


A Characterization of the Humoral Immune Response to Human Endogenous Retroviruses and Mycobacterium paratuberculosis in Crohn’s Disease
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Blood Samples
2.3. Peptides
2.4. ELISA Assays
2.5. Statistical Analysis
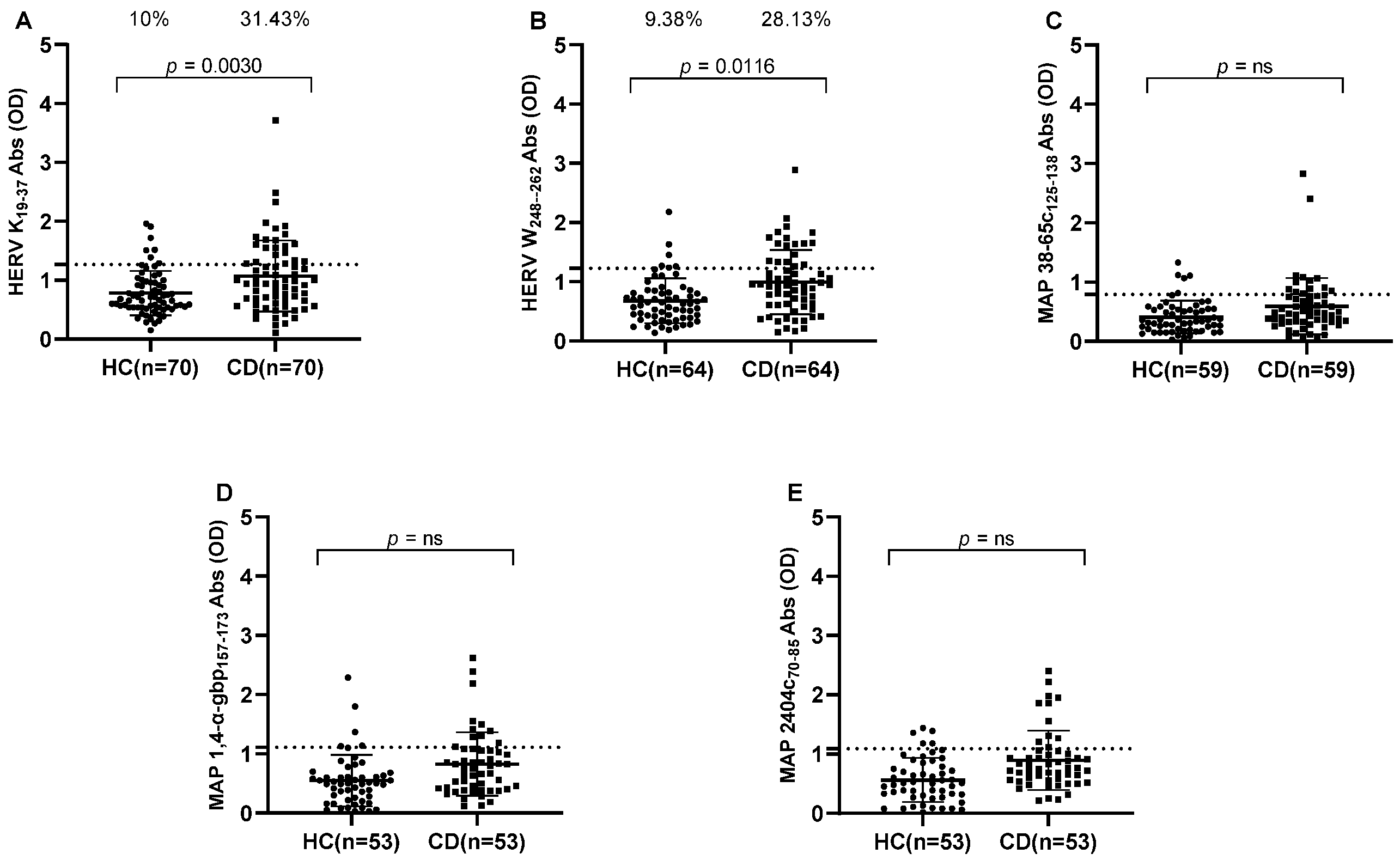
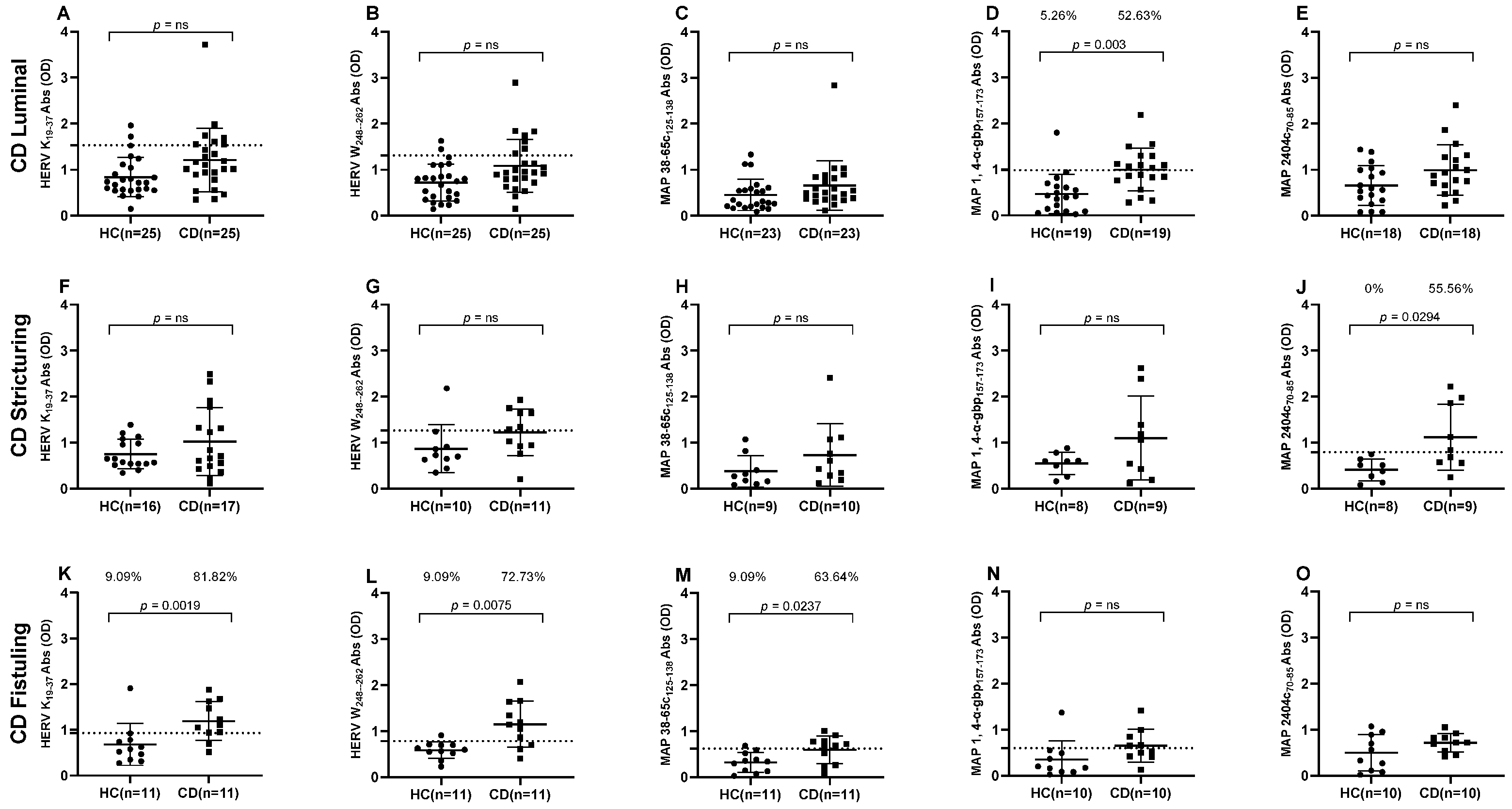
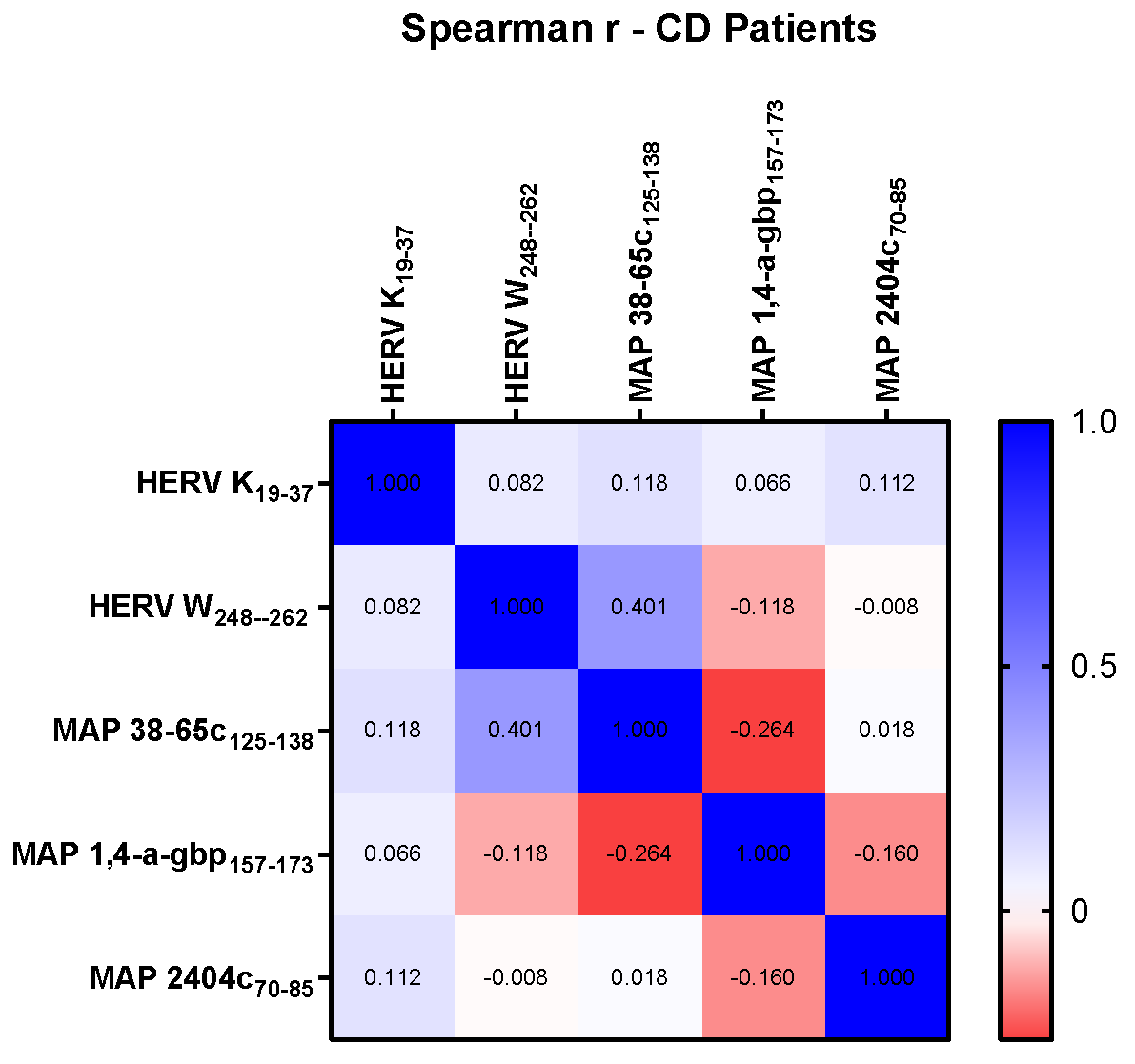
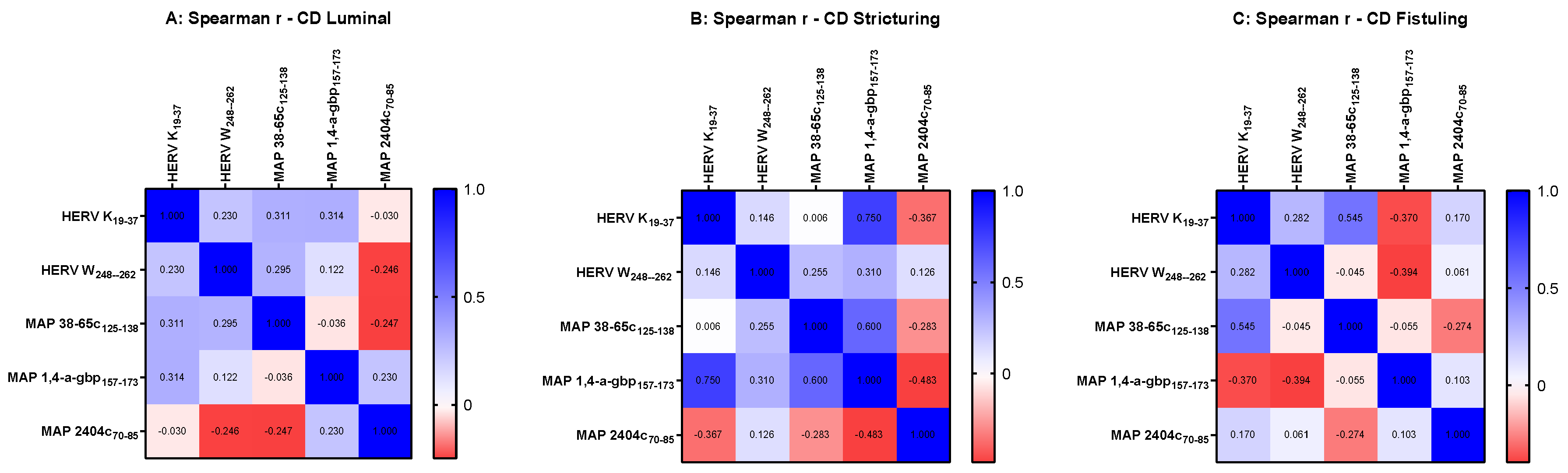
3. Results
3.1. Antibody Response to HERV and MAP Immunogenic Epitopes in Crohn’s Disease Patients Versus Healthy Controls
3.2. Correlation Analyses of HERV and MAP Antibodies
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mintz, M.J.; Lukin, D.J. Mycobacterium avium subspecies paratuberculosis (MAP) and Crohn’s disease: The debate continues. Transl. Gastroenterol. Hepatol. 2023, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Louis, E.; Collard, A.; Oger, A.; Degroote, E.; El Yafi, F.A.N.; Belaiche, J. Behaviour of Crohn’s disease according to the Vienna classification: Changing pattern over the course of the disease. Gut 2001, 49, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Choung, R.; Princen, F.; Stockfisch, T.; Torres, J.; Maue, A.; Porter, C.; Leon, F.; De Vroey, B.; Singh, S.; Riddle, M. Serologic microbial associated markers can predict Crohn’s disease behaviour years before disease diagnosis. Aliment. Pharmacol. Ther. 2016, 43, 1300–1310. [Google Scholar] [CrossRef] [PubMed]
- Sechi, L.A.; Scanu, A.M.; Molicotti, P.; Cannas, S.; Mura, M.; Dettori, G.; Fadda, G.; Zanetti, S. Detection and isolation of Mycobacterium avium subspecies paratuberculosis from intestinal mucosal biopsies of patients with and without Crohn’s disease in Sardinia. Am. J. Gastroenterol. 2005, 100, 1529–1536. [Google Scholar] [CrossRef]
- Zhang, P.; Minardi, L.; Kuenstner, J.; Zhang, S.; Zekan, S.; Kruzelock, R. Serological testing for Mycobacterial heat shock protein HSP65 antibody in health and diseases. Microorganisms 2019, 8, 47. [Google Scholar] [CrossRef]
- Aitken, J.M.; Phan, K.; Bodman, S.E.; Sharma, S.; Watt, A.; George, P.M.; Agrawal, G.; Tie, A.B. A Mycobacterium species for Crohn’s disease? Pathology 2021, 53, 818–823. [Google Scholar] [CrossRef]
- Bernstein, C.N.; Blanchard, J.F.; Rawsthorne, P.; Collins, M.T. Population-based case control study of seroprevalence of Mycobacterium paratuberculosis in patients with Crohn’s disease and ulcerative colitis. J. Clin. Microbiol. 2004, 42, 1129–1135. [Google Scholar] [CrossRef]
- Dupressoir, A.; Lavialle, C.; Heidmann, T. From ancestral infectious retroviruses to bona fide cellular genes: Role of the captured syncytins in placentation. Placenta 2012, 33, 663–671. [Google Scholar] [CrossRef]
- Stein, R.A.; DePaola, R.V. Human endogenous retroviruses: Our genomic fossils and companions. Physiol. Genom. 2023, 55, 249–258. [Google Scholar] [CrossRef]
- Johnson, W.E. Origins and evolutionary consequences of ancient endogenous retroviruses. Nat. Rev. Microbiol. 2019, 17, 355–370. [Google Scholar] [CrossRef]
- Santoni, F.A.; Guerra, J.; Luban, J. HERV-H RNA is abundant in human embryonic stem cells and a precise marker for pluripotency. Retrovirology 2012, 9, 111. [Google Scholar] [CrossRef] [PubMed]
- Soygur, B.; Sati, L. The role of syncytins in human reproduction and reproductive organ cancers. Reproduction 2016, 152, R167–R178. [Google Scholar] [CrossRef] [PubMed]
- Durnaoglu, S.; Lee, S.-K.; Ahnn, J. Syncytin, envelope protein of human endogenous retrovirus (HERV): No longer ‘fossil’in human genome. Anim. Cells Syst. 2021, 25, 358–368. [Google Scholar] [CrossRef]
- Noli, M.; Meloni, G.; Ruberto, S.; Jasemi, S.; Simula, E.R.; Cossu, D.; Bo, M.; Palermo, M.; Sechi, L.A. HERV-K envelope protein induces long-lasting production of autoantibodies in T1DM patients at onset in comparison to ZNT8 autoantibodies. Pathogens 2022, 11, 1188. [Google Scholar] [CrossRef]
- Garcia-Montojo, M.; Fathi, S.; Rastegar, C.; Simula, E.R.; Doucet-O’Hare, T.; Cheng, Y.H.; Abrams, R.P.; Pasternack, N.; Malik, N.; Bachani, M. TDP-43 proteinopathy in ALS is triggered by loss of ASRGL1 and associated with HML-2 expression. Nat. Commun. 2024, 15, 4163. [Google Scholar] [CrossRef] [PubMed]
- Balestrieri, E.; Minutolo, A.; Petrone, V.; Fanelli, M.; Iannetta, M.; Malagnino, V.; Zordan, M.; Vitale, P.; Charvet, B.; Horvat, B. Evidence of the pathogenic HERV-W envelope expression in T lymphocytes in association with the respiratory outcome of COVID-19 patients. EBioMedicine 2021, 66, 103341. [Google Scholar] [CrossRef]
- Li, W.; Xue, X.; Li, X.; Wu, X.; Zhou, P.; Xia, Y.; Zhang, J.; Zhang, M.; Zhu, F. Ancestral retrovirus envelope protein ERVWE1 upregulates circ_0001810, a potential biomarker for schizophrenia, and induces neuronal mitochondrial dysfunction via activating AK2. Cell Biosci. 2024, 14, 138. [Google Scholar] [CrossRef] [PubMed]
- Bo, M.; Carta, A.; Cipriani, C.; Cavassa, V.; Simula, E.R.; Huyen, N.T.; Phan, G.T.H.; Noli, M.; Matteucci, C.; Sotgiu, S. HERVs Endophenotype in Autism Spectrum Disorder: Human Endogenous Retroviruses, Specific Immunoreactivity, and Disease Association in Different Family Members. Microorganisms 2024, 13, 9. [Google Scholar] [CrossRef]
- Ruberto, S.; Domınguez-Mozo, M.I.; Garcıa-Martınez, M.A.; Cossu, D.; Sechi, L.A.; Alvarez-Lafuente, R. Immune response profiling of HERV-W envelope proteins in multiple sclerosis: Potential biomarkers for disease progression. Front. Immunol. 2025, 15, 1505239. [Google Scholar] [CrossRef]
- Simula, E.R.; Jasemi, S.; Paulus, K.; Sechi, L.A. Upregulation of microRNAs correlates with downregulation of HERV-K expression in Parkinson’s disease. J. Neurovirol. 2024, 30, 550–555. [Google Scholar] [CrossRef]
- Simula, E.R.; Arru, G.; Zarbo, I.R.; Solla, P.; Sechi, L.A. TDP-43 and HERV-K envelope-specific immunogenic epitopes are recognized in ALS patients. Viruses 2021, 13, 2301. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Montojo, M.; Simula, E.R.; Fathi, S.; McMahan, C.; Ghosal, A.; Berry, J.D.; Cudkowicz, M.; Elkahloun, A.; Johnson, K.; Norato, G. Antibody response to HML-2 may be protective in amyotrophic lateral sclerosis. Ann. Neurol. 2022, 92, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Levet, S.; Charvet, B.; Bertin, A.; Deschaumes, A.; Perron, H.; Hober, D. Human endogenous retroviruses and type 1 diabetes. Curr. Diabetes Rev. 2019, 19, 141. [Google Scholar] [CrossRef] [PubMed]
- Dow, C.T.; Pierce, E.S.; Sechi, L.A. Mycobacterium paratuberculosis: A HERV Turn-On for Autoimmunity, Neurodegeneration, and Cancer? Microorganisms 2024, 12, 1890. [Google Scholar] [CrossRef]
- Gao, Y.; Yu, X.-F.; Chen, T. Human endogenous retroviruses in cancer: Expression, regulation and function. Oncol. Lett. 2021, 21, 121. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Wen, X.; Xing, X.; Fozza, C.; Sechi, L.A. Endogenous retroviruses Suppressyn and Syncytin-2 as innovative prognostic biomarkers in Acute Myeloid Leukemia. Front. Cell. Infect. Microbiol. 2024, 13, 1339673. [Google Scholar] [CrossRef]
- Steiner, M.C.; Marston, J.L.; Iñiguez, L.P.; Bendall, M.L.; Chiappinelli, K.B.; Nixon, D.F.; Crandall, K.A. Locus-specific characterization of human endogenous retrovirus expression in prostate, breast, and colon cancers. Cancer Res. J. 2021, 81, 3449–3460. [Google Scholar] [CrossRef]
- Turcanova, V.L.; Bundgaard, B.; Höllsberg, P. Human herpesvirus-6B induces expression of the human endogenous retrovirus K18-encoded superantigen. J. Clin. Virol. 2009, 46, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Kwun, H.J.; Kim, H.S.; Jang, K.L. Activation of the human endogenous retrovirus W long terminal repeat by herpes simplex virus type 1 immediate early protein 1. Mol. Cells 2003, 15, 75–80. [Google Scholar] [CrossRef]
- Liu, C.; Liu, L.; Wang, X.; Liu, Y.; Wang, M.; Zhu, F. HBV X Protein induces overexpression of HERV-W env through NF-κB in HepG2 cells. Virus Genes 2017, 53, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, Y.; Li, S.; Yu, H.; Zeng, J.; Wang, X.; Zhu, F. Activation of elements in HERV-W family by caffeine and aspirin. Virus Genes 2013, 47, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, U.; Steidler, A.; Trojan, L.; Michel, M.S.; Seifarth, W.; Fabarius, A. Smoking increases transcription of human endogenous retroviruses in a newly established in vitro cell model and in normal urothelium. AIDS Res. Hum. Retrovir. 2010, 26, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.F.; Saraph, A.; Tokuyama, M. Transactivation of Human Endogenous Retroviruses by Viruses. Viruses 2024, 16, 1649. [Google Scholar] [CrossRef] [PubMed]
- Nellåker, C.; Yao, Y.; Jones-Brando, L.; Mallet, F.; Yolken, R.H.; Karlsson, H. Transactivation of elements in the human endogenous retrovirus W family by viral infection. Retrovirology 2006, 3, 44. [Google Scholar] [CrossRef]
- Yu, H.; Liu, T.; Zhao, Z.; Chen, Y.; Zeng, J.; Liu, S.; Zhu, F. Mutations in 3′-long terminal repeat of HERV-W family in chromosome 7 upregulate syncytin-1 expression in urothelial cell carcinoma of the bladder through interacting with c-Myb. Oncogene 2014, 33, 3947–3958. [Google Scholar] [CrossRef]
- Jasemi, S.; Paulus, K.; Noli, M.; Simula, E.R.; Ruberto, S.; Sechi, L.A. Antibodies against HSV-1 and curli show the highest correlation in Parkinson’s disease patients in comparison to healthy controls. Int. J. Mol. Sci. 2022, 23, 14816. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Adler, G.L.; Youssef, P.; Phan, K.; Halliday, G.M.; Dzamko, N.; Kim, W.S. Human Endogenous Retrovirus K in Astrocytes Is Altered in Parkinson’s Disease. Mov. Disord. 2025. [Google Scholar] [CrossRef]
- Jasemi, S.; Simula, E.R.; Pantaleo, A.; Sechi, L.A. Transcriptional upregulation of HERV-env genes under simulated microgravity. Viruses 2025, 17, 306. [Google Scholar] [CrossRef]
- Jasemi, S.; Simula, E.R.; Yasushi, K.; Sechi, L.A. Unveiling the impact of simulated microgravity on HSV-1 infection, neuroinflammation, and endogenous retroviral activation in SH-SY5Y cells. J. Neurovirol. 2025, 1–9. [Google Scholar] [CrossRef]
- Hosseiniporgham, S.; Sechi, L.A. Anti-HERV-K drugs and vaccines, possible therapies against tumors. Vaccines 2023, 11, 751. [Google Scholar] [CrossRef]
- Tavakolian, S.; Goudarzi, H.; Faghihloo, E. Evaluating the expression level of HERV-K env, np9, rec and gag in breast tissue. Infect. Agents Cancer 2019, 14, 42. [Google Scholar] [CrossRef]
- Gomollón, F.; Dignass, A.; Annese, V.; Tilg, H.; Van Assche, G.; Lindsay, J.O.; Peyrin-Biroulet, L.; Cullen, G.J.; Daperno, M.; Kucharzik, T. 3rd European evidence-based consensus on the diagnosis and management of Crohn’s disease 2016: Part 1: Diagnosis and medical management. J. Crohns Colitis 2017, 11, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Dolinger, M.; Torres, J.; Vermeire, S. Crohn’s disease. Lancet 2024, 403, 1177–1191. [Google Scholar] [CrossRef]
- Field, N.L.; McAloon, C.G.; Gavey, L.; Mee, J.F. Mycobacterium avium subspecies paratuberculosis infection in cattle–a review in the context of seasonal pasture-based dairy herds. Ir. Vet. J. 2022, 75, 12. [Google Scholar] [CrossRef] [PubMed]
- O′Connell, L.M.; Coffey, A.; O′Mahony, J.M. Alternatives to antibiotics in veterinary medicine: Considerations for the management of Johne’s disease. Anim. Health Res. Rev. 2023, 24, 12–27. [Google Scholar] [CrossRef]
- Tovo, P.-A.; Ribaldone, D.G.; Galliano, I.; Caviglia, G.P.; Dini, M.; Veglio, V.; Calvi, C.; Montanari, P.; Pitoni, D.; Frara, S. Enhanced transcription of human endogenous retroviruses and TRIM28 downregulation in patients with inflammatory bowel disease. Viruses 2024, 16, 1570. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Z.; Wang, P.; Li, S.; Zeng, J.; Tu, X.; Yan, Q.; Xiao, Z.; Pan, M.; Zhu, F. Syncytin-1, an endogenous retroviral protein, triggers the activation of CRP via TLR3 signal cascade in glial cells. Brain Behavior. Immun. 2018, 67, 324–334. [Google Scholar] [CrossRef]
- Chamouard, P.; Richert, Z.; Meyer, N.; Rahmi, G.; Baumann, R. Diagnostic value of C-reactive protein for predicting activity level of Crohn’s disease. Clin. Gastroenterol. Hepatol. 2006, 4, 882–887. [Google Scholar] [CrossRef]
- Klag, T.; Courth, L.; Ostaff, M.J.; Ott, G.; Stange, E.F.; Malek, N.P.; Seifarth, W.; Wehkamp, J. Human endogenous retroviruses: Residues of ancient times are differentially expressed in Crohn’s disease. Inflamm. Intest. Dis. 2019, 3, 125–137. [Google Scholar] [CrossRef]
- Wang, X.; Wu, X.; Huang, J.; Li, H.; Yan, Q.; Zhu, F. Human endogenous retrovirus W family envelope protein (HERV-W env) facilitates the production of TNF-α and IL-10 by inhibiting MyD88s in glial cells. Arch. Virol. 2021, 166, 1035–1045. [Google Scholar] [CrossRef]
- Dotlacil, V.; Coufal, S.; Lerchova, T.; Zarubova, K.; Kucerova, B.; Tlaskalova-Hogenova, H.; Kverka, M.; Skaba, R.; Bronsky, J.; Hradsky, O. Intestinal tissue levels of anti-TNF alpha, antibodies, and cytokines in paediatric Crohn disease. Sci. Rep. 2025, 15, 1138. [Google Scholar] [CrossRef]
- Derkx, B.; Taminiau, J.; Radema, S.; Stronkhorst, A.; Wortel, C.; Tytgat, G.; van Deventer, S. Tumour-necrosis-factor antibody treatment in Crohn’s disease. Lancet 1993, 342, 173–174. [Google Scholar] [CrossRef] [PubMed]
- Spoettl, T.; Hausmann, M.; Klebl, F.; Dirmeier, A.; Klump, B.; Hoffmann, J.; Herfarth, H.; Timmer, A.; Rogler, G. Serum soluble TNF receptor I and II levels correlate with disease activity in IBD patients. Inflamm. Bowel Dis. 2007, 13, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Liu, Y.; Ma, X.; Liu, L.; Li, S.; Li, R.; Wang, T.; Song, H.; Niu, D. Disrupting endogenous retroelements with a reverse transcriptase inhibitor alleviates DSS-induced colitis in mice. Mucosal Immunol. 2024, 17, 54–66. [Google Scholar] [CrossRef]
- Collins, M.T.; Lisby, G.; Moser, C.; Chicks, D.; Christensen, S.; Reichelderfer, M.; Høiby, N.; Harms, B.A.; Thomsen, O.Ø.; Skibsted, U. Results of multiple diagnostic tests for Mycobacterium avium subsp. paratuberculosis in patients with inflammatory bowel disease and in controls. J. Clin. Microbiol. 2000, 38, 4373–4381. [Google Scholar] [CrossRef] [PubMed]
- Valheim, M.; Sigurðardóttir, Ó.; Storset, A.; Aune, L.; Press, C.M. Characterization of macrophages and occurrence of T cells in intestinal lesions of subclinical paratuberculosis in goats. J. Comp. Pathol. 2004, 131, 221–232. [Google Scholar] [CrossRef]
- Niegowska, M.; Wajda-Cuszlag, M.; Stępień-Ptak, G.; Trojanek, J.; Michałkiewicz, J.; Szalecki, M.; Sechi, L.A. Anti-HERV-WEnv antibodies are correlated with seroreactivity against Mycobacterium avium subsp. paratuberculosis in children and youths at T1D risk. Sci. Rep. 2019, 9, 6282. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides | CD | HC | ||||||
|---|---|---|---|---|---|---|---|---|
| Samples (n) | Median Age | M | F | Samples (n) | Median Age | M | F | |
| HERV-K | 70 | 40.5 | 36 | 34 | 70 | 42 | 37 | 33 |
| HERV-W | 64 | 40 | 35 | 29 | 64 | 41 | 35 | 29 |
| MAP 38-65c | 59 | 41 | 31 | 28 | 59 | 43 | 32 | 27 |
| MAP 1,4-α-gbp | 53 | 39 | 25 | 28 | 53 | 39 | 26 | 27 |
| MAP 2404c | 53 | 39 | 25 | 28 | 53 | 39 | 26 | 27 |
| Epitopes | Amino Acid Sequence | Position |
|---|---|---|
| HERV-K | VWVPGPTDDRCPAKPEEEG | 19–37 |
| HERV-W | NSQCIRWVTPPTQIV | 248–262 |
| MAP 38-65c | MIAVALAGLAANFV | 125–138 |
| MAP 1,4-α-gbp | GTVELLGGPLAHPFQPL | 157–173 |
| MAP 2404c | RGFFYTPKTRREAEDL | 70–85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fayyaz, A.; Cugia, L.; Noli, M.; Jasemi, S.; Simula, E.R.; Sechi, L.A. A Characterization of the Humoral Immune Response to Human Endogenous Retroviruses and Mycobacterium paratuberculosis in Crohn’s Disease. Pathogens 2025, 14, 361. https://doi.org/10.3390/pathogens14040361
Fayyaz A, Cugia L, Noli M, Jasemi S, Simula ER, Sechi LA. A Characterization of the Humoral Immune Response to Human Endogenous Retroviruses and Mycobacterium paratuberculosis in Crohn’s Disease. Pathogens. 2025; 14(4):361. https://doi.org/10.3390/pathogens14040361
Chicago/Turabian StyleFayyaz, Alishba, Luigi Cugia, Marta Noli, Somaye Jasemi, Elena Rita Simula, and Leonardo A. Sechi. 2025. "A Characterization of the Humoral Immune Response to Human Endogenous Retroviruses and Mycobacterium paratuberculosis in Crohn’s Disease" Pathogens 14, no. 4: 361. https://doi.org/10.3390/pathogens14040361
APA StyleFayyaz, A., Cugia, L., Noli, M., Jasemi, S., Simula, E. R., & Sechi, L. A. (2025). A Characterization of the Humoral Immune Response to Human Endogenous Retroviruses and Mycobacterium paratuberculosis in Crohn’s Disease. Pathogens, 14(4), 361. https://doi.org/10.3390/pathogens14040361