An Extract Purified from the Mycelium of a Tomato Wilt-Controlling Strain of Fusarium sambucinum Can Protect Wheat against Fusarium and Common Root Rots

 , and
, and
Abstract
1. Introduction
2. Results
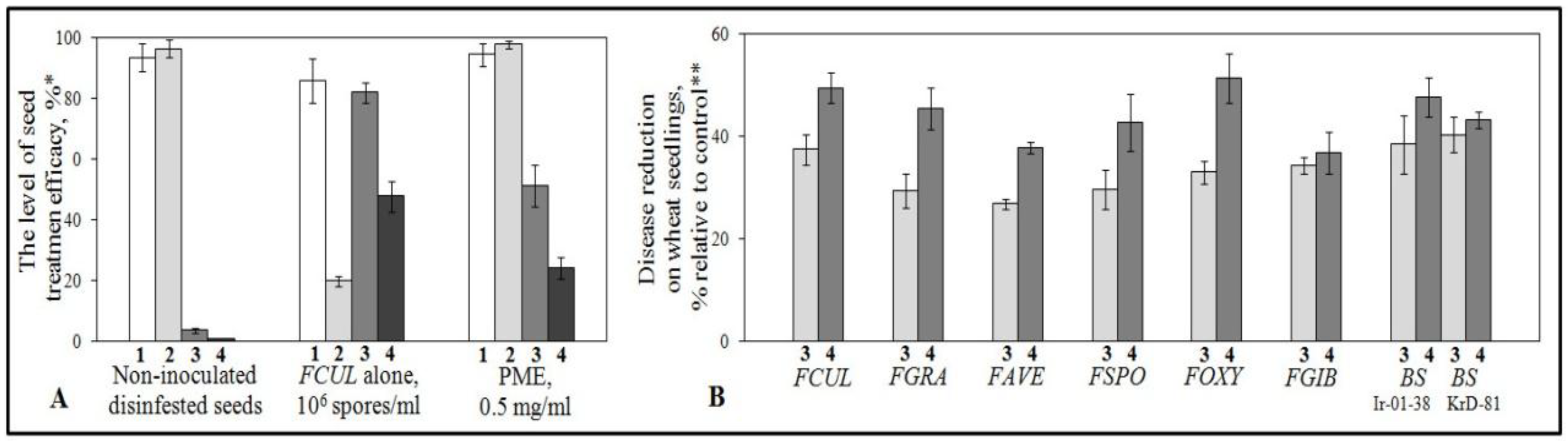
2.1. Protection of Wheat Seedlings with the Purified Mycelial Extract (PME) against Root Rot Pathogens in Vegetation Experiments with Artificial Seed Inoculation
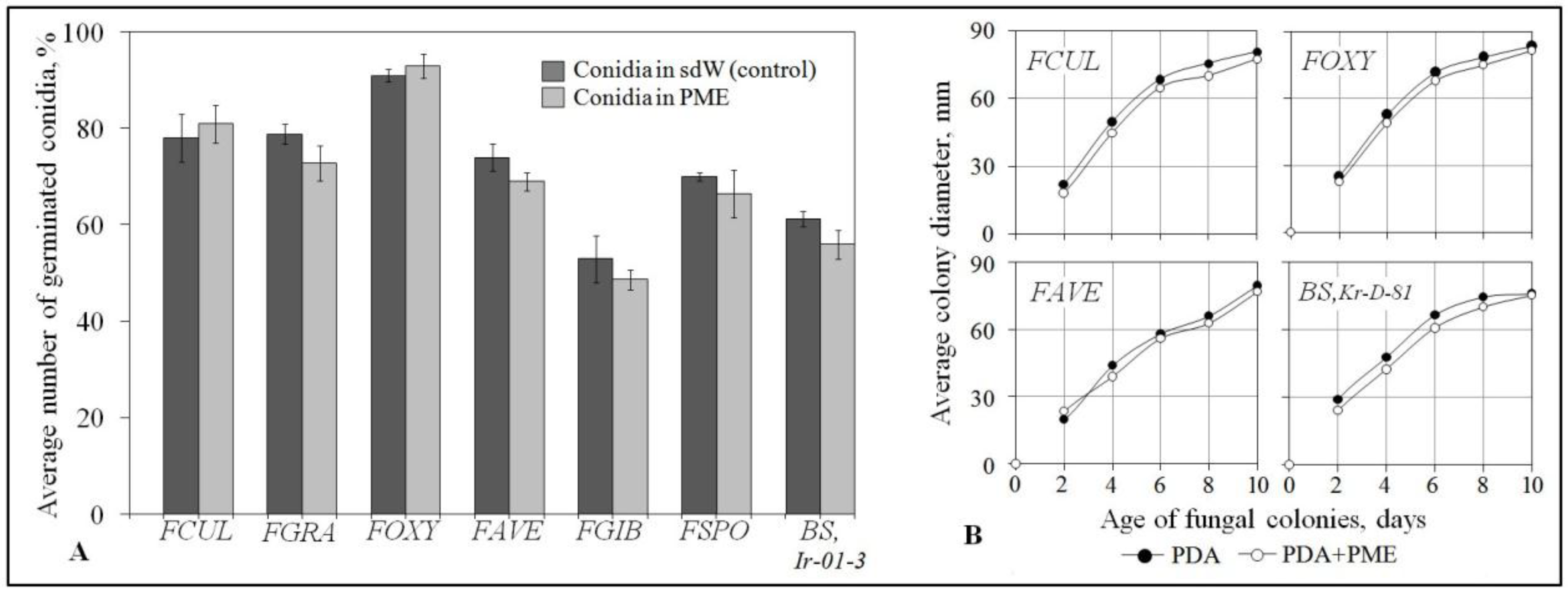
2.2. Plant-Mediated Character of the Protective Activity
2.3. Assessment of PME Influence on Production Some Regulated Fusariotoxins
2.4. Protection Wheat Plants under Field Conditions by Pre-Sowing Treatments of Naturally Infected Seeds with PME
3. Discussion
4. Materials and Methods
4.1. Fungal Cultures
4.2. Purification of Mycelium Extract and Preparation of CL Filtrates
4.3. Seed Treatment
4.4. Artificial Inoculation of Seeds
4.5. Rolled-Towel Assay for Estimation of PME Protection Effect
| 0 - no disease symptoms | |
| 1- weak symptoms (brown streaks or spots) on roots and above | |
| 2 - are manifest, but medium symptoms | |
| 3 - strong extensive symptoms | |
| 4 - crown and root decaying or plant death |
- n1 – the number of seedlings with the same index; d - index according to the rating scale (from 0 to 4);
- n – plant number per treatment.
4.6. Mycological Analysis of Seeds
4.7. In Vitro Testing the Growth and Conidium Germination of Root Rot Agents Exposed to PME
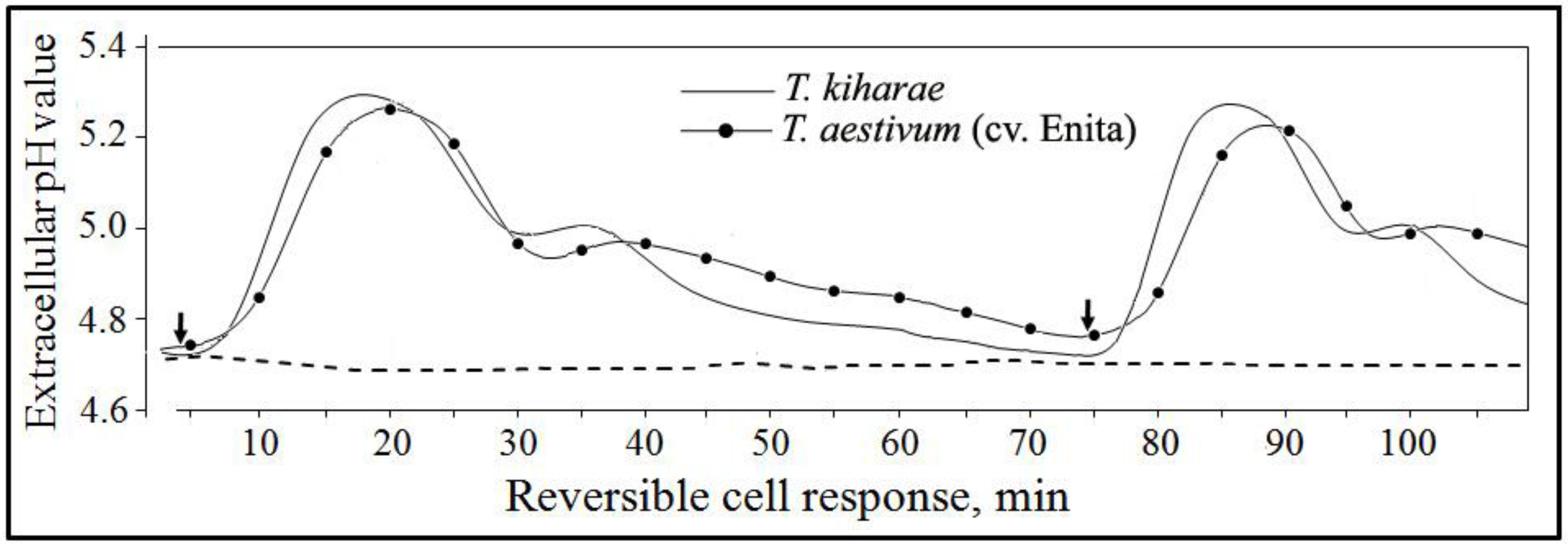
4.8. Studying the PME Effects on Cultured Wheat Cells
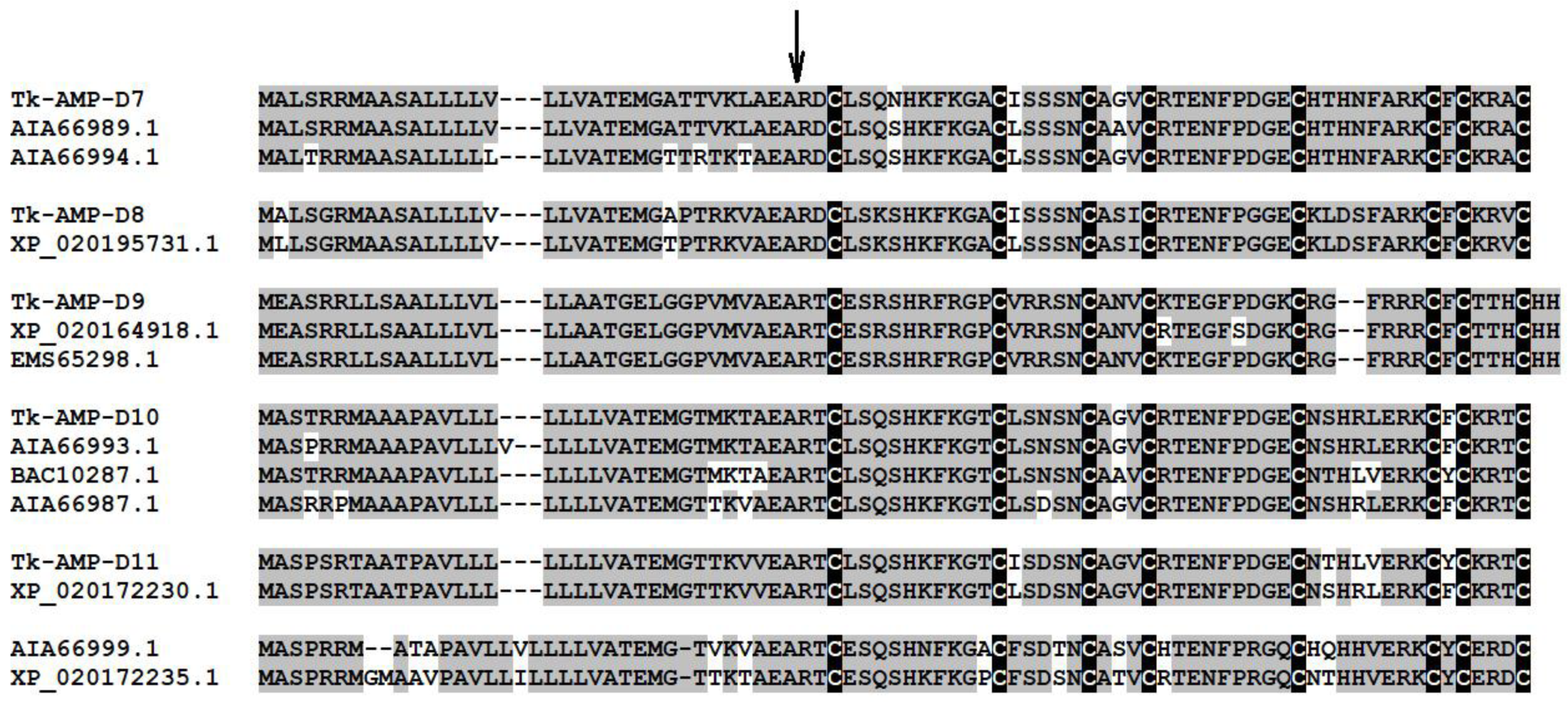
4.9. Gene Expression Assay of Defensins in Germinated Wheat Seeds
4.10. Mycotoxin Quantification After Artificial Seed Inoculation with Toxigenic Fusarium Strains and in Submeged Culture of F. Culmorum.
4.11. Small-Plot Field Experiment Design
4.12. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cook, R.J. Fusarium root, crown, and foot rots and associated seedling diseases. In Compendium of Wheat Diseases and Pests, 3rd ed.; Bockus, W.W., Bowden, R.L., Hunger, R.M., Morrill, W.L., Murray, T.D., Smiley, R.W., Eds.; The Pennsylvania State University Press: University Park, PA, USA, 2010; pp. 37–39. ISBN 978-0890543856. [Google Scholar]
- Moya-Elizondo, E.A.; Rew, L.J.; Jacobsen, B.J.; Hogg, A.C.; Dyer, A.T. Distribution and prevalence of Fusarium crown rot and common root rot pathogens of wheat in Montana. Plant Dis. 2011, 95, 1099–1108. [Google Scholar] [CrossRef]
- Smiley, R.W.; Gourlie, J.A.; Easley, S.A.; Patterson, L.-M.; Whittaker, R.G. Crop damage estimates for crown rot of wheat and barley in the Pacific Northwest. Plant Dis. 2005, 89, 595–604. [Google Scholar] [CrossRef]
- Yli-Mattila, T.; Paavanen-Huhtala, S.; Konstantinova, P.; Parikka, P.; Gagkaeva, T.Y. Molecular and morphological diversity of Fusarium species in Finland and Northwestern Russia. Eur. J. Plant Pathol. 2004, 110, 573–585. [Google Scholar] [CrossRef]
- Khan, M.R.; Fischer, S.; Egan, D.; Doohan, F.M. Biological control of fusarium seedling blight disease of wheat and barley. Phytopathology 2006, 96, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Scäfer, P.; Hückelhoven, R.; Langen, G.; Baltruschat, H.; Stein, E.; Nagarajan, S.; Kogel, K.H. Bipolaris sorokiniana, a cereal pathogen of global concern: Cytological and molecular approaches towards better control. Mol. Plant Pathol. 2002, 3, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Acharya, K.; Dutta, A.K.; Pradhan, P. Bipolaris sorokiniana (Sacc.) Shoem.: The most destructive wheat fungal pathogen in the warmer areas. Aust. J. Crop Sci. 2011, 5, 1064–1071. [Google Scholar]
- Zhong, S.; SAli, S.; Leng, Y.; Wang, R.; Garvin, D.F. Brachypodium distachyon–Cochliobolus sativus pathosystem is a new model for studying plant–fungal interactions in cereal crops. Phytopathology 2015, 105, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Gagkaeva, T.Yu.; Gavrilova, O.P.; Levitin, M.M.; Novozhilov, K.V. Fusarium diseases of grain crops. Plant Protect. Quar. 2011, S5, 69–120. [Google Scholar]
- Arseniuk, E.; Scharen, A.L.; Czembor, H.J. Pathogenicity of seed transmitted Fusarium spp. to triticale seedlings. Mycotoxin Res. 1991, 7, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Burlakoti, R.R.; Shrestha, S.M.; Sharma, R.C. Impact of seed-borne inoculum, irrigation, and cropping pattern on propagation of Bipolaris sorokiniana and epidemiology of foliar blight and common root rot in spring wheat. J. Plant Pathol. 2013, 95, 571–578. [Google Scholar]
- Burlakoti, R.R.; Shrestha, S.M.; Sharma, R.C. Effect of natural seed-borne inoculum of Bipolaris sorokiniana on the seedling emergence and vigour, and early establishment of foliar blight in spring wheat. Arch. Phytopathol. Plant Protect. 2014, 47, 812–820. [Google Scholar] [CrossRef]
- Gagkaeva, T.Y.; Dmitriev, A.P.; Pavlyushin, V.A. Grain microbiota–index of its quality and safety. Plant Protect. Quar. 2012, 9, 14–18. [Google Scholar]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium culmorum: The causal agent of foot and root rot and head blight on wheat. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Sultana, A.; Rashid, A.Q.M.B. Impact of seed transmission of Bipolaris sorokiniana on the planting value of wheat seeds. J. Environ. Sci. Nat. Res. 2012, 5, 75–78. [Google Scholar] [CrossRef]
- Giri, G.K.; Gade, R.M.; Patil, C.U. Seed borne Bipolaris sorokiniana in wheat and its chemical control. J. Soils Crops. 2001, 11, 109–112. [Google Scholar]
- Glynn, N.C.; Ray, R.; Edwards, S.G.; Hare, M.C.; Parry, D.W.; Barnett, C.J.; Beck, J.J. Quantitative Fusarium spp. and Microdochium spp. PCR assays to evaluate seed treatments for the control of Fusarium seedling blight of wheat. J. Appl. Microbiol. 2007, 102, 1645–1653. [Google Scholar] [CrossRef] [PubMed]
- Hysing, S.-C.; Wiik, L. Fusarium seedling blight of wheat and oats: Effects of infection level and fungicide seed treatments on agronomic characters. Acta Agric. Scan. B. 2014, 64, 537–546. [Google Scholar] [CrossRef]
- Moya-Elizondo, E.A.; Jacobsen, B.J. Integrated management of Fusarium crown rot of wheat using fungicide seed treatment, cultivar resistance, and induction of systemic acquired resistance (SAR). Biol. Control. 2016, 92, 153–163. [Google Scholar] [CrossRef]
- Sharma-Poudyal, D.; Sharma, R.C.; Duveiller, E. Control of Helminthosporium leaf blight of spring wheat using seed treatments and single foliar spray in Indo-Gangetic Plains of Nepal. Crop Prot. 2016, 88, 161–166. [Google Scholar] [CrossRef]
- Bello, G.M.D.; Monaco, C.I.; Simon, M.R. Biological control of seedling blight of wheat caused by Fusarium graminearum with beneficial rhizosphere microorganisms. World J. Microbiol. Biotechnol. 2002, 18, 627–636. [Google Scholar] [CrossRef]
- Johansson, P.M.; Johnsson, L.; Gerhardson, B. Suppression of wheat-seedling diseases caused by Fusarium culmorum and Microdochium nivale using bacterial seed treatment. Plant Pathol. 2003, 52, 219–227. [Google Scholar] [CrossRef]
- Knudsen, I.M.B.; Hockenhull, J.; Jensen, D.F. Biocontrol of seedling disease of barley and wheat caused by Fusarium culmorum and Bipolaris sorokiniana: Effects of selected fungal antagonists on growth and yield components. Plant Pathol. 1995, 44, 467–477. [Google Scholar] [CrossRef]
- Mao, W.; Lewis, J.A.; Hebbar, P.K.; Lumsden, R.D. Seed treatment with a fungal or a bacterial antagonist for reducing corn damping-off caused by species of Pythium and Fusarium. Plant Dis. 1997, 81, 450–454. [Google Scholar] [CrossRef]
- Shivanna, M.B.; Meera, M.S.; Hyakumachi, M. Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat. Crop Prot. 1996, 15, 497–504. [Google Scholar] [CrossRef]
- Orzali, L.; Forni, C.; Riccioni, L. Effect of chitosan seed treatment as elicitor of resistance to Fusarium graminearum in wheat. Seed Sci. Technol. 2014, 42, 132–149. [Google Scholar] [CrossRef]
- Roberti, R.; Veronesi, A.R.; Cesari, A.; Cascone, A.; Berardino, I.D.; Bertini, L.; Caruso, C. Induction of PR proteins and resistance by the biocontrol agent Clonostachys rosea in wheat plants infected with Fusarium culmorum. Plant Sci. 2008, 175, 339–347. [Google Scholar] [CrossRef]
- Naresh, M.; Hope, R.; Aldred, D. Ecophysiology of Fusarium culmorum and mycotoxin production. In Advances in Food Mycology; Hocking, A.D., Pitt, J.I., Samson, R.A., Thrane, U., Eds.; Springer: Boston, MA, USA, 2006; Volume 571, pp. 123–136. ISBN 978-0387-28385-2. [Google Scholar] [CrossRef]
- Fernandez, M.R.; Zentner, R.P.; Basnyat, P.; Gehl, D.; Selles, F.; Huber, D. Glyphosate associations with cereal diseases caused by Fusarium spp. in the Canadian Prairies. Europ. J. Agron. 2009, 31, 133–143. [Google Scholar] [CrossRef]
- Tillmann, M.; von Tiedemann, A.; Winte, M. Crop rotation effects on incidence and diversity of Fusarium species colonizing stem bases and grains of winter wheat. J. Plant Dis. Prot. 2017, 124, 121–130. [Google Scholar] [CrossRef]
- Samain, E.; van Tuinen, D.; Jeandetd, P.; Aussenac, T.; Selim, S. The plant-growth-promoting rhizobacterium Paenibacillus sp. strain B2 stimulates wheat defense mechanisms against septoria leaf blotch and root colonization by Curtobacterium plantarumstrain strian EDS. Biol. Control. 2017, 114, 87–96. [Google Scholar] [CrossRef]
- Selim, S.; Negrel, J.; Wendehenne, D.; Ochatt, S.; Gianinazzi, S.; van Tuinen, D. Stimulation of defense reactions in Medicago truncatula by antagonistic lipopeptides from Paenibacillus sp. strain B. Appl. Environ. Microbiol. 2010, 76, 7420–7428. [Google Scholar] [CrossRef] [PubMed]
- Shcherbakova, L.A.; Nazarova, T.A.; Mikityuk, O.D.; Fravel, D.R. Fusarium sambucinum isolate FS-94 induces resistance against Fusarium wilt of tomato via activation and priming of a salicylic acid-dependent signaling system. Russ. J. Plant Physiol. 2011, 58, 808–818. [Google Scholar] [CrossRef]
- Shcherbakova, L.; Semina, J.; Shumilina, D.; Fravel, D.; Dorofeeva, L. Elicitors from biocontrol Fusarium sambucinum and Pseudomonas fluorescens protect wheat from multiple fungal pathogens. IOBC-WPRS Bull. 2012, 83, 249–253. [Google Scholar]
- Shcherbakova, L.; Semina, Yu.; Nazarova, T.; Dorofeeva, L.; Dzhavakhiya, V.; Campbell, B. Potential for integrated control of the wheat pathogen, Stagonospora nodorum, by Folicur and extracellular compounds produced by isolate FS-94 of Fusarium sambucinum. IOBC-WPRS Bull. 2013, 89, 455–458. [Google Scholar]
- Semina, Y.V.; Shcherbakova, L.A.; Devyatkina, G.A. Anti-Stagonospora nodorum activity in filtrates of Fusarium sambucinum cultural liquids after growing the fungus on different nutrient media. Mikol. i Fitopatol. 2011, 45, 563–570. [Google Scholar]
- Semina, Y.V.; Krämer, R.; Shcherbakova, L.A.; Klocke, E.; Nothnagel, T. Studying the possibility of usage of culture filtrate produced by a non-pathogenic Fusarium sambucinum isolate FS-94 for carrot protection against Alternaria radicina. Plant Prot. News 2012, 2, 34–41. [Google Scholar]
- Soytong, K.; Kanokmedhakul, S.; Kukongviriyapaa, V.; Isobe, M. Application of Chaetomium species (Ketomium) as a new broad spectrum biological fungicide for plant disease control: A review article. Fungal Divers. 2001, 7, 1–15. [Google Scholar]
- Mustafa, G.; Tisserant, B.; Randoux, B.; Fontaine, J.; Sahraoui, A.L.; Reignault, P. Mechanisms involved in mycorrhizal wheat protection against powdery mildew. Commun Agric Appl Biol Sci. 2014, 79, 403–410. [Google Scholar] [PubMed]
- Xue, A.G.; Guo, W.; Chen, Y.; Siddiqui, I.; Marchand, G.; Liu, J.; Re, C. Effect of seed treatment with novel strains of Trichoderma spp. on establishment and yield of spring wheat. Crop. Prot. 2017, 96, 97–102. [Google Scholar] [CrossRef]
- Alabouvette, C.; Olivain, C.; Migheli, Q.; Steinberg, C. Microbiological control of soil-borne phytopathogenic fungi with special emphasis on wilt-inducing Fusarium oxysporum. New Phytol. 2009, 184, 529–544. [Google Scholar] [CrossRef] [PubMed]
- Bolwerk, A.; Lagopodi, A.L.; Ben, J.; Lugtenberg, J.; Bloemberg, G.V. Visualization of Interactions between a pathogenic and a beneficial Fusarium strain during biocontrol of tomato foot and root rot. MPMI 2005, 18, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Aime, S.; Alabouvette, C.; Steinberg, C.; Olivain, C. The endophytic strain Fusarium oxysporum Fo47: A good candidate for priming the defense responses in tomato roots. MPMI 2013, 26, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Shcherbakova, L.A.; Odintsova, T.I.; Stakheev, A.A.; Fravel, D.R.; Zavriev, S.K. Identification of a novel small cysteine-rich protein in the fraction from the biocontrol Fusarium oxysporum strain CS-20 that mitigates Fusarium wilt symptoms and triggers defense responses in tomato. Front. Plant Sci. 2016, 6, 1207. [Google Scholar] [CrossRef] [PubMed]
- Soković, M.D.; Glamočlija, J.M.; Ćirić, A.D. Natural Products from Plants and Fungi as Fungicides. In Fungicides–Showcases of Integrated Plant Disease Management from Around the World; Nita, M., Ed.; InTech: Rijeka, Croatia, 2010; pp. 185–232. ISBN 9809533073984. [Google Scholar]
- Thuerig, B.; Binder, A.; Boller, T.; Guyer, U.; Jimenez, S.; Rentsch, C.; Tamm, L. An aqueous extract of the dry mycelium of Penicillium chrysogenum induces resistance in several crops under controlled and field conditions. Eur. J. Plant Pathol. 2006, 114, 185–197. [Google Scholar] [CrossRef]
- Sibounnavong, P.; Kanokmedhakul, S.; Soytong, K. Antifungal activities of Chaetomium brasilense CB01 and Chaetomium cupreum CC03 against Fusarium oxysporum f.sp. lycopersici race 2. IJAT. 2012, 8, 1029–1038. [Google Scholar]
- Narusaka, M.; Minami, T.; Iwabuchi, C.; Hama-saki, T.; Takasaki, S.; Kawamura, K.; Narusaka, Y. Yeast cell wall extract induces disease resistance against bacterial and fungal pathogens in Arabidopsis thaliana and Brassica Crop. PLOS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.J.; Loake, G.J. Role of reactive oxygen intermediates and cognate redox signaling in disease resistance. Plant Physiol. 2000, 124, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Xu, H.; Yi, H.; Yang, L.; Kong, Z.; Zhang, L.; Xue, S.; Jia, H.; Ma, Z. Resistance to hemi-biotrophic F. graminearum infection is associated with coordinated and ordered expression of diverse defense signaling pathways. PLOS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Manners, J.M. Hidden weapons of microbial destruction in plant genomes. Genome Biol. 2007, 8, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Selim, S.; Sanssené, J.; Rossard, S.; Courtois, J. Systemic induction of the defensin and phytoalexin pisatin pathways in pea (Pisum sativum) against Aphanomyces euteiches by acetylated and nonacetylated oligogalacturonides. Molecules 2017, 22, 1017. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Olmedo, F.; Molina, A.; Alamillo, J.M.; Rodriguez-Palenzuela, P. Plant defense peptides. Biopolymers 1998, 47, 479–491. [Google Scholar] [CrossRef]
- Lay, F.T.; Anderson, M.A. Defensins–components of the innate immune system in plants. Curr. Protein Peptide Sci. 2005, 6, 85–101. [Google Scholar] [CrossRef]
- Sels, J.; Mathys, J.; De Coninck, B.M.; Cammue, B.P.; De Bolle, M.F. Plant pathogenesis-related (PR) proteins: A focus on PR peptides. Plant Physiol. Biochem. 2008, 46, 941–950. [Google Scholar] [CrossRef]
- Egorov, T.A.; Odintsova, T.I.; Pukhalsky, V.A.; Grishin, E.V. Diversity of wheat anti-microbial peptides. Peptides 2005, 26, 2064–2073. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, T.I.; Egorov, T.A.; Musolyamov, A.K.; Odintsova, M.S.; Pukhalsky, V.A.; Grishin, E.V. Seed defensins from T. kiharae and related species: Genome localization of defensin-encoding genes. Biochimie 2007, 89, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, T.I.; Rogozhin, E.A.; Baranov, Y.; Musolyamov, A.K.; Yalpani, N.; Egorov, T.A.; Grishin, E.V. Seed defensins of barnyard grass Echinochloa crusgalli (L.) Beauv. Biochimie 2008, 90, 1667–1673. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E.; Arimura, G.I.; Mithoefer, A. Natural elicitors, effectors and modulators of plant responses. Nat. Prod. Rep. 2012, 29, 1288–1303. [Google Scholar] [CrossRef] [PubMed]
- Selim, S.; Roisin-Fichter, C.; Andry, J.B.; Bogdanow, B. Accuracy of real-time PCR to study Mycosphaerella graminicola epidemic in wheat: From spore arrival to fungicide efficiency. In Fungicides–Beneficial and Harmful Aspects; Thajuddin, N., Ed.; InTech: Rijeka, Croatia, 2011; pp. 219–238. ISBN 978-953-307-451-1. [Google Scholar]
- Somai-Jemmali, L.; Selim, S.; Siah, A.; Hamada, W. Fungicide sensitivity of Mycosphaerella graminicola Tunisian isolates: The importance of drug transporter genes in the process of fungicide tolerance. Phytopathol. Mediterr. 2014, 53, 83–93. [Google Scholar] [CrossRef]
- Roze, L.V.; Laivenieks, M.; Hong, S.Y.; Wee, J.; Wong, S.S.; Vanos, B.; Awad, D.; Ehrlich, K.C.; Linz, J.E. Aflatoxin biosynthesis is a novel source of reactive oxygen species–a potential redox signal to initiate resistance to oxidative stress? Toxins 2015, 7, 1411–1430. [Google Scholar] [CrossRef] [PubMed]
- Geisen, R.; Touhami, N.; Schmidt-Heydt, M. Mycotoxins as adaptation factors to food related environments. Curr. Opin. Food Sci. 2017, 17, 1–8. [Google Scholar] [CrossRef]
- Campbell, B.C.; Chan, K.L.; Kim, J.H. Chemosensitization as a means to augment commercial antifungal agents. Front. Microbiol. 2013, 3, 79. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, D.M.; Kazan, K.; Praud, S.; Torney, F.J.; Rusu, A.; Manners, J.M. Early activation of wheat polyamine biosynthesis during Fusarium head blight implicates putrescine as an inducer of trichothecene mycotoxin production. BMC Plant Biol. 2010, 10, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.J.; Orlandi, E.W.; Mock, N.M. Harpin, an elicitor of the hypersensitive response in tobacco caused by Erwinia amylovora, elicits active oxygen production in suspension cells. Plant Physiol. 1993, 102, 1341–1344. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.F.; Summerell, B.A. Media–recipes and preparation. In The Fusarium Laboratory Manual; Blackwell Publishing: Ames, Iowa, IA, USA, 2006; pp. 3–14. ISBN 978-0-813-81919-8. [Google Scholar] [CrossRef]
- Toledo, J.; Reis, E.M.; Forcelini, C.A. Comparison of methods for the detection of Bipolaris sorokiniana in barley seeds. Fitopatol. Bras. 2002, 27, 389–394. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. Species descriptions. In The Fusarium Laboratory Manual; Blackwell Publishing: Ames, Iowa, IA, USA, 2006; pp. 121–174. ISBN 978-0-813-81919-8. [Google Scholar] [CrossRef]
- Pushkareva, V.L.; Slezina, M.P.; Korostyleva, T.V.; Shcherbakova, L.A.; Istomina, E.A.; Ermolaeva, S.A.; Ogarkova, O.A.; Odintsova, T.I. Antimicrobial activity of wild plant seed exstracts against human bacterial and plant fungal pathogens. Am. J. Plant Sci. 2017, 8, 1572–1592. [Google Scholar] [CrossRef]
- Turhan, H.; Baser, I. Callus induction from mature embryo of winter wheat (Triticum aestivum L.). Asian J. Plant Sci. 2004, 3, 17–19. [Google Scholar] [CrossRef]
- Felix, G.; Duran, J.D.; Volko, S.; Boller, T. Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J. 1999, 18, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Chkanikov, D.I.; Sokolova, G.D.; Devyatkina, G.A.; Kazakevich, G.D.; Nazarova, T.A. The toxin biosynthesis by Fusarium graminearum Schw. and its sporulation in a submerged culture. Doklady RASCHN 1994, 6, 22–23. [Google Scholar]
- Greenhalgh, R.; Blackwell, B.A.; Savard, M.; Miller, J.D.; Taylor, A. Secondary metabolites produced by Fusarium sporotrichioides DAOM165006 in liquid culture. Agric. Food Chem. 1988, 36, 216–219. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Concentration of Unlyzed Cells **, 103/mL | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| Untreated cells * | 58.3 ± 3.69 | 58.4 ± 6.03 | 55.0 ± 6.23 |
| Cells + PME | 60.9 ± 4.01 | 56.4 ± 3.94 | 54.7 ± 4.52 |
| Cells + F. culmorum | 28.1 ± 1.53 | 10.7 ± 1.34 | 5.2 ± 1.62 |
| Cells + PME + F. culmorum | 40.4 ± 2.56 | 33.3 ± 2.80 | 17.0 ± 1.39 |
| PME, mg/mL | Mycotoxins, µg/g | |||
|---|---|---|---|---|
| Grain | Submerged Culture | |||
| F. culmorum | F. graminearum | F. culmorum | ||
| DON | AcDON | ZER | DON | |
| 0.2 | 78.54 a | 10.63 c | 0.78 d | not used |
| 0.5 | 74.69 b | 12.88 c | 0.86 d | 0.376 e |
| 1.0 | 61.14 b | 10.55 c | 0.67 d | 0.374 e |
| 0 * | 89.96 a | 14.99 c | 0.94 d | 0.432 e |
| Test Year, Cultivar | Seed Treatment | Development of Root Rots Caused by | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Fusarium spp. | B. sorokiniana | ||||||||
| Incidence | DI, % | Incidence | DI, % | ||||||
| Z29 | Z83 | Z29 | Z83 | Z29 | Z83 | Z29 | Z83 | ||
| 2013, cv. Enita | Control * | 53.7 | 72.3 | 17.7 | 22.6 | 12.7 | 15.9 | 6.4 | 6.7 |
| PME | 23.8 | 43.3 | 9.5 | 13.6 | 6.3 | 9.1 | 1.4 | 2.8 | |
| p= | 0.03 | 0.002 | 0.04 | 0.01 | 0.02 | 0.05 | 0.003 | 0.01 | |
| 2014, cv. Enita | Control * | 52.6 | 67.9 | 18.5 | 20.6 | 14.3 | 17.6 | 4.6 | 8.8 |
| PME | 31.5 | 42.9 | 10.4 | 12.5 | 7.7 | 11.4 | 1.3 | 4.2 | |
| p= | 0.04 | 0.02 | 0.05 | 0.05 | 0.01 | 0.05 | 0.01 | 0.02 | |
| 2015, cv. Enita | Control * | 44.0 | 55.3 | 12.1 | 18.2 | 23.4 | 25.5 | 3.9 | 6.8 |
| PME | 22.6 | 36.5 | 7.5 | 11.6 | 19.4 | 22.7 | 1.3 | 2.7 | |
| p= | 0.05 | 0.004 | 0.05 | 0.05 | 0.07 | 0.08 | 0.04 | 0.03 | |
| 2016, cv. Zlata | Control * | 46.7 | 54.6 | 15.7 | 17.9 | 6.9 | 10.9 | 3.0 | 4.3 |
| PME | 25.8 | 29.6 | 8.3 | 9.7 | 4.1 | 5.3 | 1.2 | 1.8 | |
| p= | 0.04 | 0.02 | 0.004 | 0.05 | 0.05 | 0.04 | 0.003 | 0.002 | |
| Pme-20C ** | 39.6 | 48.7 | 9.9 | 11.5 | 5.0 | 9.4 | 2.1 | 3.1 | |
| p= | 0.06 | 0.08 | 0.03 | 0.05 | 0.09 | 0.09 | 0.04 | 0.05 | |
| Test Year | Seed Treatment | Yield Parameters * | |||
|---|---|---|---|---|---|
| Number of Productive Stems Per Plot | Weight of 1000 Kernels, g | Total Grain Weight from One Plot, g | Yield Increase, % of Control ** | ||
| cv. Enita | |||||
| 2013 | PME | 490 a | 37.3 a | 540.1 a | 8.4 |
| Untreated seeds | 483 a | 32.5 b | 498.4 b | ||
| 2014 | PME | 523 a | 36.8 a | 533.8 a | 6.0 |
| Untreated seeds | 498 a | 34.4 b | 486.2 b | ||
| 2015 | PME | 475 a | 32.2 b | 501.9 b | 2.0 *** |
| Untreated seeds | 467 a | 30.6 b | 492.2 b | ||
| cv. Zlata | |||||
| 2016 | PME | 541 b | 37.2 a | 581.2 c | 7.3 |
| Untreated seeds | 464 a | 33.8 b | 541.5 a | ||
| # | Peptide Accession Number | mRNA Accession Number and Annotation | Primer Sequences (5′→3′); Annealing Temperature (T),0C; Fragment Length (L), bp | T. kiharae defensin | Sequence similarity, (%) |
|---|---|---|---|---|---|
| 1 | AIA66989.1 | KJ551519.1 T. aestivum, defensin (PDF4) | FOR AGCTGAGCAGATCGATGGCG REV GGCTAGCAGGCCCTCTTGCA T=60 L=265 | Tk−AMP−D7 | 96% |
| 2 | AIA66994.1 | KJ551524.1 T. aestivum, defensin (PDF9) | 90% | ||
| 3 | BAC10287.1 | AB089942.1 T. aestivum, defensin (Tad1) | FOR GTGAAGCGAGCAAGCAGAGAGA REV TAGGGACGAACAGATCTAA T=60 L=360 | Tk−AMP−D8 | 94% |
| 4 | AIA66993.1 | KJ551523.1 T. aestivum, defensin (PDF8) | 98% | ||
| 5 | AIA66987.1 | KJ551517.1 T. aestivum, defensin (PDF2) | 94% | ||
| 6 | XP_020164918.1 | XM_020309329.1 Ae. Tauschii, defensin Ec-AMP-D2-like (LOC109750363) | FOR GCTGCTCACACACAACACAC REV CAGAAAGGCCACCCGAAAGA T=59 L=372 | Tk−AMP−D9 | 98% |
| 7 | XP_020195731.1 | XM_020340142.1 Ae. Tauschii, AMP-D1.2-like (LOC109781554) | FOR GCTAGCTTTACACACAGCCC REV ACCGTAGCTAGCATCGGACC T=59 L=330 | Tk−AMP−10 | 96% |
| 8 | XP_020172230.1 | XM_020316641.1 Ae. Tauschii, defensin Tk-AMP-D2-like (LOC109757808) | FOR CAAGCAGAGAGATGGCGTCC REV CGGATGCATGGAGATGAACCA T=59 L=402 | Tk−AMP−11 | 94% |
| 9 | AIA66999.1 | KJ551529.1 T. aestivum, defensin (PDF14) | FOR AGAGAGCAAGTGCAGAAGAGA REV AAGCTCAGCAGTCCCGCTCGCA T=69 L=281 | - | - |
| 10 | XP_020172235.1 | XM_020316646.1 Ae. Tauschii, defensin Tk-AMP-D2 (LOC109757811) | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shcherbakova, L.A.; Nazarova, T.A.; Mikityuk, O.D.; Istomina, E.A.; Odintsova, T.I. An Extract Purified from the Mycelium of a Tomato Wilt-Controlling Strain of Fusarium sambucinum Can Protect Wheat against Fusarium and Common Root Rots. Pathogens 2018, 7, 61. https://doi.org/10.3390/pathogens7030061
Shcherbakova LA, Nazarova TA, Mikityuk OD, Istomina EA, Odintsova TI. An Extract Purified from the Mycelium of a Tomato Wilt-Controlling Strain of Fusarium sambucinum Can Protect Wheat against Fusarium and Common Root Rots. Pathogens. 2018; 7(3):61. https://doi.org/10.3390/pathogens7030061
Chicago/Turabian StyleShcherbakova, Larisa A., Tatyana A. Nazarova, Oleg D. Mikityuk, Ekaterina A. Istomina, and Tatyana I. Odintsova. 2018. "An Extract Purified from the Mycelium of a Tomato Wilt-Controlling Strain of Fusarium sambucinum Can Protect Wheat against Fusarium and Common Root Rots" Pathogens 7, no. 3: 61. https://doi.org/10.3390/pathogens7030061
APA StyleShcherbakova, L. A., Nazarova, T. A., Mikityuk, O. D., Istomina, E. A., & Odintsova, T. I. (2018). An Extract Purified from the Mycelium of a Tomato Wilt-Controlling Strain of Fusarium sambucinum Can Protect Wheat against Fusarium and Common Root Rots. Pathogens, 7(3), 61. https://doi.org/10.3390/pathogens7030061