Inactivation of Zika Virus by Photoactive Iodonaphthyl Azide Preserves Immunogenic Potential of the Virus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. ZIKV Is Inactivated by INA
2.2. RNA Isolated from INA-iZIKV Is Non-infectious
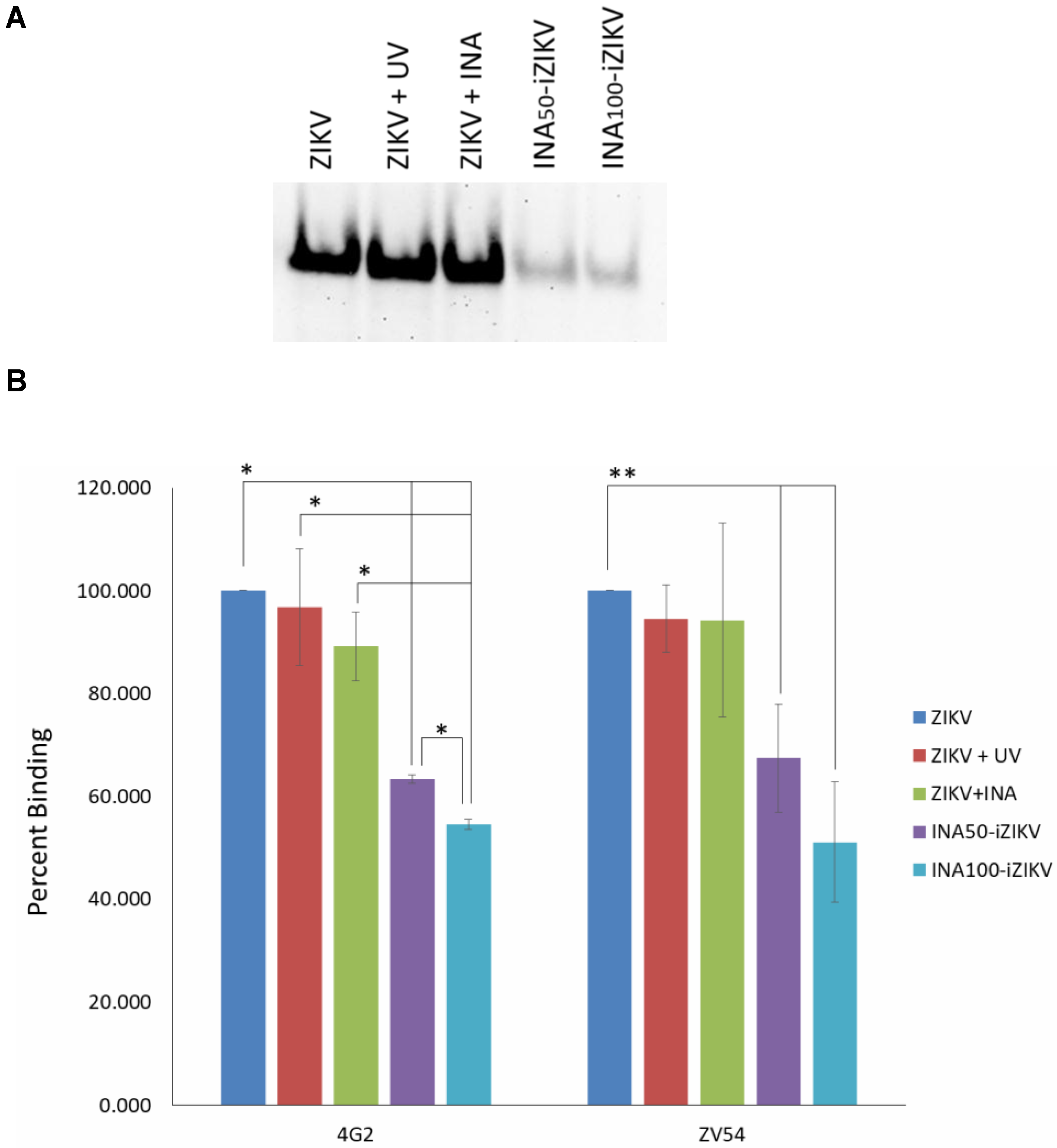
2.3. INA-iZIKV Binds to Anti-ZIKV Monoclonal Antibodies
2.4. Immunization with INA-iZIKV Induces Anti-ZIKV Antibody Response
3. Discussion
4. Methods
4.1. Virus and Inactivation
4.2. Serial Passaging of Inactivated Virus In-Vitro
4.3. Immunofluorescence
4.4. Western Blot Analysis of INA-iZIKV
4.5. ELISA to Determine Binding of Anti-ZIKV Antibodies to Native form of INA-iZIKV
4.6. RNA Isolation and Transfection
4.7. Immunization of Mice with INA-iZIKV
4.8. Total Anti-ZIKV IgG Titer
4.9. ZIKV-neutralizing Antibody Titer
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kindhauser, M.K.; Allen, T.; Frank, V.; Santhana, R.S.; Dye, C. Zika: The origin and spread of a mosquito-borne virus. Bull. World Health Organ. 2016, 94, 675. [Google Scholar] [CrossRef] [PubMed]
- Ladhani, S.N.; O’Connor, C.; Kirkbride, H.; Brooks, T.; Morgan, D. Outbreak of Zika virus disease in the Americas and the association with microcephaly, congenital malformations and Guillain-Barre syndrome. Arch. Dis. Child. 2016, 101, 600–602. [Google Scholar] [CrossRef][Green Version]
- Satterfield-Nash, A.; Kotzky, K.; Allen, J.; Bertolli, J.; Moore, C.A.; Pereira, I.O.; Pessoa, A.; Melo, F.; Santelli, A.; Boyle, C.A.; et al. Health and Development at Age 19–24 Months of 19 Children Who Were Born with Microcephaly and Laboratory Evidence of Congenital Zika Virus Infection During the 2015 Zika Virus Outbreak - Brazil, 2017. Morb. Mortal. Wkly. Rep. 2017, 66, 1347–1351. [Google Scholar] [CrossRef]
- Siedner, M.J.; Ryan, E.T.; Bogoch, I.I. Gone or forgotten? The rise and fall of Zika virus. Lancet Public Health 2018, 3, e109–e110. [Google Scholar] [CrossRef]
- Yadav, P.D.; Malhotra, B.; Sapkal, G.; Nyayanit, D.A.; Deshpande, G.; Gupta, N.; Padinjaremattathil, U.T.; Sharma, H.; Sahay, R.R.; Sharma, P.; et al. Zika virus outbreak in Rajasthan, India in 2018 was caused by a virus endemic to Asia. Infect. Genet. Evol. 2019, 69, 199–202. [Google Scholar] [CrossRef]
- Gardner, L.M.; Bota, A.; Gangavarapu, K.; Kraemer, M.U.G.; Grubaugh, N.D. Inferring the risk factors behind the geographical spread and transmission of Zika in the Americas. PLoS Negl. Trop. Dis. 2018, 12, e0006194. [Google Scholar] [CrossRef]
- De Oliveira Garcia, M.H. Zika: The continuing threat. Bull. World Health Organ. 2019, 97, 6–7. [Google Scholar] [CrossRef]
- Nurnberger, C.; Bodmer, B.S.; Fiedler, A.H.; Gabriel, G.; Muhlebach, M.D. A Measles Virus-Based Vaccine Candidate Mediates Protection against Zika Virus in an Allogeneic Mouse Pregnancy Model. J. Virol. 2019, 93, e01485–e01518. [Google Scholar] [CrossRef] [PubMed]
- Modjarrad, K.; Lin, L.; George, S.L.; Stephenson, K.E.; Eckels, K.H.; De La Barrera, R.A.; Jarman, R.G.; Sondergaard, E.; Tennant, J.; Ansel, J.L.; et al. Preliminary aggregate safety and immunogenicity results from three trials of a purified inactivated Zika virus vaccine candidate: Phase 1, randomised, double-blind, placebo-controlled clinical trials. Lancet 2018, 391, 563–571. [Google Scholar] [CrossRef]
- Medina, L.O.; To, A.; Lieberman, M.M.; Wong, T.A.S.; Namekar, M.; Nakano, E.; Andersen, H.; Yalley-Ogunro, J.; Greenhouse, J.; Higgs, S.; et al. A Recombinant Subunit Based Zika Virus Vaccine Is Efficacious in Non-human Primates. Front. Immunol. 2018, 9, 2464. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.D.T. Current status of Zika vaccine development: Zika vaccines advance into clinical evaluation. NPJ Vaccines 2018, 3, 24. [Google Scholar] [CrossRef] [PubMed]
- Tebas, P.; Roberts, C.C.; Muthumani, K.; Reuschel, E.L.; Kudchodkar, S.B.; Zaidi, F.I.; White, S.; Khan, A.S.; Racine, T.; Choi, H.; et al. Safety and Immunogenicity of an Anti-Zika Virus DNA Vaccine—Preliminary Report. N. Engl. J. Med. 2017. [Google Scholar] [CrossRef] [PubMed]
- Bercovici, T.; Gitler, C. 5-[125I]Iodonaphthyl azide, a reagent to determine the penetration of proteins into the lipid bilayer of biological membranes. Biochemistry 1978, 17, 1484–1489. [Google Scholar] [CrossRef] [PubMed]
- Raviv, Y.; Bercovici, T.; Gitler, C.; Salomon, Y. Detection of nearest neighbors to specific fluorescently tagged ligands in rod outer segment and lymphocyte plasma membranes by photosensitization of 5-iodonaphthyl 1-azide. Biochemistry 1989, 28, 1313–1319. [Google Scholar] [CrossRef]
- Raviv, Y.; Bercovici, T.; Gitler, C.; Salomon, Y. Selective photoinduced uncoupling of the response of adenylate cyclase to gonadotropins by 5-iodonaphthyl 1-azide. Biochemistry 1984, 23, 503–508. [Google Scholar] [CrossRef]
- Raviv, Y.; Viard, M.; Bess, J.W., Jr.; Chertova, E.; Blumenthal, R. Inactivation of retroviruses with preservation of structural integrity by targeting the hydrophobic domain of the viral envelope. J. Virol. 2005, 79, 12394–12400. [Google Scholar] [CrossRef]
- Sharma, A.; Gupta, P.; Glass, P.J.; Parker, M.D.; Maheshwari, R.K. Safety and protective efficacy of INA-inactivated Venezuelan equine encephalitis virus: Implication in vaccine development. Vaccine 2011, 29, 953–959. [Google Scholar] [CrossRef][Green Version]
- Sharma, A.; Gupta, P.; Maheshwari, R.K. Inactivation of Chikungunya virus by 1,5 iodonapthyl azide. Virol. J. 2012, 9, 301. [Google Scholar] [CrossRef]
- Raviprakash, K.; Sun, P.; Raviv, Y.; Luke, T.; Martin, N.; Kochel, T. Dengue virus photo-inactivated in presence of 1,5-iodonaphthylazide (INA) or AMT, a psoralen compound (4′-aminomethyl-trioxsalen) is highly immunogenic in mice. Hum. Vaccin. Immunother. 2013, 9, 2336–2341. [Google Scholar] [CrossRef]
- Gupta, P.; Sharma, A.; Spurgers, K.B.; Bakken, R.R.; Eccleston, L.T.; Cohen, J.W.; Honnold, S.P.; Glass, P.J.; Maheshwari, R.K. 1,5-Iodonaphthyl azide-inactivated V3526 protects against aerosol challenge with virulent venezuelan equine encephalitis virus. Vaccine 2016, 34, 2762–2765. [Google Scholar] [CrossRef]
- Honnold, S.P.; Bakken, R.R.; Fisher, D.; Lind, C.M.; Cohen, J.W.; Eccleston, L.T.; Spurgers, K.B.; Maheshwari, R.K.; Glass, P.J. Second generation inactivated eastern equine encephalitis virus vaccine candidates protect mice against a lethal aerosol challenge. PLoS ONE 2014, 9, e104708. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Sharma, A.; Mathias, V.; Raviv, Y.; Blumenthal, R.; Maheshwari, R.K. Inactivation of non-enveloped virus by 1,5 iodonaphthyl azide. BMC Res. Notes 2015, 8, 44. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, S.; Armstrong, N.; Zhao, H.; Hou, W.; Liu, J.; Chen, C.; Wan, J.; Wang, W.; Zhong, C.; Liu, C.; et al. Zika Virus Fatally Infects Wild Type Neonatal Mice and Replicates in Central Nervous System. Viruses 2018, 10, 49. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shi, L.; Karlin, A. A photochemical approach to the lipid accessibility of engineered cysteinyl residues. Proc. Natl. Acad. Sci. USA 2003, 100, 886–891. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Fernandez, E.; Dowd, K.A.; Speer, S.D.; Platt, D.J.; Gorman, M.J.; Govero, J.; Nelson, C.A.; Pierson, T.C.; Diamond, M.S.; et al. Structural Basis of Zika Virus-Specific Antibody Protection. Cell 2016, 166, 1016–1027. [Google Scholar] [CrossRef]
- Larocca, R.A.; Abbink, P.; Peron, J.P.; Zanotto, P.M.; Iampietro, M.J.; Badamchi-Zadeh, A.; Boyd, M.; Kirilova, M.; Nityanandam, R.; Mercado, N.B.; et al. Vaccine protection against Zika virus from Brazil. Nature 2016, 536, 474–478. [Google Scholar] [CrossRef]
- Sharma, A.; Bhomia, M.; Yeh, T.J.; Singh, J.; Maheshwari, R.K. Miltefosine inhibits Chikungunya virus replication in human primary dermal fibroblasts. F1000Research 2018, 7, 9. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Austin, A.L.; Galasso, B.; Nickens, C.; Knollmann-Ritschel, B.; Sharma, A. Inactivation of Zika Virus by Photoactive Iodonaphthyl Azide Preserves Immunogenic Potential of the Virus. Pathogens 2019, 8, 188. https://doi.org/10.3390/pathogens8040188
Austin AL, Galasso B, Nickens C, Knollmann-Ritschel B, Sharma A. Inactivation of Zika Virus by Photoactive Iodonaphthyl Azide Preserves Immunogenic Potential of the Virus. Pathogens. 2019; 8(4):188. https://doi.org/10.3390/pathogens8040188
Chicago/Turabian StyleAustin, Amy L., Bianca Galasso, Caitlin Nickens, Barbara Knollmann-Ritschel, and Anuj Sharma. 2019. "Inactivation of Zika Virus by Photoactive Iodonaphthyl Azide Preserves Immunogenic Potential of the Virus" Pathogens 8, no. 4: 188. https://doi.org/10.3390/pathogens8040188
APA StyleAustin, A. L., Galasso, B., Nickens, C., Knollmann-Ritschel, B., & Sharma, A. (2019). Inactivation of Zika Virus by Photoactive Iodonaphthyl Azide Preserves Immunogenic Potential of the Virus. Pathogens, 8(4), 188. https://doi.org/10.3390/pathogens8040188