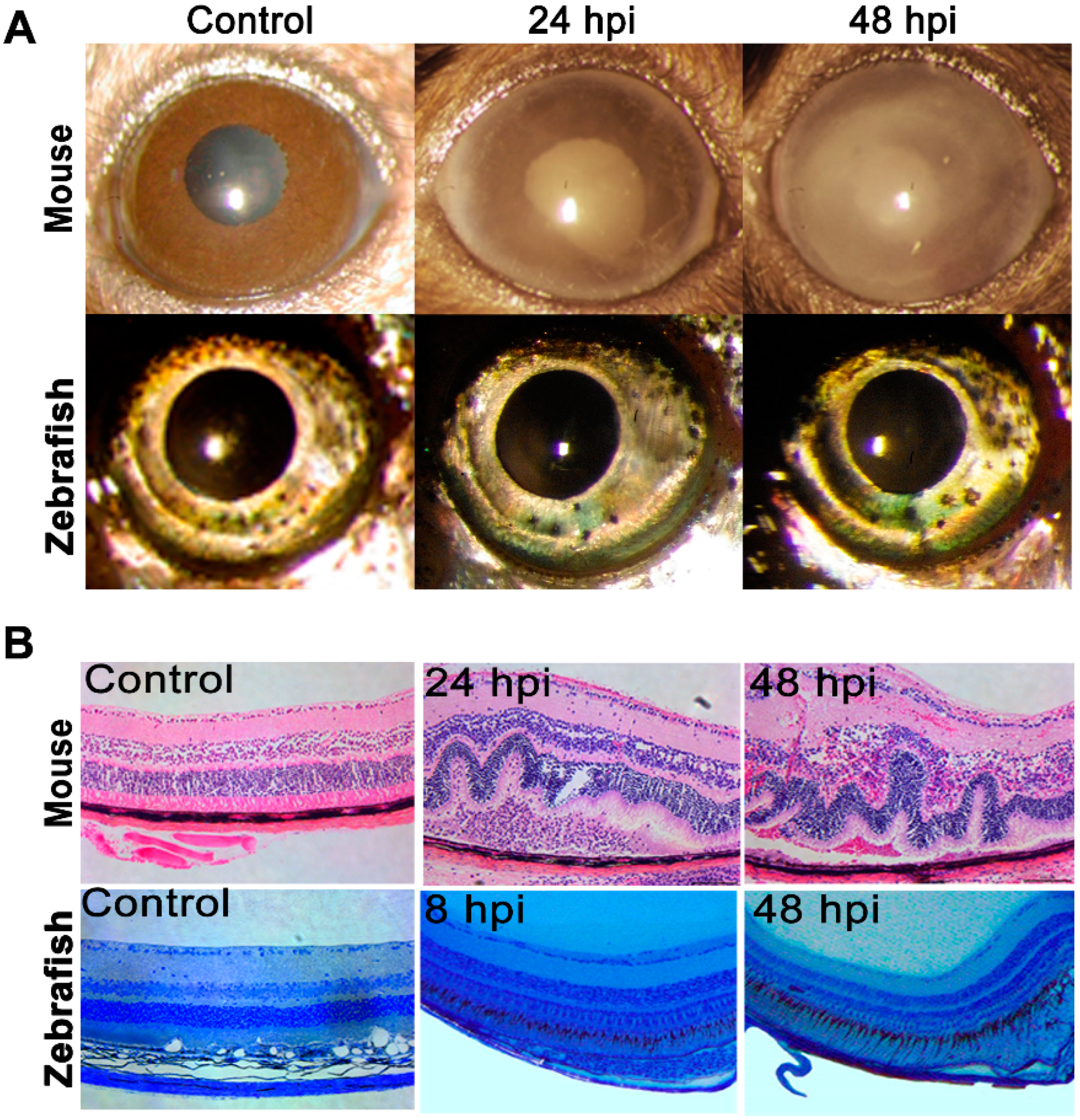
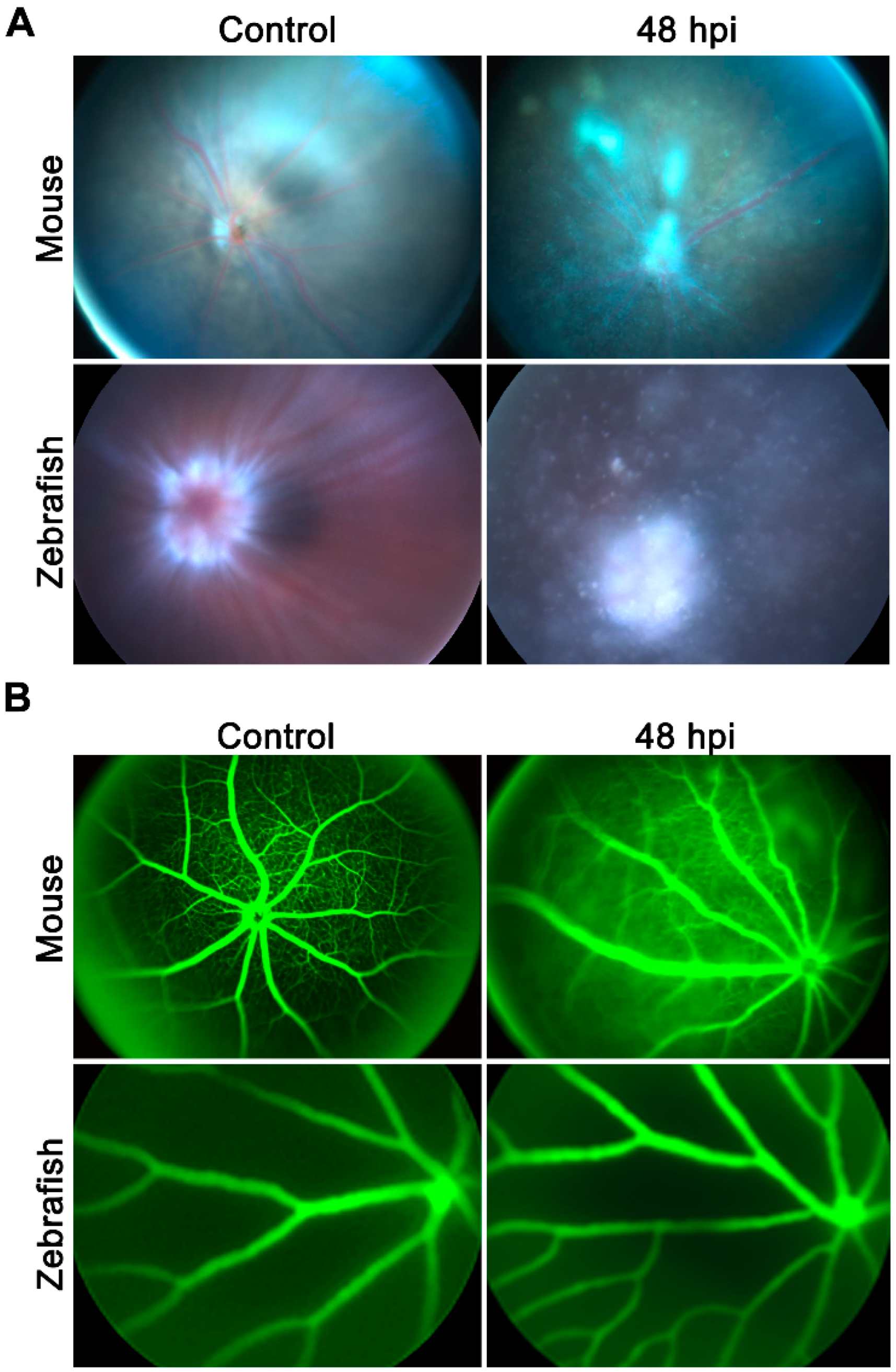
2.1. Zebrafish Did Not Develop Staphylococcal Endophthalmitis
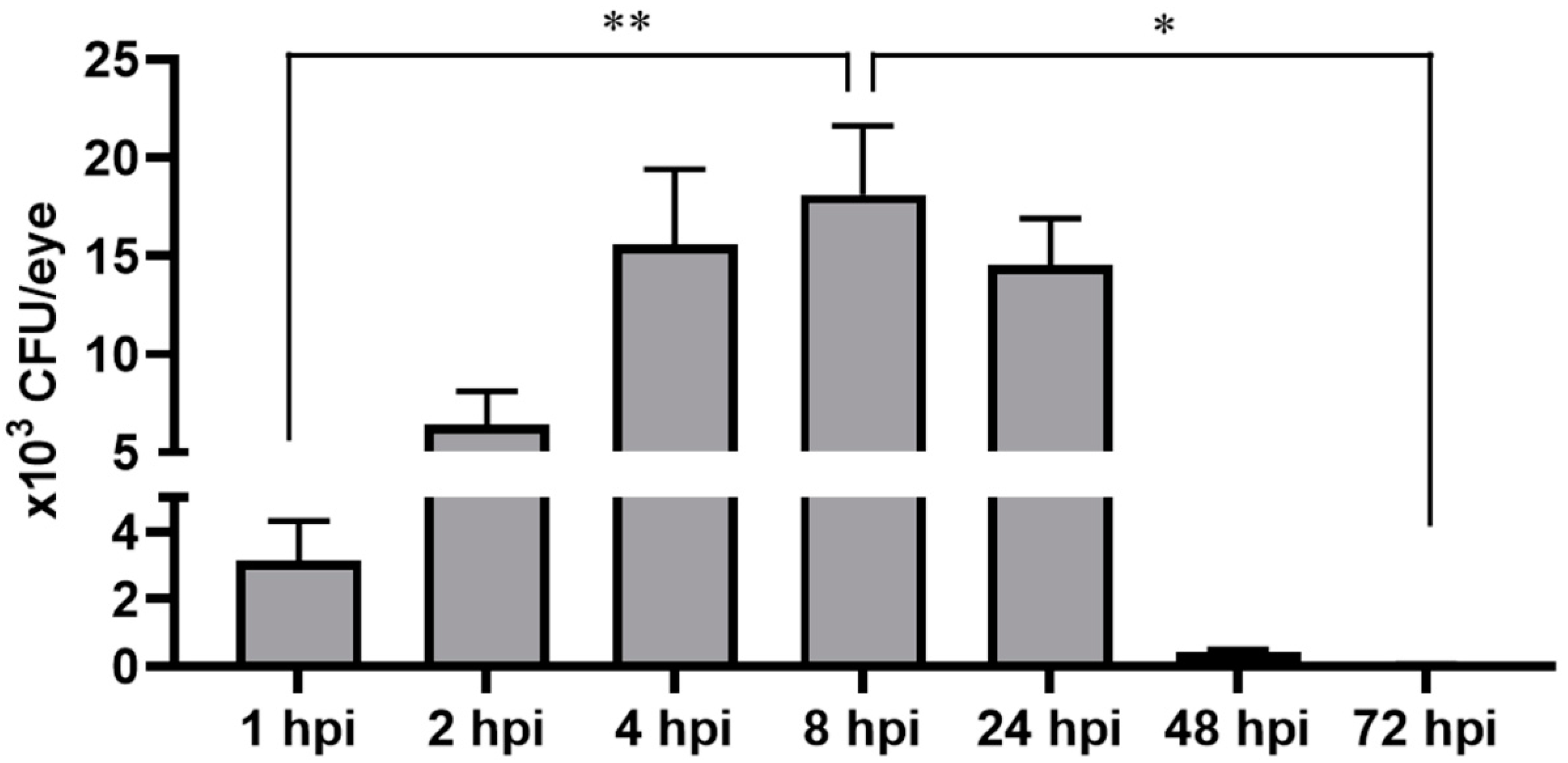
2.2. Zebrafish Rapidly Cleared Bacterial Burden from Eyes
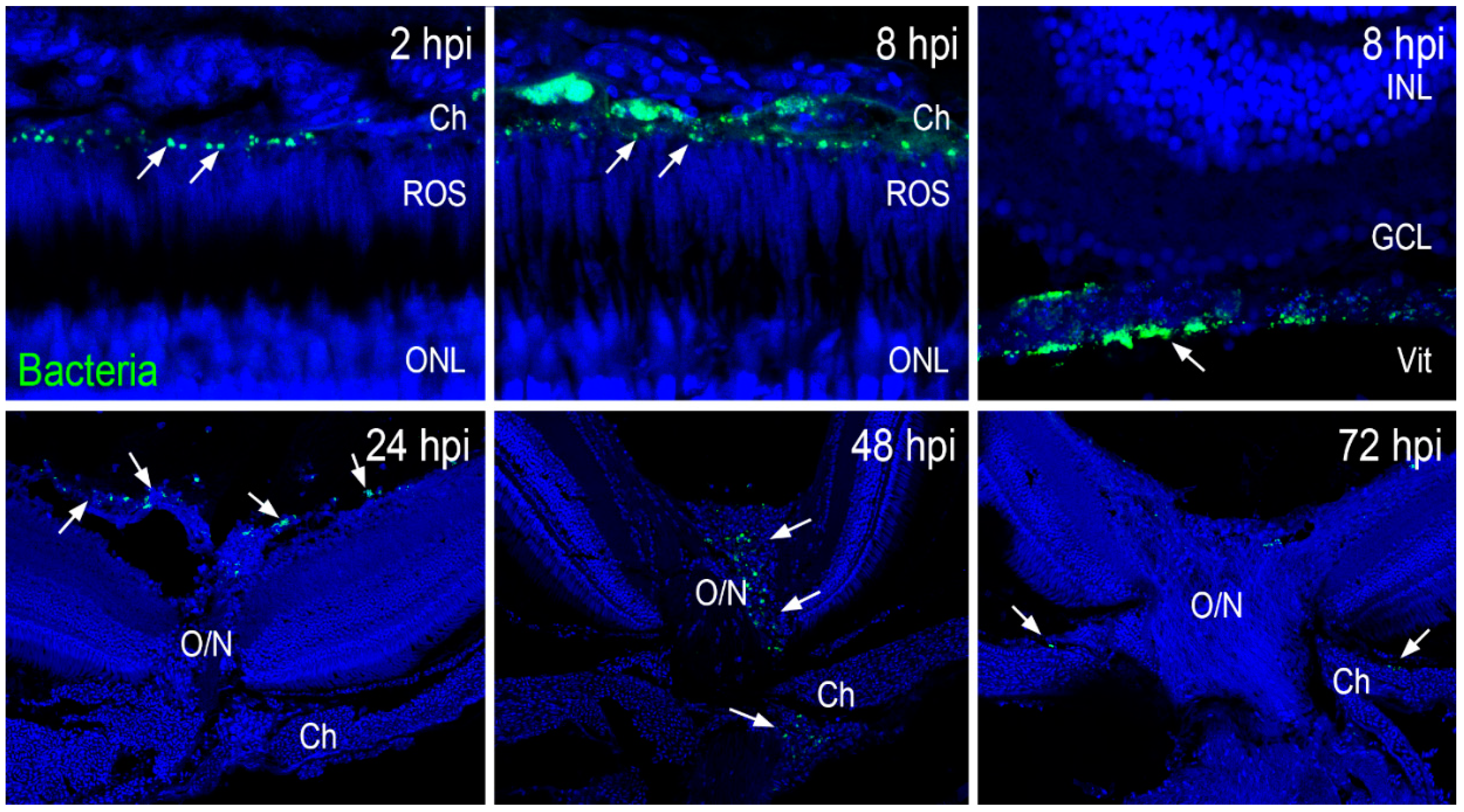
2.3. Zebrafish Eyes Cleared S. aureus through the Vasculature and Optic Nerve Head
2.4. Zebrafish Eyes Recruited Monocytes/Macrophages during S. aureus Infection
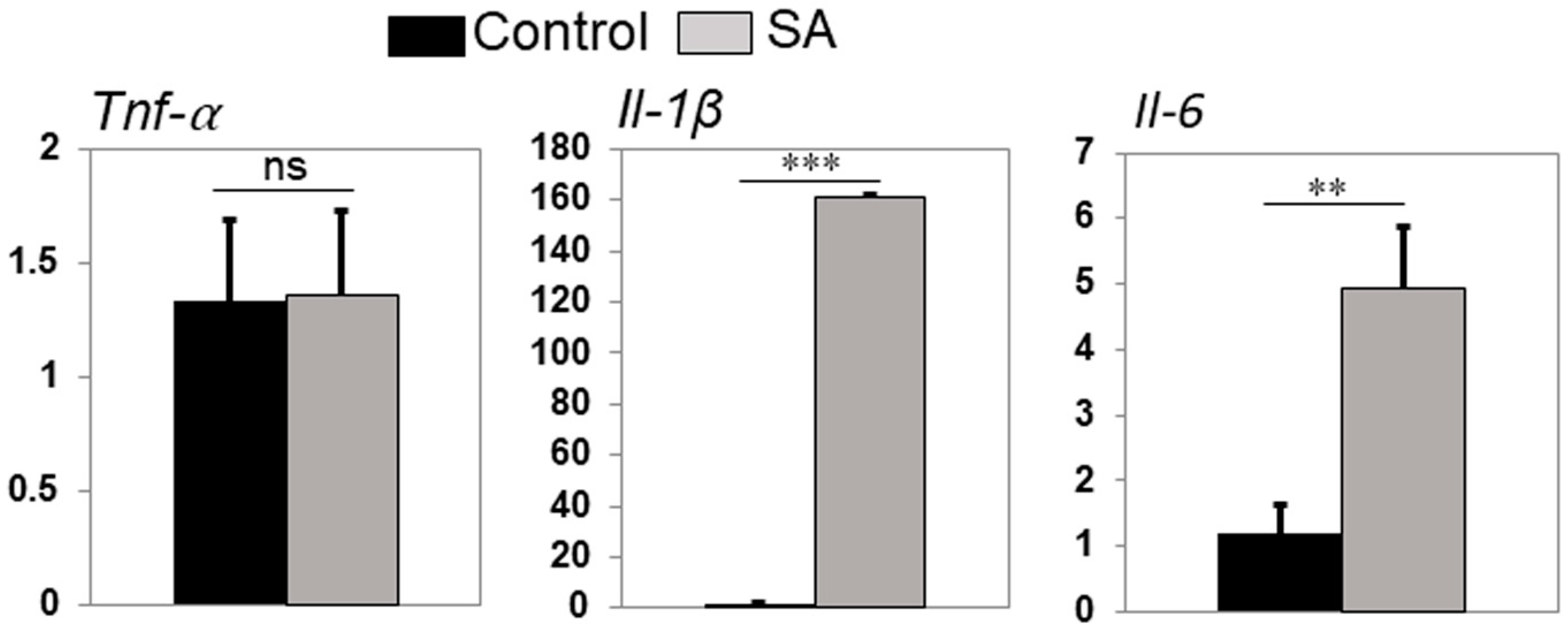
2.5. Infected Zebrafish Eyes Differentially Expressed Inflammatory Mediators
3. Discussion
4. Materials and Methods
4.1. Animals and Bacterial Maintenance
4.2. S. aureus Intraocular Injections
4.3. Bacterial Burden Estimation
4.4. Immunohistochemistry
4.5. qRT-PCR
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Callegan, M.C.; Gilmore, M.S.; Gregory, M.; Ramadan, R.T.; Wiskur, B.J.; Moyer, A.L.; Hunt, J.J.; Novosad, B.D. Bacterial endophthalmitis: Therapeutic challenges and host-pathogen interactions. Prog. Retin. Eye Res. 2007, 26, 189–203. [Google Scholar] [CrossRef]
- Taylor, H. Cataract: How much surgery do we have to do? Br. J. Ophthalmol. 2000, 84, 1–2. [Google Scholar] [CrossRef]
- Chiquet, C.; Cornut, P.L.; Benito, Y.; Thuret, G.; Maurin, M.; Lafontaine, P.O.; Pechinot, A.; Palombi, K.; Lina, G.; Bron, A.; et al. Eubacterial PCR for bacterial detection and identification in 100 acute postcataract surgery endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1971–1978. [Google Scholar] [CrossRef]
- Campbell, R.J.; Bronskill, S.E.; Bell, C.M.; Paterson, J.M.; Whitehead, M.; Gill, S.S. Rapid expansion of intravitreal drug injection procedures, 2000 to 2008: A population-based analysis. Arch. Ophthalmol. 2010, 128, 359–362. [Google Scholar] [CrossRef]
- Sadaka, A.; Durand, M.L.; Gilmore, M.S. Bacterial endophthalmitis in the age of outpatient intravitreal therapies and cataract surgeries: Host-microbe interactions in intraocular infection. Prog. Retin. Eye Res. 2012, 31, 316–331. [Google Scholar] [CrossRef] [PubMed]
- Tosi, M.F. Innate immune responses to infection. J. Allergy Clin. Immunol. 2005, 116, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Talreja, D.; Singh, P.K.; Kumar, A. In Vivo Role of TLR2 and MyD88 Signaling in Eliciting Innate Immune Responses in Staphylococcal Endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2015, 56, 1719–1732. [Google Scholar] [CrossRef] [PubMed]
- Streilein, J.W. Ocular immune privilege: Therapeutic opportunities from an experiment of nature. Nat. Rev. Immunol. 2003, 3, 879–889. [Google Scholar] [CrossRef]
- Moyer, A.L.; Ramadan, R.T.; Novosad, B.D.; Astley, R.; Callegan, M.C. Bacillus cereus-induced permeability of the blood-ocular barrier during experimental endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3783–3793. [Google Scholar] [CrossRef]
- Callegan, M.C.; Booth, M.C.; Jett, B.D.; Gilmore, M.S. Pathogenesis of gram-positive bacterial endophthalmitis. Infect. Immun. 1999, 67, 3348–3356. [Google Scholar]
- Kumar, A.; Kumar, A. Role of Staphylococcus aureus Virulence Factors in Inducing Inflammation and Vascular Permeability in a Mouse Model of Bacterial Endophthalmitis. PLoS ONE 2015, 10, e0128423. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Donovan, D.M.; Kumar, A. Intravitreal Injection of the Chimeric Phage Endolysin Ply187 Protects Mice from Staphylococcus aureus Endophthalmitis. Antimicrob. Agents Chemother. 2014, 58, 4621–4629. [Google Scholar] [CrossRef] [PubMed]
- Neely, M.N.; Pfeifer, J.D.; Caparon, M. Streptococcus-zebrafish model of bacterial pathogenesis. Infect. Immun. 2002, 70, 3904–3914. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.C.; Breen, P.; Britton, S.; Neely, M.N.; Withey, J.H. Quantifying Vibrio cholerae Enterotoxicity in a Zebrafish Infection Model. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [PubMed]
- Runft, D.L.; Mitchell, K.C.; Abuaita, B.H.; Allen, J.P.; Bajer, S.; Ginsburg, K.; Neely, M.N.; Withey, J.H. Zebrafish as a natural host model for Vibrio cholerae colonization and transmission. Appl. Environ. Microbiol. 2014, 80, 1710–1717. [Google Scholar] [CrossRef] [PubMed]
- Howlader, D.R.; Sinha, R.; Nag, D.; Majumder, N.; Mukherjee, P.; Bhaumik, U.; Maiti, S.; Withey, J.H.; Koley, H. Zebrafish as a novel model for non-typhoidal Salmonella pathogenesis, transmission and vaccine efficacy. Vaccine 2016, 34, 5099–5106. [Google Scholar] [CrossRef]
- Hall, C.J.; Boyle, R.H.; Astin, J.W.; Flores, M.V.; Oehlers, S.H.; Sanderson, L.E.; Ellett, F.; Lieschke, G.J.; Crosier, K.E.; Crosier, P.S. Immunoresponsive gene 1 augments bactericidal activity of macrophage-lineage cells by regulating beta-oxidation-dependent mitochondrial ROS production. Cell Metab. 2013, 18, 265–278. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Cao, X.; Jin, X.; Jin, T. Pattern recognition receptors in zebrafish provide functional and evolutionary insight into innate immune signaling pathways. Cell. Mol. Immunol. 2017, 14, 80–89. [Google Scholar] [CrossRef]
- Oosterhof, N.; Boddeke, E.; van Ham, T.J. Immune cell dynamics in the CNS: Learning from the zebrafish. Glia 2015, 63, 719–735. [Google Scholar] [CrossRef]
- Sullivan, C.; Kim, C.H. Zebrafish as a model for infectious disease and immune function. Fish Shellfish Immunol. 2008, 25, 341–350. [Google Scholar] [CrossRef]
- Tobin, D.M.; May, R.C.; Wheeler, R.T. Zebrafish: A see-through host and a fluorescent toolbox to probe host-pathogen interaction. PLoS Pathog. 2012, 8, e1002349. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, J.; Xiong, J.W.; Peng, J. Haploinsufficiency of Def activates p53-dependent TGFbeta signalling and causes scar formation after partial hepatectomy. PLoS ONE 2014, 9, e96576. [Google Scholar] [CrossRef]
- Trede, N.S.; Zapata, A.; Zon, L.I. Fishing for lymphoid genes. Trends Immunol. 2001, 22, 302–307. [Google Scholar] [CrossRef]
- Postlethwait, J.H.; Yan, Y.L.; Gates, M.A.; Horne, S.; Amores, A.; Brownlie, A.; Donovan, A.; Egan, E.S.; Force, A.; Gong, Z.; et al. Vertebrate genome evolution and the zebrafish gene map. Nat. Genet. 1998, 18, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Rajamani, D.; Singh, P.K.; Rottmann, B.G.; Singh, N.; Bhasin, M.K.; Kumar, A. Temporal retinal transcriptome and systems biology analysis identifies key pathways and hub genes in Staphylococcus aureus endophthalmitis. Sci. Rep. 2016, 6, 21502. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Giri, S.; Kumar, A. 5-Aminoimidazole-4-carboxamide ribonucleoside-mediated adenosine monophosphate-activated protein kinase activation induces protective innate responses in bacterial endophthalmitis. Cell. Microbiol. 2016, 18, 1815–1830. [Google Scholar] [CrossRef]
- Gregory, M.; Callegan, M.C.; Gilmore, M.S. Role of bacterial and host factors in infectious endophthalmitis. Chem. Immunol. Allergy 2007, 92, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Pandey, R.K.; Miller, L.J.; Singh, P.K.; Kanwar, M. Muller glia in retinal innate immunity: A perspective on their roles in endophthalmitis. Crit. Rev. Immunol. 2013, 33, 119–135. [Google Scholar] [CrossRef]
- Miller, F.C.; Coburn, P.S.; Huzzatul, M.M.; LaGrow, A.L.; Livingston, E.; Callegan, M.C. Targets of immunomodulation in bacterial endophthalmitis. Prog. Retin. Eye Res. 2019. [Google Scholar] [CrossRef]
- Talreja, D.; Kaye, K.S.; Yu, F.S.; Walia, S.K.; Kumar, A. Pathogenicity of ocular isolates of Acinetobacter baumannii in a mouse model of bacterial endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2014, 55, 2392–2402. [Google Scholar] [CrossRef][Green Version]
- Parkunan, S.M.; Randall, C.B.; Coburn, P.S.; Astley, R.A.; Staats, R.L.; Callegan, M.C. Unexpected Roles for Toll-Like Receptor 4 and TRIF in Intraocular Infection with Gram-Positive Bacteria. Infect. Immun. 2015, 83, 3926–3936. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.J.; Astley, R.; Wheatley, N.; Wang, J.-T.; Callegan, M.C. TLR4 Contributes to the Host Response to Klebsiella Intraocular Infection. Curr. Eye Res. 2014, 39, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Whiston, E.A.; Sugi, N.; Kamradt, M.C.; Sack, C.; Heimer, S.R.; Engelbert, M.; Wawrousek, E.F.; Gilmore, M.S.; Ksander, B.R.; Gregory, M.S. alphaB-crystallin protects retinal tissue during Staphylococcus aureus-induced endophthalmitis. Infect. Immun. 2008, 76, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Engelbert, M.; Gilmore, M.S. Fas ligand but not complement is critical for control of experimental Staphylococcus aureus Endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2479–2486. [Google Scholar] [CrossRef]
- Kochan, T.; Singla, A.; Tosi, J.; Kumar, A. Toll-Like Receptor 2 Ligand Pretreatment Attenuates Retinal Microglial Inflammatory Response but Enhances Phagocytic Activity toward Staphylococcus aureus. Infect. Immun. 2012, 80, 2076–2088. [Google Scholar] [CrossRef]
- Giese, M.J.; Sumner, H.L.; Berliner, J.A.; Mondino, B.J. Cytokine expression in a rat model of Staphylococcus aureus endophthalmitis. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2785–2790. [Google Scholar]
- Nakazawa, T.; Kayama, M.; Ryu, M.; Kunikata, H.; Watanabe, R.; Yasuda, M.; Kinugawa, J.; Vavvas, D.; Miller, J.W. Tumor Necrosis Factor-α Mediates Photoreceptor Death in a Rodent Model of Retinal Detachment. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1384–1391. [Google Scholar] [CrossRef]
- Ramadan, R.T.; Moyer, A.L.; Callegan, M.C. A role for tumor necrosis factor-alpha in experimental Bacillus cereus endophthalmitis pathogenesis. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4482–4489. [Google Scholar] [CrossRef][Green Version]
- Pires, S.; Parker, D. IL-1β activation in response to Staphylococcus aureus lung infection requires inflammasome-dependent and independent mechanisms. Eur. J. Immunol. 2018, 48, 1707–1716. [Google Scholar] [CrossRef]
- Miller, L.S.; Pietras, E.M.; Uricchio, L.H.; Hirano, K.; Rao, S.; Lin, H.; O’Connell, R.M.; Iwakura, Y.; Cheung, A.L.; Cheng, G.; et al. Inflammasome-Mediated Production of IL-1β Is Required for Neutrophil Recruitment against Staphylococcus aureus In Vivo. J. Immunol. 2007, 179, 6933–6942. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio Rerio), 4th ed.; University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, F.; Rolain, M.; Zhou, X.Y.; Singh, P.K.; Thummel, R.; Kumar, A. Zebrafish are Resistant to Staphylococcus aureus Endophthalmitis. Pathogens 2019, 8, 207. https://doi.org/10.3390/pathogens8040207
Mei F, Rolain M, Zhou XY, Singh PK, Thummel R, Kumar A. Zebrafish are Resistant to Staphylococcus aureus Endophthalmitis. Pathogens. 2019; 8(4):207. https://doi.org/10.3390/pathogens8040207
Chicago/Turabian StyleMei, Frank, Matthew Rolain, Xiao Yi Zhou, Pawan Kumar Singh, Ryan Thummel, and Ashok Kumar. 2019. "Zebrafish are Resistant to Staphylococcus aureus Endophthalmitis" Pathogens 8, no. 4: 207. https://doi.org/10.3390/pathogens8040207
APA StyleMei, F., Rolain, M., Zhou, X. Y., Singh, P. K., Thummel, R., & Kumar, A. (2019). Zebrafish are Resistant to Staphylococcus aureus Endophthalmitis. Pathogens, 8(4), 207. https://doi.org/10.3390/pathogens8040207