Proteomic Differences between Listeria monocytogenes Isolates from Food and Clinical Environments



Abstract
:1. Introduction
2. Results and Discussion
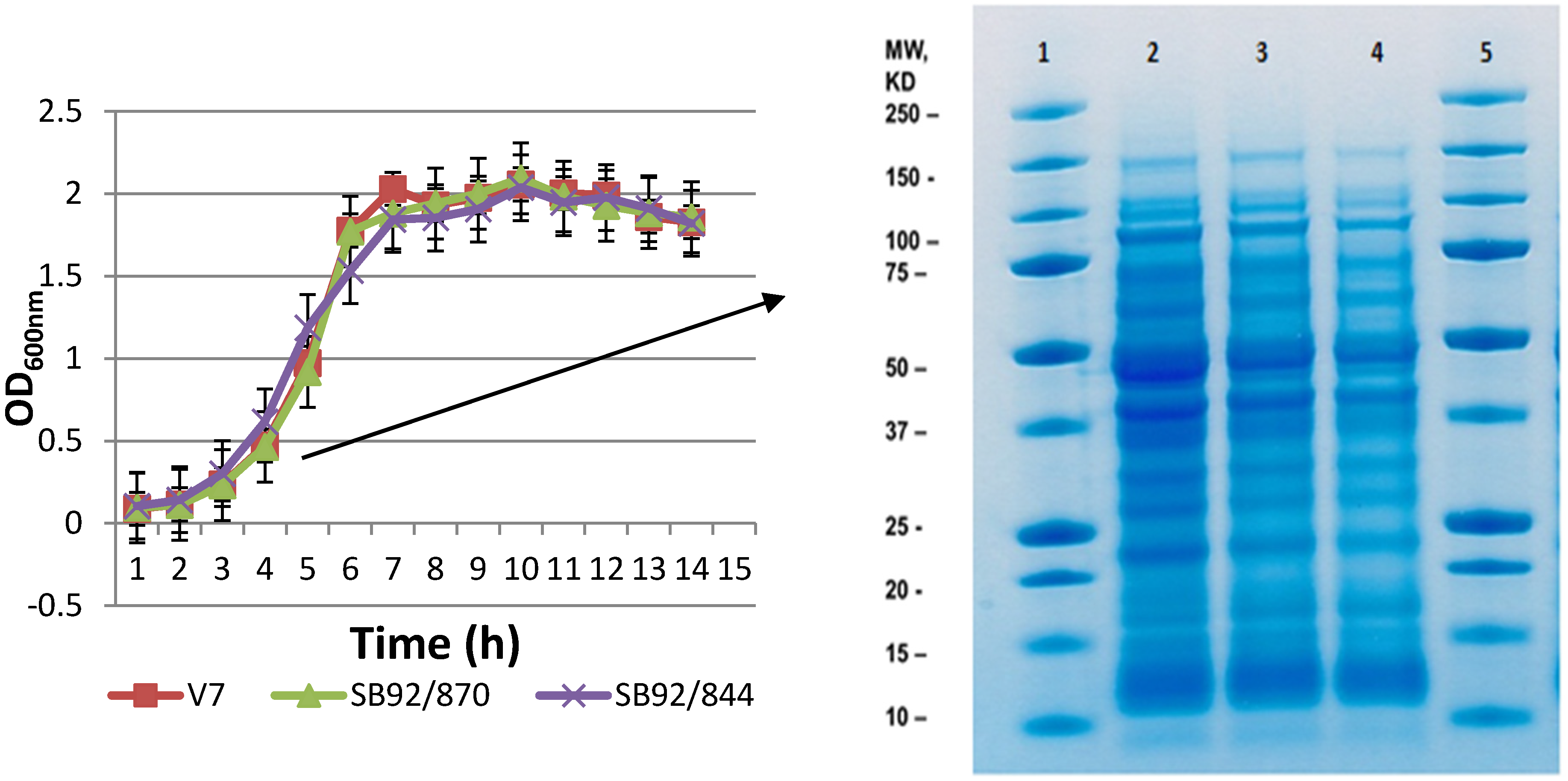
2.1. Growth and 1D SDS PAGE Profiles
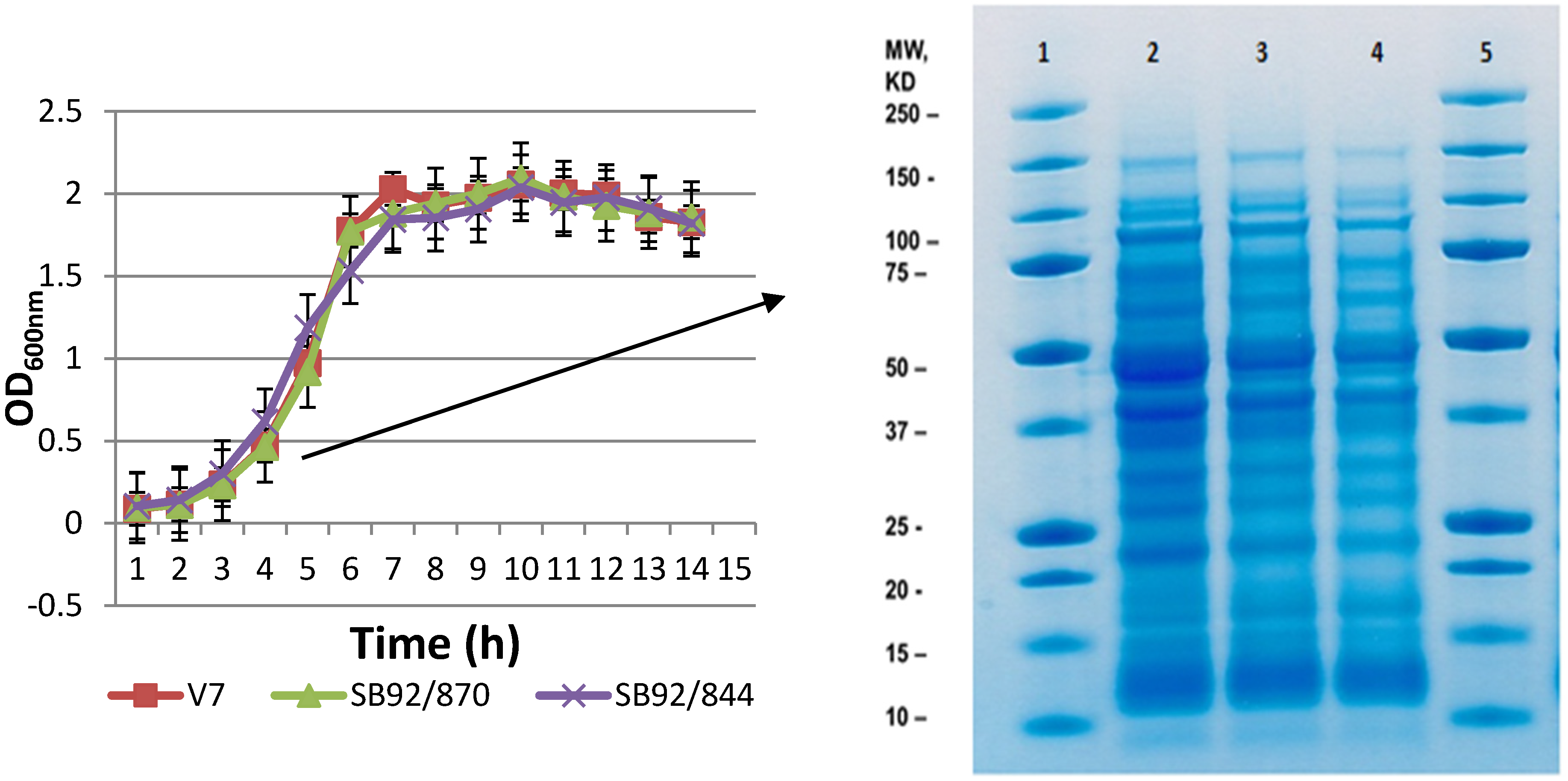
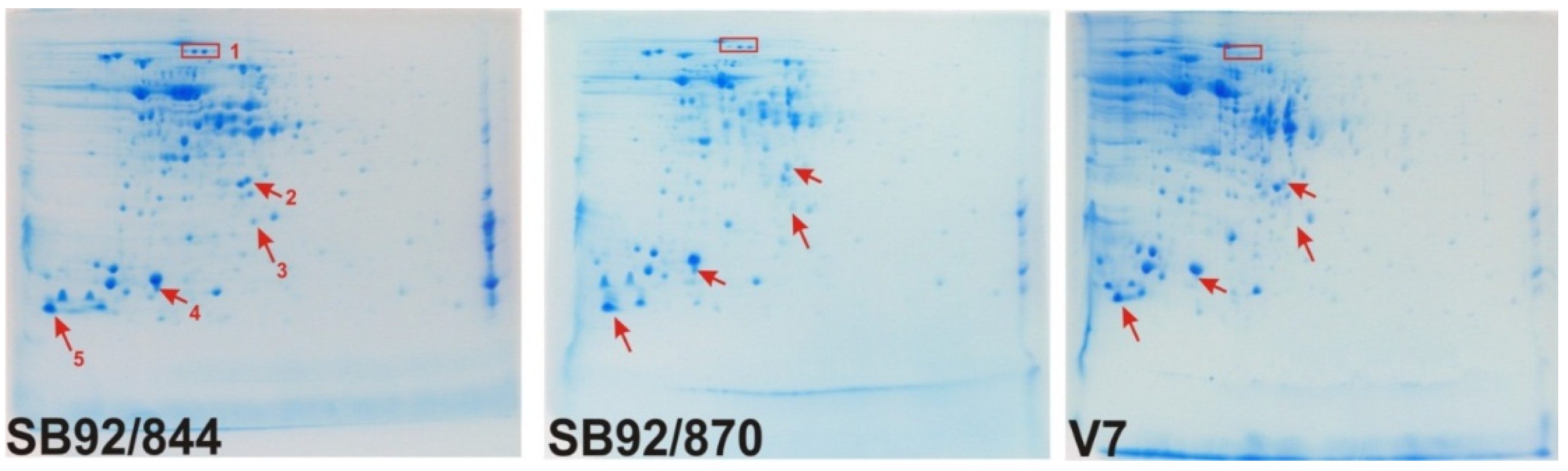
2.2. 2DE Proteome Profiles
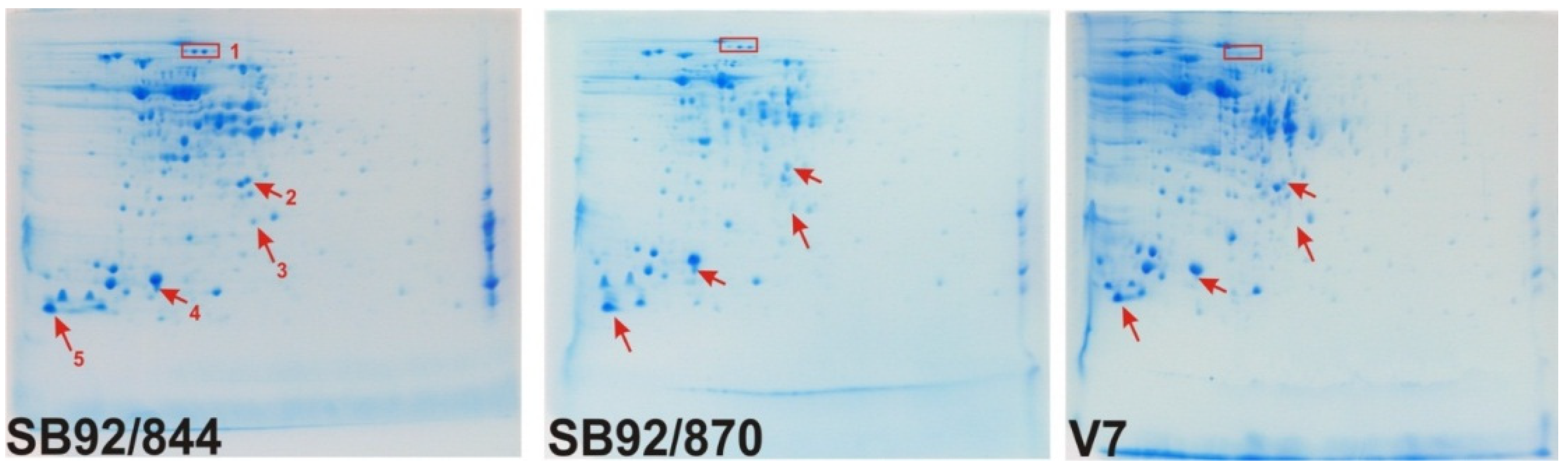

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. a | Identified Protein | SB92/844 vs. V7 b | SB92/870 vs. V7 c | SB92/844 vs. SB92/870 d |
|---|---|---|---|---|
| 0001 | Major cold-shock protein | Protein only present in SB92/844 | ||
| 0002 | Major cold-shock protein, partial | ↑(+1.8) | ↑(+1.7) | (+1.0) |
| 0004 | Major cold-shock protein, partial | (+1.2) | ↑(+1.7) | (−1.3) |
| 1001 | Major cold-shock protein, partial | (−1.1) | (+1.4) | ↓(−1.6) |
| 1003 | 50S ribosomal protein L7/L12 | ↑(+2.0) | (+1.2) | ↑(+1.6) |
| 1103 | Regulatory protein SpoVG | (+1.0) | (+1.3) | (−1.3) |
| 2001 | co-chaperonin GroES | ↓(−1.9) | ↓(−1.8) | (−1.1) |
| 2402 | CD4+ T-cell-stimulating antigen | Protein not present in V7, but present in SB92/870 | ↓(−1.7) | |
| 3004 | Phosphocarrier protein HPr | (+1.2) | ↑(+1.5) | (−1.3) |
| 3005 | Phosphocarrier protein HPr | Protein not present in V7, but present in SB92/870 | ↓(−1.7) | |
| 5802 | Glyceraldehyde-3-phosphate dehydrogenase | Protein not present in V7, but present in SB92/870 | (−1.1) | |
| 5803 | Glyceraldehyde-3-phosphate dehydrogenase | Protein not present in V7, but present in SB92/870 | (+1.1) | |
| 6102 | Superoxide dismutase | ↓(−1.6) | ↓(−2.0) | (+1.3) |
| 6205 | Ribosome recycling factor | ↑(+2.3) | ↑(+1.7) | (+1.3) |
| 6502 | Glyceraldehyde-3-phosphate dehydrogenase | ↓(−3.0) | ↓(−3.1) | (+1.1) |
| 6603 | Glyceraldehyde-3-phosphate dehydrogenase | (+1.0) | ↓(−2.6) | ↑(+2.6) |
| 7102 | PTS system, glucose-specific, IIA component, putative | Protein not present in V7, but present in SB92/870 | ↓(−4.2) | |
| 7201 | Phosphoglycerate mutase | ↓(−1.9) | Protein not present in SB92/870 | |
| 9203 | Major cold-shock protein | ↑(+3.5) | ↑(+1.5) | ↓(−5.4) |
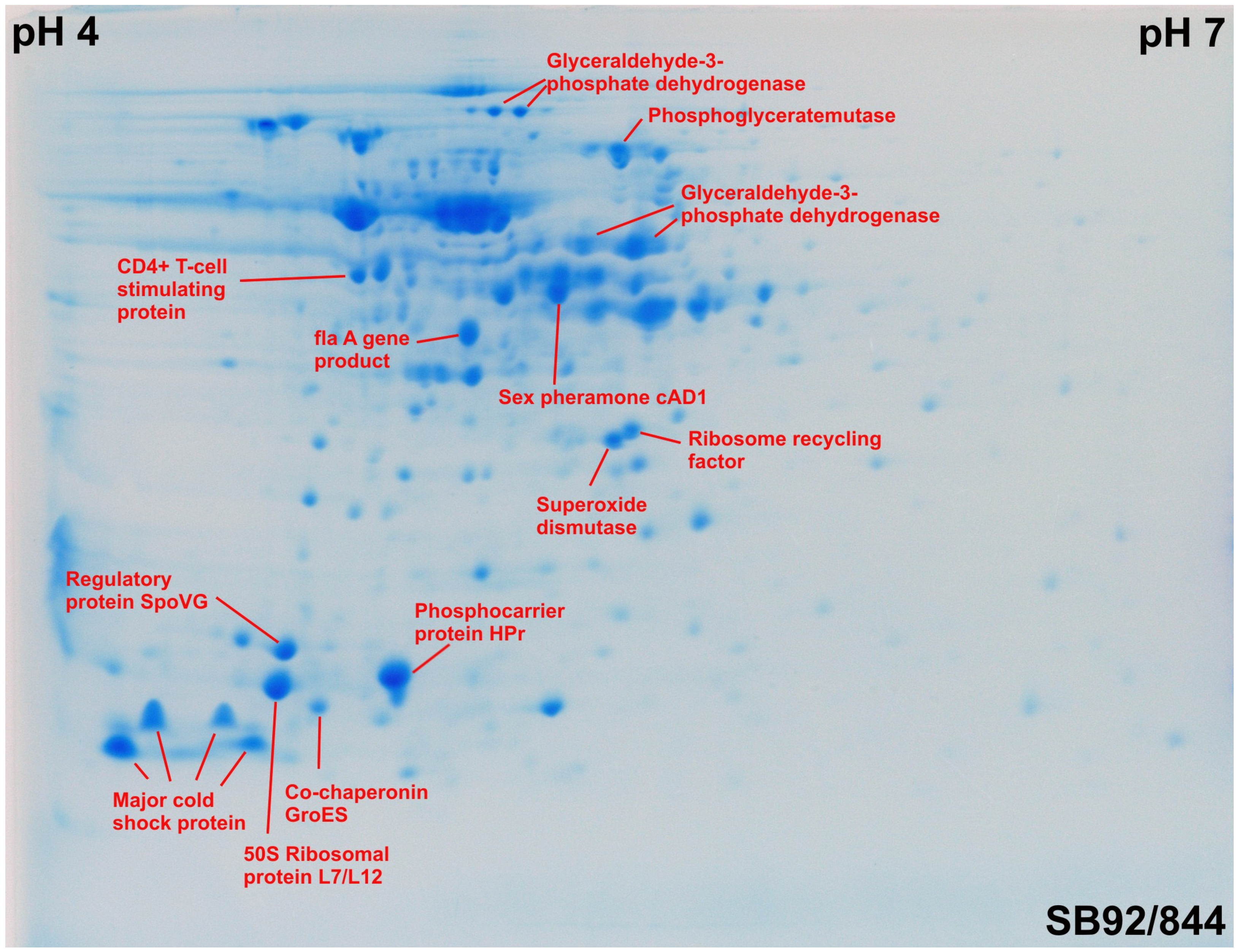
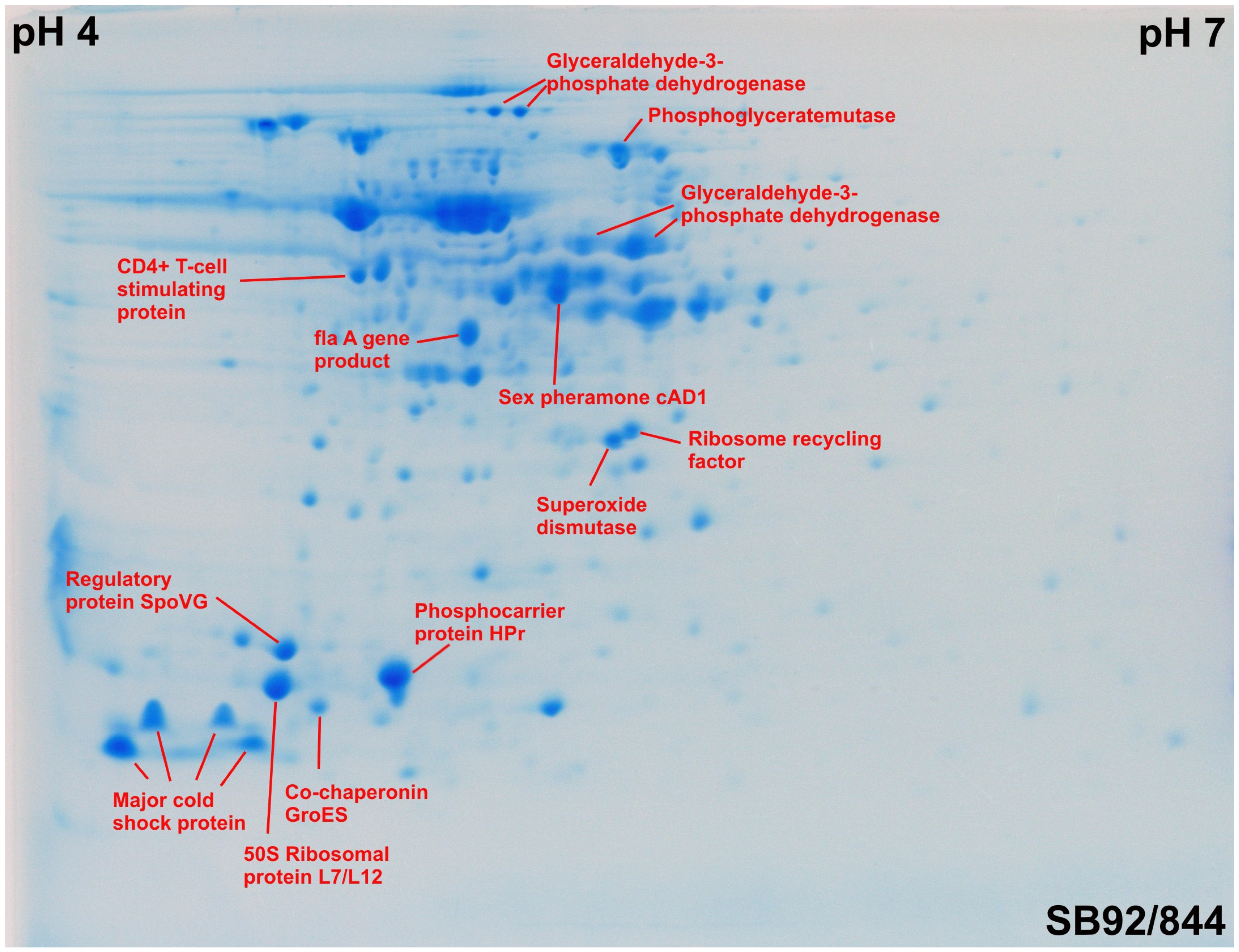
2.3. Identification of Proteins by MALD-TOF MS and Ion Trap MS
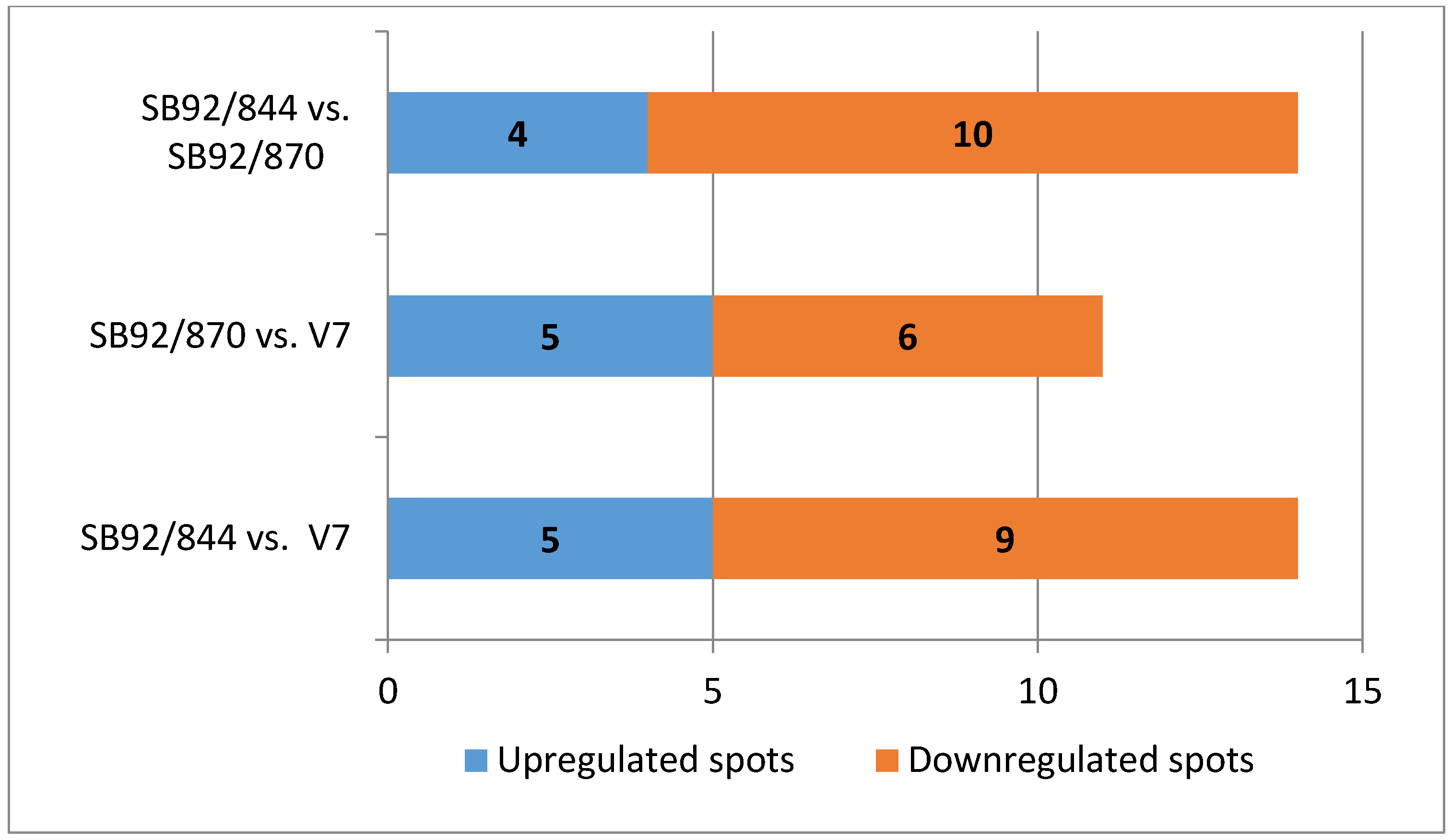
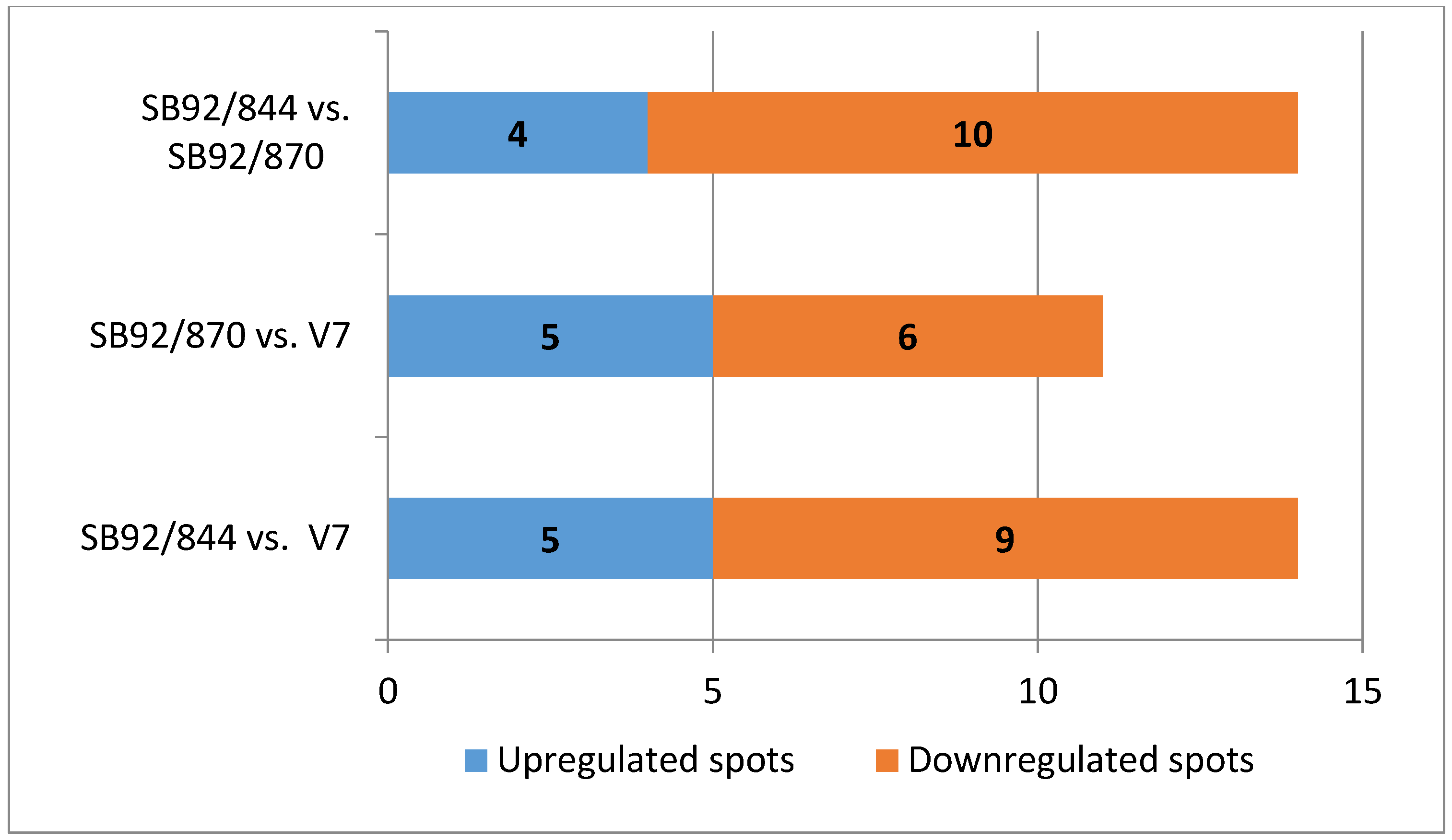
2.4. Comparative Analysis of Up and Down Regulated Proteins
3. Experimental Section
3.1. Bacterial Strains and Growth Conditions
3.2. Protein Extraction
3.3. 1D SDS PAGE
3.4. 2DE and MS Analysis
3.5. Protein Identification and Classification
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Orsi, R.H.; Bakker, H.C.D.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar]
- Trost, M.; Wehmhoner, D.; Karst, U.; Dieterich, G.; Wehland, J.; Jansch, L. Comparative proteome analysis of secretory proteins from pathogenic and nonpathogenic Listeria species. Proteomics 2005, 5, 1544–1557. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.D.; Wallbanks, S.; Lane, D.J.; Shah, J.; Nietupski, R.; Smida, J.; Dorsch, M.; Stackebrandt, E. Phylogenetic analysis of the genus Listeria based on reverse transcriptase sequencing of 16S r RNA. Int. J. Syst. Evol. Microbiol. 1991, 41, 240–246. [Google Scholar]
- Graves, L.M.; Helsel, L.O.; Steigerwalt, A.G.; Morey, R.E.; Daneshvar, M.I.; Roof, S.E.; Orsi, R.H.; Fortes, E.D.; Milillo, S.R.; den Bakker, H.C.; et al. Listeria marthii sp. nov., isolated from the natural environment, Finger Lakes National Forest. Int. J. Syst. Evol. Microbiol. 2009, 60, 1280–1288. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, B.; Gerner-Smidt, P. The epidemiology of human listeriosis. Microbes Infect. 2007, 9, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607–625. [Google Scholar] [CrossRef] [PubMed]
- Bryan, F.L. The “danger zone” revaluated. Food Saf. Mag. 2004, 10, 55–69. [Google Scholar]
- Cabrita, P.; Fonseca, C.; Freitas, R.; Carreira, R.; Capelo, J.L.; Trigo, M.J.; Ferreira, R.B.; Brito, L. A secretome-based methodology may provide a better characterization of the virulence of Listeria monocytogenes: Preliminary results. Talanta 2010, 83, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Cacace, G.; Mazzeo, M.F.; Sorrentino, A.; Spada, V.; Malorni, A.; Siciliano, R.A. Proteomics for the elucidation of cold adaptation mechanisms in Listeria monocytogenes. J. Proteomics 2010, 73, 2021–2030. [Google Scholar] [CrossRef] [PubMed]
- Duché, O.; Trémoulet, F.; Namane, A.; Labadie, J.; European Listeria Genome Consortium. A proteomic analysis of the salt stress response of Listeria monocytogenes. FEMS Microbiol. Lett. 2002, 215, 183–188. [Google Scholar]
- Folio, P.; Chavant, P.; Belkorchia, A.; Chambon, C.; Hébraud, M. Two-dimensional electrophoresis database of Listeria monocytogenes EGDe proteome and proteomic analysis of mid-log and stationary growth phase cells. Proteomics 2000, 10, 3187–3201. [Google Scholar]
- Gardan, R.; Duché, O.; Leroy-Se’trin, S.; Labadie, J. Role of ctc from Listeria monocytogenes in osmotolerance. Appl. Environ. Microbiol. 2003, 69, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Ramnath, M.; Rechinger, K.B.; Jansch, L.; Hastings, J.W.; Knochel, S.; Gravesen, A. Development of a Listeria monocytogenes EGDe partial proteome referrence map and comparison with the protein profiles of food isolates. Appl. Environ. Microbiol. 2003, 69, 3368–3376. [Google Scholar] [CrossRef] [PubMed]
- Wemekamp-Kamphuis, H.H.; Wouters, J.A.; de Leeuw, P.P.; Hain, T.; Chakraborty, T.; Abee, Y. Identification of sigma factor sigma B-controlled genes and their impact on acid stress, high hydrostatic pressure, and freeze survival in Listeria monocytogenes EGD-e. Appl. Environ. Microbiol. 2004, 70, 3457–3466. [Google Scholar] [CrossRef] [PubMed]
- Hefford, M.A.; D’Aoust, S.; Cyr, T.D.; Austin, J.W.; Sanders, G.; Kheradpir, E.; Kalmokoff, M.L. Proteomic and microscopic analysis of biofilms formed by Listeria monocytogenes 568. Can. J. Microbiol. 2005, 51, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Dumas, E.; Meunier, B.; Berdague, J.L.; Chambon, C.; Desvaux, M.; Hebraud, M. Comparative analysis of extracellular and intracellular proteomes of Listeria monocytogenes strains reveals a correlation between protein expression and serovar. Appl. Environ. Microbiol. 2008, 74, 7399–7409. [Google Scholar] [CrossRef] [PubMed]
- Van de Velde, S.; Delaive, E.; Dieu, M.; Carryn, S.; Bambeke, F.V.; Devreese, B.; Raes, M.; Tulkens, P.M. Isolation and 2-D-DIGE proteomic analysis of intracellular and extracellular forms of Listeria monocytogenes. Proteomics 2009, 9, 5484–5496. [Google Scholar] [CrossRef]
- Calvo, E.; Pucciarelli, M.G.; Bierne, H.; Cossart, P.; Albar, J.P.; Portillo, F.G. Analysis of the Listeria cell wall proteome by two-dimensional nanoliquid chromatography coupled to amss spectrometry. Proteomics 2005, 5, 433–443. [Google Scholar] [PubMed]
- Donaldson, J.R.; Nanduri, B.; Burgess, S.C.; Lawrence, M. Comparative proteomic analysis of Listeria monocytogenes strains F2365 and EGD. Appl. Environ. Microbiol. 2009, 75, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Pucciarelli, M.G.; Calvo, E.; Sabet, C.; Bierne, H.; Cossart, P.; Portillo, F.G. Indeitification of substrates of the Listeria monocytogenes sortases A and B by a non-gel proteomic analysis. Proteomics 2005, 5, 4808–4817. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Hussain, M.A. Advances in proteomics-based detection techniques of Listeria monocytogenes: a potential risk in New Zealand. Internet J. Food Safety. 2012, 14, 70–74. [Google Scholar]
- Trémoulet, F.; Duché, O.; Namane, A.; Martinie, B.; Labadie, J.C. Comparison of protein patterns of Listeria monocytogenes grown in biofilm or in planktonic mode by proteomic analysis. FEMS Microbiol. Lett. 2002, 210, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Phan-Thanh, L.; Gormon, T. Analysis of heat and cold shock proteins in Listeria by two-dimensional electrophoresis. Electrophoresis 1995, 16, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Jedrzejas, M.J. Structure, function, and evolution of phosphoglycerate mutases: Comparison with fructose-2,6-bisphosphatase, acid phosphatase, and alkaline phosphatase. Prog. Biophys. Mol. Biol. 2000, 73, 263–287. [Google Scholar] [CrossRef] [PubMed]
- Rigden, D.C.; LittleJohn, J.E.; Henderson, K.; Jedrzejas, M.J. Structures of phosphate and trivanadate complexes of Bacillus stearothermophilus phosphatase PhoE: Structural and functional analysis in the cofactor-dependent phosphoglycerate mutase superfamily. J. Mol. Biol. 2003, 325, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Nørrung, B.; Andersen, J.K. Variations in virulence between different electrophoretic types of Listeria monocytogenes. Lett. Appl. Microbiol. 2000, 30, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Loepfe, C.; Raimann, E.; Stephan, R.; Tasara, T. Reduced host cell invasiveness and oxidative stress tolerance indouble and triple csp gene family deletion mutants of Listeria monocytogenes. Foodborne Pathog. Dis. 2010, 7, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Duodu, S.; Holst-Jensen, A.; Skjerdal, T.; Cappelier, J.-M.; Pilet, M.-F.; Loncarevic, S. Influence of storage temperature on gene expression and virulence potential of Listeria monocytogenes strains grown in a salmon matrix. Food Microbiol. 2010, 27, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Johansson, J.; Mandin, P.; Renzoni, A.; Chiaruttini, C.; Springer, M.; Cossart, P. An RNA thermosensor controls expression of virulence genes in Listeria monocytogenes. Cell 2002, 110, 551–561. [Google Scholar] [CrossRef] [PubMed]
- McGann, P.; Wiedmann, M.; Boor, K.J. The alternative sigma factor sigma B and the virulence gene regulator PrfA both regulate transcription of Listeria monocytogenes internalins. Appl. Environ. Microbiol. 2007, 73, 2919–2930. [Google Scholar] [CrossRef] [PubMed]
- Ermolenko, D.N.; Makhatadze, G.I. Bacterial cold-shock proteins. Cell. Mol. Life Sci. 2002, 59, 1902–1913. [Google Scholar] [CrossRef] [PubMed]
- Phadtare, S.; Alsina, J.; Inouye, M. Cold-shock response and cold-shock proteins. Curr. Opin. Microbiol. 1999, 2, 175–180. [Google Scholar] [PubMed]
- Murray, E.G.D.; Webb, R.E.; Swann, M.B.R. A disease of rabbits characterized by a large mononuclear leucocytosis, caused by a hitherto undescribed bacillus Bacterium monocytogenes (n. sp.). J. Pathol. Bacteriol. 1926, 29, 407–439. [Google Scholar] [CrossRef]
- Dykes, G.A.; Moorhead, S.M. Survival of osmotic and acid stress by Listeria monocytogenes strains of clinical or meat origin. Int. J. Food Microbiol. 2000, 56, 161–166. [Google Scholar] [CrossRef]
- Cacace, G.; Mazzeo, M.F.; Sorrentino, A.; Spada, V.; Malorni, A.; Siciliano, R.A. Proteomics for the elucidation of cold adaptation mechanisms in Listeria monocytogenes. J. Proteomics. 2010, 73, 2021–2030. [Google Scholar] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, G.; Mason, S.L.; Hudson, J.A.; Clerens, S.; Plowman, J.E.; Hussain, M.A. Proteomic Differences between Listeria monocytogenes Isolates from Food and Clinical Environments. Pathogens 2014, 3, 920-933. https://doi.org/10.3390/pathogens3040920
Huang G, Mason SL, Hudson JA, Clerens S, Plowman JE, Hussain MA. Proteomic Differences between Listeria monocytogenes Isolates from Food and Clinical Environments. Pathogens. 2014; 3(4):920-933. https://doi.org/10.3390/pathogens3040920
Chicago/Turabian StyleHuang, Ge, Susan L. Mason, J. Andrew Hudson, Stefan Clerens, Jeffrey E. Plowman, and Malik A. Hussain. 2014. "Proteomic Differences between Listeria monocytogenes Isolates from Food and Clinical Environments" Pathogens 3, no. 4: 920-933. https://doi.org/10.3390/pathogens3040920
APA StyleHuang, G., Mason, S. L., Hudson, J. A., Clerens, S., Plowman, J. E., & Hussain, M. A. (2014). Proteomic Differences between Listeria monocytogenes Isolates from Food and Clinical Environments. Pathogens, 3(4), 920-933. https://doi.org/10.3390/pathogens3040920