In Vitro Benznidazole and Nifurtimox Susceptibility Profile of Trypanosoma cruzi Strains Belonging to Discrete Typing Units TcI, TcII, and TcV



Abstract
1. Introduction
2. Results
2.1. Genetic Diversity of T. cruzi Strains under Studies
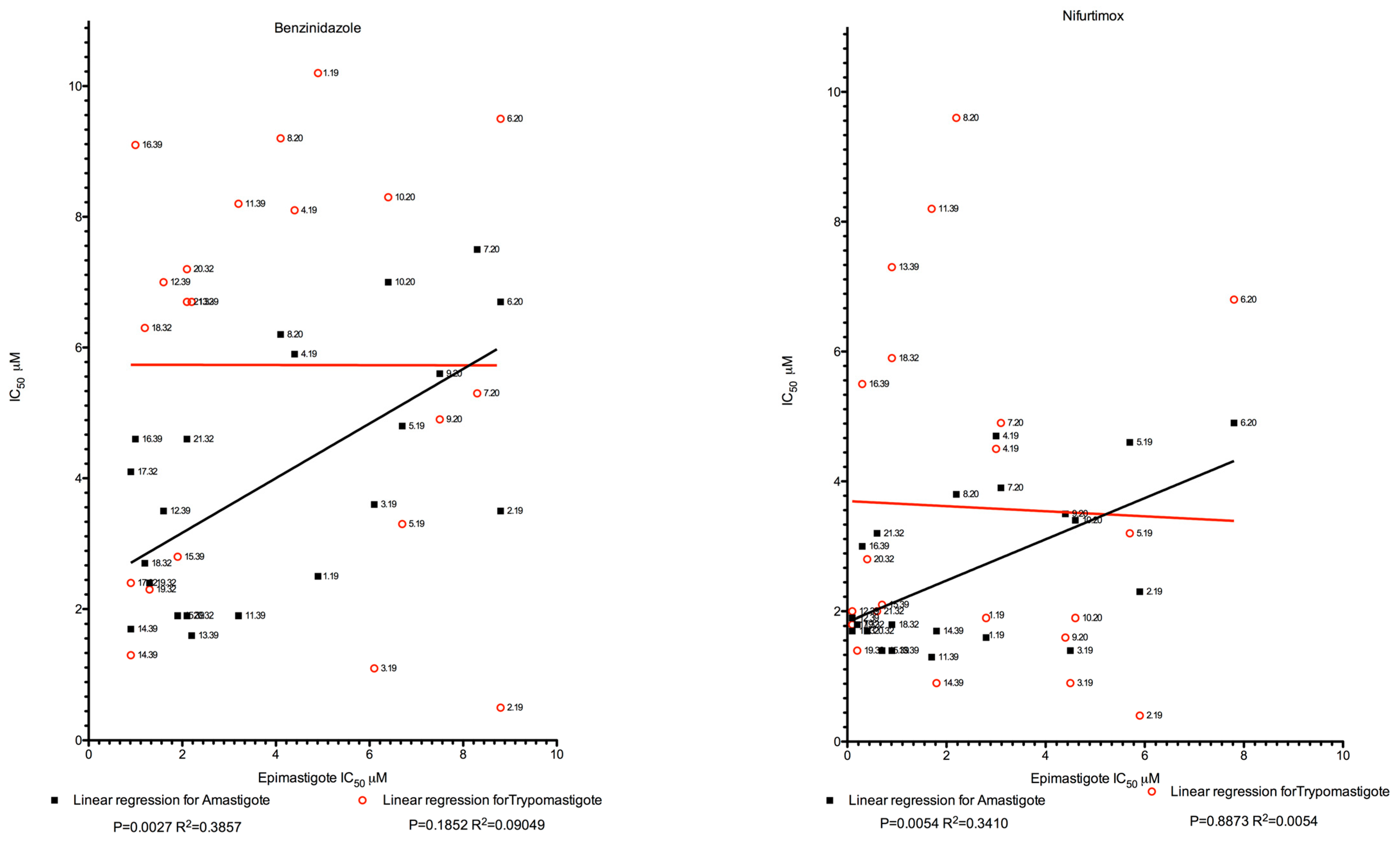
2.2. Epimastigote, Trypomastigote, and Amastigote Susceptibility Towards NFX and BZN
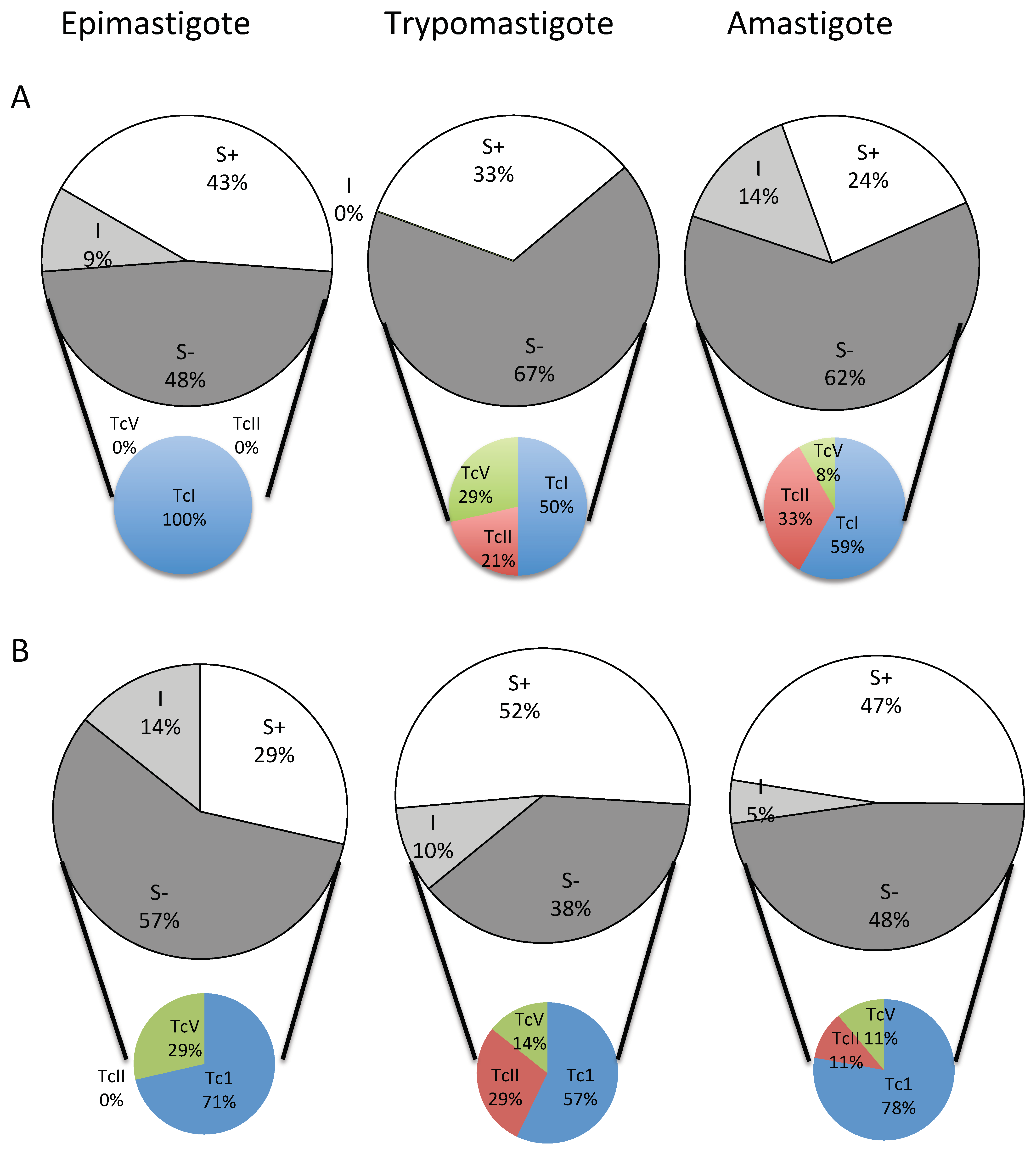
2.3. Frequency of BZN and NFX S− (Not Wild-Type) Phenotypes
3. Discussion
4. Materials and Methods
4.1. Parasites and Strains
4.2. Parasite and Host Cell Cultivation
4.3. Drugs
4.4. In Vitro Tests on Epimastigotes
4.5. In Vitro Tests on Trypomastigotes
4.6. In Vitro Tests on Intracellular Amastigotes
4.7. Cutoff Determination and Drug Sensitivity Correlation at Distinct Parasite Stages
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bern, C. Chagas’ Disease. N. Engl. J. Med. 2015, 373, 1882. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.R.L.; Nascimento, R.J.; Sturm, N.R. Evolution and pathology in chagas disease—A review. Mem. Inst. Oswaldo Cruz 2006, 101, 463–491. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.R.L.; Hecht, M.M.; Guimaro, M.C.; Sousa, A.O.; Nitz, N. Pathogenesis of chagas’ disease: Parasite persistence and autoimmunity. Clin. Microbiol. Rev. 2011, 24, 592–630. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.R.L.; Gomes, C.; Nitz, N.; Sousa, A.O.; Alves, R.M.; Guimaro, M.C.; Cordeiro, C.; Bernal, F.M.; Rosa, A.C.; Hejnar, J.; et al. Trypanosoma cruzi in the chicken model: Chagas-like heart disease in the absence of parasitism. PLoS Negl. Trop. Dis. 2011, 5, e1000. [Google Scholar] [CrossRef]
- Pérez-Molina, J.A.; Norman, F.; López-Vélez, R. Chagas disease in non-endemic countries: Epidemiology, clinical presentation and treatment. Curr. Infect. Dis. Rep. 2012, 14, 263–274. [Google Scholar] [CrossRef]
- Meymandi, S.; Hernandez, S.; Park, S.; Sanchez, D.R.; Forsyth, C. Treatment of Chagas Disease in the United States. Curr. Treat. Options Infect. Dis. 2018, 10, 373–388. [Google Scholar] [CrossRef]
- Gajdács, M.; Albericio, F. Antibiotic Resistance: From the Bench to Patients. Antibiotics 2019, 8, 129. [Google Scholar] [CrossRef]
- Gajdács, M. The Concept of an Ideal Antibiotic: Implications for Drug Design. Molecules 2019, 24, 892. [Google Scholar] [CrossRef]
- Molina, I.; Gómez i Prat, J.; Salvador, F.; Treviño, B.; Sulleiro, E.; Serre, N.; Pou, D.; Roure, S.; Cabezos, J.; Valerio, L.; et al. Randomized trial of posaconazole and benznidazole for chronic Chagas’ disease. N. Engl. J. Med. 2014, 370, 1899–1908. [Google Scholar] [CrossRef]
- Morillo, C.A.; Marin-Neto, J.A.; Avezum, A.; Sosa-Estani, S.; Rassi, A.; Rosas, F.; Villena, E.; Quiroz, R.; Bonilla, R.; Britto, C.; et al. Randomized Trial of Benznidazole for Chronic Chagas’ Cardiomyopathy. N. Engl. J. Med. 2015, 373, 1295–1306. [Google Scholar] [CrossRef]
- Zingales, B.; Andrade, S.G.; Briones, M.R.S.; Campbell, D.A.; Chiari, E.; Fernandes, O.; Guhl, F.; Lages-Silva, E.; Macedo, A.M.; Machado, C.R.; et al. A new consensus for Trypanosoma cruzi intraspecific nomenclature: Second revision meeting recommends TcI to TcVI. Mem. Inst. Oswaldo Cruz 2009, 104, 1051–1054. [Google Scholar] [CrossRef] [PubMed]
- Brisse, S.; Barnabé, C.; Tibayrenc, M. Identification of six Trypanosoma cruzi phylogenetic lineages by random amplified polymorphic DNA and multilocus enzyme electrophoresis. Int. J. Parasitol. 2000, 30, 35–44. [Google Scholar] [CrossRef]
- Pinto, C.M.; Kalko, E.K.V.; Cottontail, I.; Wellinghausen, N.; Cottontail, V.M. TcBat a bat-exclusive lineage of Trypanosoma cruzi in the Panama Canal Zone, with comments on its classification and the use of the 18S rRNA gene for lineage identification. Infect. Genet. Evol. 2012, 12, 1328–1332. [Google Scholar] [CrossRef]
- Zingales, B.; Miles, M.A.; Campbell, D.A.; Tibayrenc, M.; Macedo, A.M.; Teixeira, M.M.G.; Schijman, A.G.; Llewellyn, M.S.; Lages-Silva, E.; Machado, C.R.; et al. The revised Trypanosoma cruzi subspecific nomenclature: Rationale, epidemiological relevance and research applications. Infect. Genet. Evol. 2012, 12, 240–253. [Google Scholar] [CrossRef]
- Marcili, A.; Lima, L.; Cavazzana, M.; Junqueira, A.C.V.; Veludo, H.H.; Maia Da Silva, F.; Campaner, M.; Paiva, F.; Nunes, V.L.B.; Teixeira, M.M.G. A new genotype of Trypanosoma cruzi associated with bats evidenced by phylogenetic analyses using SSU rDNA, cytochrome b and Histone H2B genes and genotyping based on ITS1 rDNA. Parasitology 2009, 136, 641–655. [Google Scholar] [CrossRef] [PubMed]
- Tibayrenc, M.; Ward, P.; Moya, A.; Ayala, F.J. Natural populations of Trypanosoma cruzi, the agent of Chagas disease, have a complex multiclonal structure. Proc. Natl. Acad. Sci. USA 1986, 83, 115–119. [Google Scholar] [CrossRef]
- Neal, R.A.; van Bueren, J. Comparative studies of drug susceptibility of five strains of Trypanosoma cruzi in vivo and in vitro. Trans. R. Soc. Trop. Med. Hyg. 1988, 82, 709–714. [Google Scholar] [CrossRef]
- Canavaci, A.M.C.; Bustamante, J.M.; Padilla, A.M.; Perez Brandan, C.M.; Simpson, L.J.; Xu, D.; Boehlke, C.L.; Tarleton, R.L. In vitro and in vivo high-throughput assays for the testing of anti-Trypanosoma cruzi compounds. PLoS Negl. Trop. Dis. 2010, 4, e740. [Google Scholar] [CrossRef]
- Filardi, L.S.; Brener, Z. Susceptibility and natural resistance of Trypanosoma cruzi strains to drugs used clinically in Chagas disease. Trans. R. Soc. Trop. Med. Hyg. 1987, 81, 755–759. [Google Scholar] [CrossRef]
- Molina, J.; Martins-Filho, O.; Brener, Z.; Romanha, A.J.; Loebenberg, D.; Urbina, J.A. Activities of the triazole derivative SCH 56592 (posaconazole) against drug-resistant strains of the protozoan parasite Trypanosoma (Schizotrypanum) cruzi in immunocompetent and immunosuppressed murine hosts. Antimicrob. Agents Chemother. 2000, 44, 150–155. [Google Scholar] [CrossRef]
- Toledo, M.J.; Bahia, M.T.; Carneiro, C.M.; Martins-Filho, O.A.; Tibayrenc, M.; Barnabé, C.; Tafuri, W.L.; de Lana, M. Chemotherapy with benznidazole and itraconazole for mice infected with different Trypanosoma cruzi clonal genotypes. Antimicrob. Agents Chemother. 2003, 47, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Revollo, S.; Oury, B.; Laurent, J.P.; Barnabé, C.; Quesney, V.; Carrière, V.; Noël, S.; Tibayrenc, M. Trypanosoma cruzi: Impact of clonal evolution of the parasite on its biological and medical properties. Exp. Parasitol. 1998, 89, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, D.; Barnabé, C.; Sereno, D.; Tibayrenc, M. Lack of correlation between in vitro susceptibility to Benznidazole and phylogenetic diversity of Trypanosoma cruzi, the agent of Chagas disease. Exp. Parasitol. 2004, 108, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Kahlmeter, G. Defining antibiotic resistance-towards international harmonization. Upsala J. Med. Sci. 2014, 119, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Sereno, D.; Harrat, Z.; Eddaikra, N. Meta-analysis and discussion on challenges to translate Leishmania drug resistance phenotyping into the clinic. Acta Trop. 2019, 191, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Eddaikra, N.; Ait-Oudhia, K.; Kherrachi, I.; Oury, B.; Moulti-Mati, F.; Benikhlef, R.; Harrat, Z.; Sereno, D. Antimony susceptibility of Leishmania isolates collected over a 30-year period in Algeria. PLoS Negl. Trop. Dis. 2018, 12, e0006310. [Google Scholar] [CrossRef]
- Brenière, S.F.; Waleckx, E.; Barnabé, C. Over Six Thousand Trypanosoma cruzi Strains Classified into Discrete Typing Units (DTUs): Attempt at an Inventory. PLoS Negl. Trop. Dis. 2016, 10, e0004792. [Google Scholar] [CrossRef]
- Wilkinson, S.R.; Taylor, M.C.; Horn, D.; Kelly, J.M.; Cheeseman, I. A mechanism for cross-resistance to nifurtimox and benznidazole in trypanosomes. Proc. Natl. Acad. Sci. USA 2008, 105, 5022–5027. [Google Scholar] [CrossRef]
- Wyllie, S.; Foth, B.J.; Kelner, A.; Sokolova, A.Y.; Berriman, M.; Fairlamb, A.H. Nitroheterocyclic drug resistance mechanisms in Trypanosoma brucei. J. Antimicrob. Chemother. 2016, 71, 625–634. [Google Scholar] [CrossRef]
- Moraes, C.B.; Giardini, M.A.; Kim, H.; Franco, C.H.; Araujo-Junior, A.M.; Schenkman, S.; Chatelain, E.; Freitas-Junior, L.H. Nitroheterocyclic compounds are more efficacious than CYP51 inhibitors against Trypanosoma cruzi: Implications for Chagas disease drug discovery and development. Sci. Rep. 2014, 4, 4703. [Google Scholar] [CrossRef]
- MacLean, L.M.; Thomas, J.; Lewis, M.D.; Cotillo, I.; Gray, D.W.; De Rycker, M. Development of Trypanosoma cruzi in vitro assays to identify compounds suitable for progression in Chagas’ disease drug discovery. PLoS Negl. Trop. Dis. 2018, 12, e0006612. [Google Scholar] [CrossRef] [PubMed]
- Fabbro, D.L.; Streiger, M.L.; Arias, E.D.; Bizai, M.L.; del Barco, M.; Amicone, N.A. Trypanocide treatment among adults with chronic Chagas disease living in Santa Fe city (Argentina), over a mean follow-up of 21 years: Parasitological, serological and clinical evolution. Rev. Soc. Bras. Med. Trop. 2007, 40, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cançado, J.R. Long term evaluation of etiological treatment of Chagas disease with benznidazole. Rev. Inst. Med. Trop. Sao Paulo 2002, 44, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Valdéz, F.J.; Padilla, A.; Wang, W.; Orr, D.; Tarleton, R.L. Spontaneous dormancy protects Trypanosoma cruzi during extended drug exposure. Elife 2018, 7, e34039. [Google Scholar] [CrossRef] [PubMed]
- Huber, W.; Koella, J.C. A comparison of three methods of estimating EC50 in studies of drug resistance of malaria parasites. Acta Trop. 1993, 55, 257–261. [Google Scholar] [CrossRef]
- Smith, P.; Endris, R.; Kronvall, G.; Thomas, V.; Verner-Jeffreys, D.; Wilhelm, C.; Dalsgaard, I. Epidemiological cut-off values for Flavobacterium psychrophilum MIC data generated by a standard test protocol. J. Fish Dis. 2016, 39, 143–154. [Google Scholar] [CrossRef]
- Smith, P.; Kronvall, G. How many strains are required to set an epidemiological cut-off value for MIC values determined for bacteria isolated from aquatic animals? Aquac. Int. 2015, 23, 465–470. [Google Scholar] [CrossRef]
- Budczies, J.; Klauschen, F.; Sinn, B.V.; Győrffy, B.; Schmitt, W.D.; Darb-Esfahani, S.; Denkert, C. Cutoff Finder: A Comprehensive and Straightforward Web Application Enabling Rapid Biomarker Cutoff Optimization. PLoS ONE 2012, 7, e51862. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strain Identification | Ref | DTU | Country | Host |
|---|---|---|---|---|
| SP104 cl1 | 1/19 | TcI | Chile | Mepraia spinolai |
| CUTIA cl1 | 2/19 | TcI | Brazil | Dasiprocta aguti |
| Gamba cl1 | 3/19 | TcI | Brazil | Didelphis azarae |
| 13379 cl7 | 4/19 | TcI | Bolivia | Homo sapiens |
| OPS21 cl11 | 5/19 | TcI | Venezuela | Homo sapiens |
| SO34 cl4 | 6/20 | TcI | Bolivia | Triatoma infestans |
| CUICA cl1 | 7/20 | TcI | Brazil | Philander opossum |
| P209 cl1 | 8/20 | TcI | Bolivia | Homo sapiens |
| Esquilo cl1 | 9/20 | TcI | Brazil | Sciurus aestuans ingrami |
| P11 cl3 | 10/20 | TcI | Bolivia | Homo sapiens |
| SC43 cl1 | 11/39 | TcV | Bolivia | Triatoma infestans |
| Bug2148 cl1 | 12/39 | TcV | Brazil | Triatoma infestans |
| Bug2149 cl10 | 13/39 | TcV | Brazil | Triatoma infestans |
| SO3 cl5 | 14/39 | TcV | Bolivia | Triatoma infestans |
| MN cl2 | 15/39 | TcV | Chile | Homo sapiens |
| NR cl3 | 16/39 | TcV | Chile | Homo sapiens |
| MAS1 cl1 | 17/32 | TcII | Brazil | Homo sapiens |
| CBB cl1 | 18/32 | TcII | Chile | Homo sapiens |
| Tu18 cl2 | 19/32 | TcII | Bolivia | Triatoma infestans |
| IVV cl4 | 20/32 | TcII | Chile | Homo sapiens |
| MVB cl8 | 21/32 | TcII | Chile | Homo sapiens |
| Ref | Epi (µM) | Status COwt = 2.68 S+<2.15>I<3.22>S− | Trypo (µM) | Status COwt = 4.01 S+<3.21>I<4.82>S− | Ama (µM) | Status COwt =2.67 S+<2.14>I<3.21>S− |
|---|---|---|---|---|---|---|
| 1/19 | 4.90 | S− | 10.20 | S− | 2.5 | I |
| 2/19 | 8.80 | S− | 0.50 | S+ | 3.5 | S− |
| 3/19 | 6.10 | S− | 1.10 | S+ | 3.6 | S− |
| 4/19 | 4.40 | S− | 8.10 | S− | 5.9 | S− |
| 5/19 | 6.70 | S− | 3.30 | S+ | 4.8 | S− |
| 6/20 | 8.80 | S− | 9.50 | S− | 6.7 | S− |
| 7/20 | 8.30 | S− | 5.30 | S− | 7.5 | S− |
| 8/20 | 4.10 | S− | 9.20 | S− | 6.2 | S− |
| 9/20 | 7.50 | S− | 4.90 | S− | 5.6 | S− |
| 10/20 | 6.40 | S− | 8.30 | S− | 7.0 | S− |
| TcI mean | 6.60 ± 1.75 | 6.04 ± 3.54 | 5.33 ± 1.67 | |||
| 11/39 | 3.20 | I | 8.20 | S− | 1.9 | S+ |
| 12/39 | 1.60 | S+ | 7.00 | S− | 3.5 | S− |
| 13/39 | 2.20 | I | 6.70 | S− | 1.6 | S+ |
| 14/39 | 0.90 | S+ | 1.30 | S+ | 1.7 | S+ |
| 15/39 | 1.90 | S+ | 2.80 | S+ | 1.9 | S+ |
| 16/39 | 1.00 | S+ | 9.10 | S− | 4.6 | S− |
| TcV mean | 1.81 ± 0.85 | 5.85 ± 3.10 | 2.53 ± 1.23 | |||
| 17/32 | 0.90 | S+ | 2.40 | S+ | 4.1 | S− |
| 18/32 | 1.20 | S+ | 6.30 | S− | 2.7 | I |
| 19/32 | 1.30 | S+ | 2.30 | S+ | 2.4 | I |
| 20/32 | 2.10 | S+ | 7.20 | S− | 1.9 | S+ |
| 21/32 | 2.10 | S+ | 6.70 | S− | 4.6 | S− |
| TcII mean | 1.52 ± 0.54 | 4.98 ± 2.42 | 3.14 ± 1.15 | |||
| Total mean | 4.02 ± 2.82 | 5.73 ± 3.07 | 4.00 ± 1.90 |
| Ref | Epi (µM) | Status COwt= 1.089 S+<0.87>I<1.30 >S− | Trypo (µM) | Status COwt = 3.24 S+<2.59>I<3.88>S− | Ama (µM) | Status COwt =2.67 S+<1.70>I<2.54>S− |
|---|---|---|---|---|---|---|
| 1/19 | 2.80 | S− | 1.90 | S+ | 1.60 | S+ |
| 2/19 | 5.90 | S− | 0.40 | S+ | 2.30 | I |
| 3/19 | 4.50 | S− | 0.90 | S+ | 1.40 | S+ |
| 4/19 | 3.00 | S− | 4.50 | S− | 4.70 | S− |
| 5/19 | 5.70 | S− | 3.20 | I | 4.60 | S− |
| 6/20 | 7.80 | S− | 6.80 | S− | 4.90 | S− |
| 7/20 | 3.10 | S− | 4.90 | S− | 3.90 | S− |
| 8/20 | 2.20 | S− | 9.60 | S− | 3.80 | S− |
| 9/20 | 4.40 | S− | 1.60 | S+ | 3.50 | S− |
| 10/20 | 4.60 | S− | 1.90 | S+ | 3.40 | S− |
| TcI mean | 4.40 ± 1.71 | 3.57 ± 2.91 | 3.41 ± 1.25 | |||
| 11/39 | 1.70 | S− | 8.20 | S− | 1.30 | S+ |
| 12/39 | 0.10 | S+ | 2.00 | S+ | 1.90 | I |
| 13/39 | 0.90 | I | 7.30 | S− | 1.40 | S+ |
| 14/39 | 1.80 | S− | 0.90 | S+ | 1.70 | S+ |
| 15/39 | 0.70 | S+ | 2.10 | S+ | 1.40 | S+ |
| 16/39 | 0.30 | S+ | 5.50 | S− | 3.00 | S− |
| TcV mean | 0.91 ± 0.70 | 4.33 ± 3.07 | 1.78 ± 0.63 | |||
| 17/32 | 0.10 | S+ | 1.80 | S+ | 1.70 | S+ |
| 18/32 | 0.90 | I | 5.90 | S− | 1.80 | I |
| 19/32 | 0.20 | S+ | 1.40 | S+ | 1.80 | I |
| 20/32 | 0.40 | S+ | 2.80 | I | 1.70 | S+ |
| 21/32 | 0.60 | S+ | 2.00 | S+ | 3.20 | S− |
| TcII mean | 0.44 ± 0.32 | 2.78 ± 1.81 | 2.04 ± 0.65 | |||
| Total mean | 2.46 ± 2.25 | 3.60 ± 2.67 | 2.61 ± 1.22 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Revollo, S.; Oury, B.; Vela, A.; Tibayrenc, M.; Sereno, D. In Vitro Benznidazole and Nifurtimox Susceptibility Profile of Trypanosoma cruzi Strains Belonging to Discrete Typing Units TcI, TcII, and TcV. Pathogens 2019, 8, 197. https://doi.org/10.3390/pathogens8040197
Revollo S, Oury B, Vela A, Tibayrenc M, Sereno D. In Vitro Benznidazole and Nifurtimox Susceptibility Profile of Trypanosoma cruzi Strains Belonging to Discrete Typing Units TcI, TcII, and TcV. Pathogens. 2019; 8(4):197. https://doi.org/10.3390/pathogens8040197
Chicago/Turabian StyleRevollo, Susana, Bruno Oury, Andrea Vela, Michel Tibayrenc, and Denis Sereno. 2019. "In Vitro Benznidazole and Nifurtimox Susceptibility Profile of Trypanosoma cruzi Strains Belonging to Discrete Typing Units TcI, TcII, and TcV" Pathogens 8, no. 4: 197. https://doi.org/10.3390/pathogens8040197
APA StyleRevollo, S., Oury, B., Vela, A., Tibayrenc, M., & Sereno, D. (2019). In Vitro Benznidazole and Nifurtimox Susceptibility Profile of Trypanosoma cruzi Strains Belonging to Discrete Typing Units TcI, TcII, and TcV. Pathogens, 8(4), 197. https://doi.org/10.3390/pathogens8040197