
Why Are Cytomegalovirus-Encoded G-Protein-Coupled Receptors Essential for Infection but Only Variably Conserved?


Abstract
1. Introduction
2. Conservation and Diversity of Cytomegalovirus GPCRs
3. Human Cytomegalovirus UL33 and Its Homologues
3.1. UL33 and Its Homologues Facilitate the Infection of Specific Cell Types and Organs, Which Vary Between Model Organisms
3.2. UL33 and Its Homologues Also Signal Constitutively
3.3. UL33 and Its Homologues—Questions Yet to Be Answered
4. Human Cytomegalovirus UL78 and Its Homologues
5. Human Cytomegalovirus US27 and Its Homologues
6. Human Cytomegalovirus US28 and Its Homologues
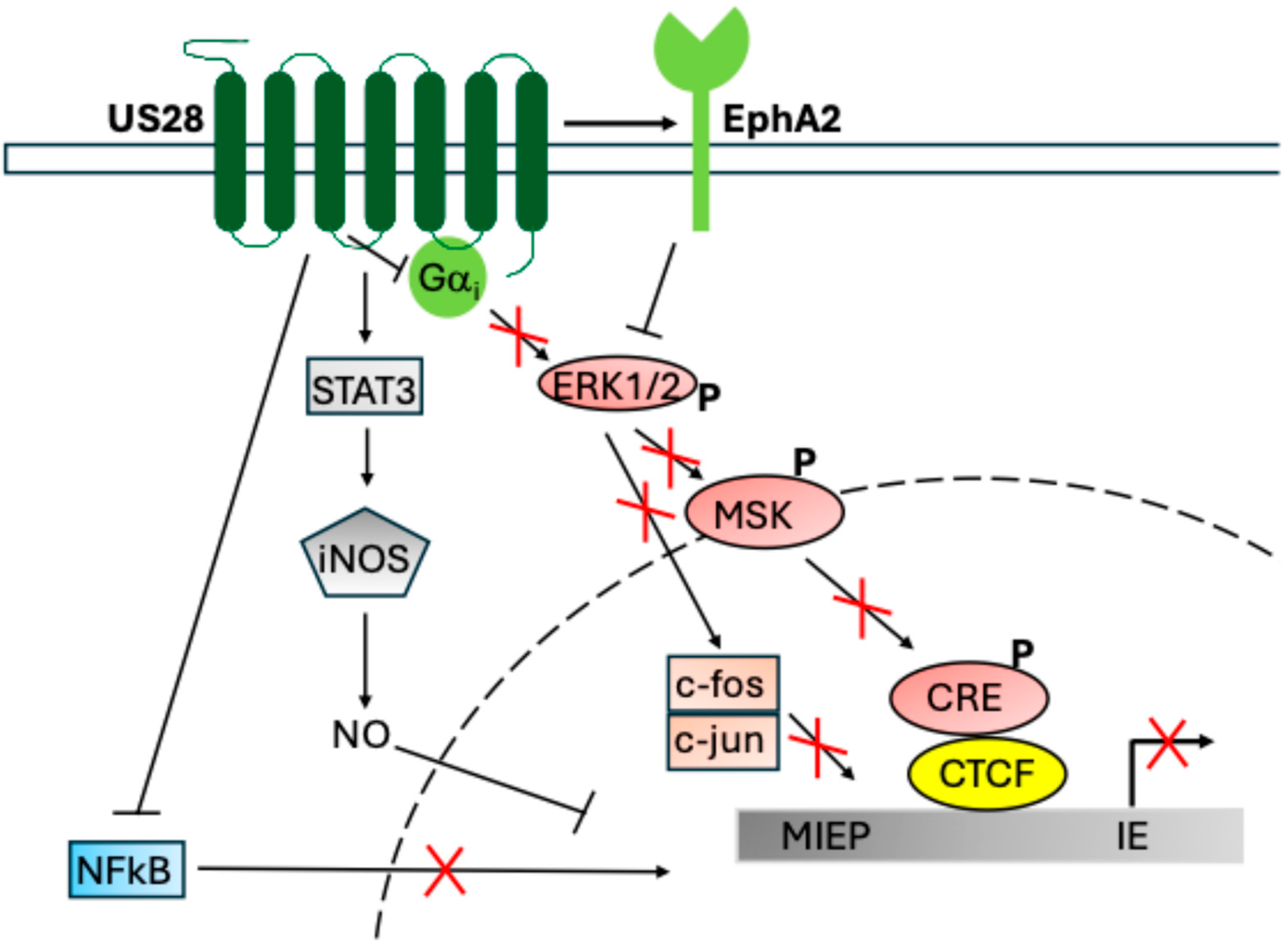
7. US28 and Latent Infection
US28 Homologues in Primate CMVs
8. Functions Derived from CMV-Encoded GPCRs in the Virion
9. Inhibitors and Drugs Targeting GPCRs as Possible Therapeutic Options

{kind=link}
{kind=link}
{kind=link}
| Small Molecules Discovered to Act on US28 | ||||
| Type | Target | Mechanism | References | |
| VUF2274 | Antagonist and inverse agonist | CCR1 CCL5 | Inhibits the PLC-β signalling pathway by sterically blocking chemokine binding and lowering the US28 constitutive signalling activity | [17,136,151,152] |
| Methiothepin/octoclothepin | Antagonist or partial inverse agonist | CCL5 | Blocks CCL5 and inhibits US28 constitutive activity | [16,88,141,153,154] |
| Hydroisoquinoline-based | Antagonist | CX3CR1 | Inhibits US28 constitutive activity (p42/p44 mitogen-activated protein kinase (MAPK) and p38 MAPK-dependent pathways) | [153] |
| Flavonoids | Inverse agonist | --- | Inhibit US28 constitutive activity | [155] |
| Biphenyl amide | Inverse agonist | --- | Inhibit US28 constitutive activity | [156] |
| Macromolecules Designed to Target US28 (Human Cytomegalovirus) | ||||
| Type | Mechanism | References | ||
| F49A-FTP | Fusion toxin protein (FTP) | A modified CX3CL1 which binds US28 with high affinity and targets PE to kill infected cells | [140,148,149] | |
| Bivalent nanobody | Inverse agonist | Displaces CCL5 binding to US28 and prevents constitutive activation of NF-κB and inositol triphosphate (IP3) accumulation. Impairs US28-enhanced tumour growth in vitro and in vivo | [145,157] | |
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jurak, I.; Brune, W. Induction of apoptosis limits cytomegalovirus cross-species infection. EMBO J. 2006, 25, 2634–2642. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Maul, G.G. Mouse cytomegalovirus crosses the species barrier with help from a few human cytomegalovirus proteins. J. Virol. 2006, 80, 7510–7521. [Google Scholar] [CrossRef] [PubMed]
- Burwitz, B.J.; Malouli, D.; Bimber, B.N.; Reed, J.S.; Ventura, A.B.; Hancock, M.H.; Uebelhoer, L.S.; Bhusari, A.; Hammond, K.B.; Espinosa Trethewy, R.G. Cross-species rhesus cytomegalovirus infection of cynomolgus macaques. PLoS Pathog. 2016, 12, e1006014. [Google Scholar] [CrossRef] [PubMed]
- McSharry, B.P.; Avdic, S.; Slobedman, B. Human cytomegalovirus encoded homologs of cytokines, chemokines and their receptors: Roles in immunomodulation. Viruses 2012, 4, 2448–2470. [Google Scholar] [CrossRef]
- Fanunza, E.; Cheng, A.Z.; Auerbach, A.A.; Stefanovska, B.; Moraes, S.N.; Lokensgard, J.R.; Biolatti, M.; Dell’Oste, V.; Bierle, C.J.; Bresnahan, W.A. Human cytomegalovirus mediates APOBEC3B relocalization early during infection through a ribonucleotide reductase-independent mechanism. J. Virol. 2023, 97, e0078123. [Google Scholar] [CrossRef]
- Reyburn, H.T.; Mandelboim, O.; Valés-Gómez, M.; Davis, D.M.; Pazmany, L.; Strominger, J.L. The class I MHC homologue of human cytomegalovirus inhibits attack by natural killer cells. Nature 1997, 386, 514–517. [Google Scholar] [CrossRef]
- Geppetti, P.; Veldhuis, N.A.; Lieu, T.; Bunnett, N.W. G protein-coupled receptors: Dynamic machines for signaling pain and itch. Neuron 2015, 88, 635–649. [Google Scholar] [CrossRef]
- Zhou, Q.; Yang, D.; Wu, M.; Guo, Y.; Guo, W.; Zhong, L.; Cai, X.; Dai, A.; Jang, W.; Shakhnovich, E.I. Common activation mechanism of class A GPCRs. eLife 2019, 8, e50279. [Google Scholar] [CrossRef]
- Bachelerie, F.; Graham, G.J.; Locati, M.; Mantovani, A.; Murphy, P.M.; Nibbs, R.; Rot, A.; Sozzani, S.; Thelen, M. New nomenclature for atypical chemokine receptors. Nat. Immunol. 2014, 15, 207–208. [Google Scholar] [CrossRef]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef]
- O’Connor, C.; Adams, J. 4.2 G-Protein-Coupled Receptors Play Many Different Roles in Eukaryotic Cell Signaling. In Essentials of Cell Biology; NPG Education: Cambridge, MA, USA, 2010. [Google Scholar]
- Rosenkilde, M.M.; Tsutsumi, N.; Knerr, J.M.; Kildedal, D.F.; Garcia, K.C. Viral G Protein–Coupled Receptors Encoded by β- and γ-Herpesviruses. Annu. Rev. Virol. 2022, 9, 329–351. [Google Scholar] [CrossRef] [PubMed]
- Chee, M.S.; Satchwell, S.C.; Preddie, E.; Weston, K.M.; Barrell, B.G. Human cytomegalovirus encodes three G protein-coupled receptor homologues. Nature 1990, 344, 774–777. [Google Scholar] [CrossRef]
- Holst, P.J.; Lüttichau, H.R.; Schwartz, T.W.; Rosenkilde, M.M. Virally encoded chemokines and chemokine receptors in the role of viral infections. Host Response Mech. Infect. Dis. 2003, 10, 232–252. [Google Scholar]
- Randolph-Habecker, J.; Rahill, B.; Torok-Storb, B.; Vieira, J.; Kolattukudy, P.E.; Rovin, B.H.; Sedmak, D.D. The expression of the cytomegalovirus chemokine receptor homolog US28 sequesters biologically active CC chemokines and alters IL-8 production. Cytokine 2002, 19, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Vischer, H.F.; Siderius, M.; Leurs, R.; Smit, M.J. Herpesvirus-encoded GPCRs: Neglected players in inflammatory and proliferative diseases? Nat. Rev. Drug Discov. 2014, 13, 123–139. [Google Scholar] [CrossRef]
- Casarosa, P.; Gruijthuijsen, Y.K.; Michel, D.; Beisser, P.S.; Holl, J.; Fitzsimons, C.P.; Verzijl, D.; Bruggeman, C.A.; Mertens, T.; Leurs, R.; et al. Constitutive signaling of the human cytomegalovirus-encoded receptor UL33 differs from that of its rat cytomegalovirus homolog R33 by promiscuous activation of G proteins of the Gq, Gi, and Gs classes. J. Biol. Chem. 2003, 278, 50010–50023. [Google Scholar] [CrossRef]
- Waldhoer, M.; Kledal, T.N.; Farrell, H.; Schwartz, T.W. Murine Cytomegalovirus (CMV) M33 and Human CMV US28 Receptors Exhibit Similar Constitutive Signaling Activities. J. Virol. 2002, 76, 8161–8168. [Google Scholar] [CrossRef]
- Bittencourt, F.M.; Wu, S.-E.; Bridges, J.P.; Miller, W.E. The M33 G protein-coupled receptor encoded by murine cytomegalovirus is dispensable for hematogenous dissemination but is required for growth within the salivary gland. J. Virol. 2014, 88, 11811–11824. [Google Scholar] [CrossRef]
- Cekinovic, D.; Lisnic, V.; Jonjic, S.; Yurochko, A.; Miller, W. Human Cytomegaloviruses Methods and Protocols; Springer Nature: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Scalzo, A.A.; Corbett, A.J.; Rawlinson, W.D.; Scott, G.M.; Degli-Esposti, M.A. The interplay between host and viral factors in shaping the outcome of cytomegalovirus infection. Immunol. Cell Biol. 2007, 85, 46–54. [Google Scholar] [CrossRef]
- Sacher, T.; Mohr, C.A.; Weyn, A.; Schlichting, C.; Koszinowski, U.H.; Ruzsics, Z. The role of cell types in cytomegalovirus infection in vivo. Eur. J. Cell Biol. 2012, 91, 70–77. [Google Scholar] [CrossRef]
- Bale Jr, J.F.; O’Neil, M.E. Detection of murine cytomegalovirus DNA in circulating leukocytes harvested during acute infection of mice. J. Virol. 1989, 63, 2667–2673. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.M.; Quirk, M.R.; Jordan, M.C. Biphasic viremia and viral gene expression in leukocytes during acute cytomegalovirus infection of mice. J. Virol. 1994, 68, 6305–6311. [Google Scholar] [CrossRef] [PubMed]
- Henson, D.; Strano, A.J. Mouse cytomegalovirus: Necrosis of infected and morphologically normal submaxillary gland acinar cells during termination of chronic infection. Am. J. Pathol. 1972, 68, 183. [Google Scholar] [PubMed]
- Humphreys, I.R.; De Trez, C.; Kinkade, A.; Benedict, C.A.; Croft, M.; Ware, C.F. Cytomegalovirus exploits IL-10–mediated immune regulation in the salivary glands. J. Exp. Med. 2007, 204, 1217–1225. [Google Scholar] [CrossRef]
- Campbell, A.E.; Cavanaugh, V.J.; Slater, J.S. The salivary glands as a privileged site of cytomegalovirus immune evasion and persistence. Med. Microbiol. Immunol. 2008, 197, 205–213. [Google Scholar] [CrossRef]
- Jonjić, S.; Pavić, I.; Lucin, P.; Rukavina, D.; Koszinowski, U.H. Efficacious control of cytomegalovirus infection after long-term depletion of CD8+ T lymphocytes. J. Virol. 1990, 64, 5457–5464. [Google Scholar] [CrossRef]
- Walton, S.M.; Mandaric, S.; Torti, N.; Zimmermann, A.; Hengel, H.; Oxenius, A. Absence of cross-presenting cells in the salivary gland and viral immune evasion confine cytomegalovirus immune control to effector CD4 T cells. PLoS Pathog. 2011, 7, e1002214. [Google Scholar] [CrossRef]
- Roark, H.K.; Jenks, J.A.; Permar, S.R.; Schleiss, M.R. Animal models of congenital cytomegalovirus transmission: Implications for vaccine development. J. Infect. Dis. 2020, 221, S60–S73. [Google Scholar] [CrossRef]
- Vink, C.; Beuken, E.; Bruggeman, C.A. Complete DNA sequence of the rat cytomegalovirus genome. J. Virol. 2000, 74, 7656–7665. [Google Scholar] [CrossRef]
- Weekes, M.P.; Tomasec, P.; Huttlin, E.L.; Fielding, C.A.; Nusinow, D.; Stanton, R.J.; Wang, E.C.; Aicheler, R.; Murrell, I.; Wilkinson, G.W. Quantitative temporal viromics: An approach to investigate host-pathogen interaction. Cell 2014, 157, 1460–1472. [Google Scholar] [CrossRef]
- van Senten, J.R.; Bebelman, M.P.; van Gasselt, P.; Bergkamp, N.D.; van den Bor, J.; Siderius, M.; Smit, M.J. Human Cytomegalovirus-Encoded G Protein-Coupled Receptor UL33 Facilitates Virus Dissemination via the Extracellular and Cell-to-Cell Route. Viruses 2020, 12, 594. [Google Scholar] [CrossRef] [PubMed]
- Krishna, B.A.; Wass, A.B.; Dooley, A.L.; O’Connor, C.M. CMV-encoded GPCR pUL33 activates CREB and facilitates its recruitment to the MIE locus for efficient viral reactivation. J. Cell Sci. 2021, 134, jcs254268. [Google Scholar] [CrossRef] [PubMed]
- Margulies, B.J.; Browne, H.; Gibson, W. Identification of the human cytomegalovirus G protein-coupled receptor homologue encoded by UL33 in infected cells and enveloped virus particles. Virology 1996, 225, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, G.W.; Davison, A.J.; Tomasec, P.; Fielding, C.A.; Aicheler, R.; Murrell, I.; Seirafian, S.; Wang, E.C.; Weekes, M.; Lehner, P.J. Human cytomegalovirus: Taking the strain. Med. Microbiol. Immunol. 2015, 204, 273–284. [Google Scholar] [CrossRef]
- Freeman, M.R.; Dooley, A.L.; Beucler, M.J.; Sanders, W.; Moorman, N.J.; O’Connor, C.M.; Miller, W.E. The Human Cytomegalovirus vGPCR UL33 is Essential for Efficient Lytic Replication in Epithelial Cells. bioRxiv 2024. [Google Scholar] [CrossRef]
- Beisser, P.S.; Vink, C.; Van Dam, J.G.; Grauls, G.; Vanherle, S.J.; Bruggeman, C.A. The R33 G protein-coupled receptor gene of rat cytomegalovirus plays an essential role in the pathogenesis of viral infection. J. Virol. 1998, 72, 2352–2363. [Google Scholar] [CrossRef]
- Kaptein, S.J.; Beisser, P.S.; Gruijthuijsen, Y.K.; Savelkouls, K.G.; van Cleef, K.W.; Beuken, E.; Grauls, G.E.; Bruggeman, C.A.; Vink, C. The rat cytomegalovirus R78 G protein-coupled receptor gene is required for production of infectious virus in the spleen. J. Gen. Virol. 2003, 84, 2517–2530. [Google Scholar] [CrossRef]
- Farrell, H.E.; Abraham, A.M.; Cardin, R.D.; Sparre-Ulrich, A.H.; Rosenkilde, M.M.; Spiess, K.; Jensen, T.H.; Kledal, T.N.; Davis-Poynter, N. Partial functional complementation between human and mouse cytomegalovirus chemokine receptor homologues. J. Virol. 2011, 85, 6091–6095. [Google Scholar] [CrossRef]
- Davis-Poynter, N.J.; Lynch, D.M.; Vally, H.; Shellam, G.R.; Rawlinson, W.D.; Barrell, B.G.; Farrell, H.E. Identification and characterization of a G protein-coupled receptor homolog encoded by murine cytomegalovirus. J. Virol. 1997, 71, 1521–1529. [Google Scholar] [CrossRef]
- Gruijthuijsen, Y.K.; Casarosa, P.; Kaptein, S.J.; Broers, J.L.; Leurs, R.; Bruggeman, C.A.; Smit, M.J.; Vink, C. The rat cytomegalovirus R33-encoded G protein-coupled receptor signals in a constitutive fashion. J. Virol. 2002, 76, 1328–1338. [Google Scholar] [CrossRef]
- Bonavita, C.M.; White, T.M.; Francis, J.; Farrell, H.E.; Davis-Poynter, N.J.; Cardin, R.D. The viral G-protein-coupled receptor homologs M33 and US28 promote cardiac dysfunction during murine cytomegalovirus infection. Viruses 2023, 15, 711. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Watanabe, S.; Katano, H.; Noguchi, K.; Sato, Y.; Kojima, S.; Miura, T.; Majima, R.; Yamada, S.; Inoue, N. Roles of GP33, a guinea pig cytomegalovirus-encoded G protein-coupled receptor homolog, in cellular signaling, viral growth and inflammation in vitro and in vivo. PLoS Pathog. 2018, 14, e1007487. [Google Scholar] [CrossRef] [PubMed]
- Deckers, M.; Hofmann, J.; Kreuzer, K.-A.; Reinhard, H.; Edubio, A.; Hengel, H.; Voigt, S.; Ehlers, B. High genotypic diversity and a novel variant of human cytomegalovirus revealed by combined UL33/UL55 genotyping with broad-range PCR. Virol. J. 2009, 6, 210. [Google Scholar] [CrossRef] [PubMed]
- Tschische, P.; Tadagaki, K.; Kamal, M.; Jockers, R.; Waldhoer, M. Heteromerization of human cytomegalovirus encoded chemokine receptors. Biochem. Pharmacol. 2011, 82, 610–619. [Google Scholar] [CrossRef]
- Van Senten, J.R.; Bebelman, M.P.; Fan, T.S.; Heukers, R.; Bergkamp, N.D.; Van Gasselt, P.; Langemeijer, E.V.; Slinger, E.; Lagerweij, T.; Rahbar, A. The human cytomegalovirus-encoded G protein–coupled receptor UL33 exhibits oncomodulatory properties. J. Biol. Chem. 2019, 294, 16297–16308. [Google Scholar] [CrossRef]
- Sherrill, J.D.; Stropes, M.P.; Schneider, O.D.; Koch, D.E.; Bittencourt, F.M.; Miller, J.L.; Miller, W.E. Activation of intracellular signaling pathways by the murine cytomegalovirus G protein-coupled receptor M33 occurs via PLC-β/PKC-dependent and-independent mechanisms. J. Virol. 2009, 83, 8141–8152. [Google Scholar] [CrossRef]
- Case, R.; Sharp, E.; Benned-Jensen, T.; Rosenkilde, M.M.; Davis-Poynter, N.; Farrell, H.E. Functional analysis of the murine cytomegalovirus chemokine receptor homologue M33: Ablation of constitutive signaling is associated with an attenuated phenotype in vivo. J. Virol. 2008, 82, 1884–1898. [Google Scholar] [CrossRef]
- Krishna, B.A.; Poole, E.L.; Jackson, S.E.; Smit, M.J.; Wills, M.R.; Sinclair, J.H. Latency-Associated Expression of Human Cytomegalovirus US28 Attenuates Cell Signaling Pathways To Maintain Latent Infection. mBio 2017, 8, e01754-17. [Google Scholar] [CrossRef]
- Michel, D.; Milotić, I.; Wagner, M.; Vaida, B.; Holl, J.; Ansorge, R.; Mertens, T. The human cytomegalovirus UL78 gene is highly conserved among clinical isolates, but is dispensable for replication in fibroblasts and a renal artery organ-culture system. J. Gen. Virol. 2005, 86, 297–306. [Google Scholar] [CrossRef]
- Cheng, S.; Caviness, K.; Buehler, J.; Smithey, M.; Nikolich-Zugich, J.; Goodrum, F. Transcriptome-wide characterization of human cytomegalovirus in natural infection and experimental latency. Proc. Natl. Acad. Sci. USA 2017, 114, E10586–E10595. [Google Scholar] [CrossRef]
- Rossetto, C.C.; Tarrant-Elorza, M.; Pari, G.S. Cis and trans acting factors involved in human cytomegalovirus experimental and natural latent infection of CD14 (+) monocytes and CD34 (+) cells. PLoS Pathog. 2013, 9, e1003366. [Google Scholar] [CrossRef] [PubMed]
- Goodrum, F.D.; Jordan, C.T.; High, K.; Shenk, T. Human cytomegalovirus gene expression during infection of primary hematopoietic progenitor cells: A model for latency. Proc. Natl. Acad. Sci. USA 2002, 99, 16255–16260. [Google Scholar] [CrossRef] [PubMed]
- Gompels, U.; Nicholas, J.; Lawrence, G.; Jones, M.; Thomson, B.; Martin, M.; Efstathiou, S.; Craxton, M.; Macaulay, H. The DNA sequence of human herpesvirus-6: Structure, coding content, and genome evolution. Virology 1995, 209, 29–51. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Y.; Zhou, Q.; Cong, Z.; Lin, S.; Yan, J.; Chen, X.; Yang, D.; Ying, T.; Wang, M.-W. A homotrimeric GPCR architecture of the human cytomegalovirus revealed by cryo-EM. Cell Discov. 2024, 10, 52. [Google Scholar] [CrossRef]
- Tsutsumi, N.; Maeda, S.; Qu, Q.; Vögele, M.; Jude, K.M.; Suomivuori, C.-M.; Panova, O.; Waghray, D.; Kato, H.E.; Velasco, A. Atypical structural snapshots of human cytomegalovirus GPCR interactions with host G proteins. Sci. Adv. 2022, 8, eabl5442. [Google Scholar] [CrossRef]
- O’Connor, C.M.; Shenk, T. Human cytomegalovirus pUL78 G protein-coupled receptor homologue is required for timely cell entry in epithelial cells but not fibroblasts. J. Virol. 2012, 86, 11425–11433. [Google Scholar] [CrossRef]
- Rovati, G.E.; Capra, V.; Neubig, R.R. The highly conserved DRY motif of class AG protein-coupled receptors: Beyond the ground state. Mol. Pharmacol. 2007, 71, 959–964. [Google Scholar] [CrossRef]
- Tadagaki, K.; Tudor, D.; Gbahou, F.; Tschische, P.; Waldhoer, M.; Bomsel, M.; Jockers, R.; Kamal, M. Human cytomegalovirus-encoded UL33 and UL78 heteromerize with host CCR5 and CXCR4 impairing their HIV coreceptor activity. Blood J. Am. Soc. Hematol. 2012, 119, 4908–4918. [Google Scholar] [CrossRef]
- Wagner, S.; Arnold, F.; Wu, Z.; Schubert, A.; Walliser, C.; Tadagaki, K.; Jockers, R.; Mertens, T.; Michel, D. The 7-transmembrane protein homologue UL78 of the human cytomegalovirus forms oligomers and traffics between the plasma membrane and different intracellular compartments. Arch. Virol. 2012, 157, 935–949. [Google Scholar] [CrossRef]
- Oliveira, S.A.; Shenk, T.E. Murine cytomegalovirus M78 protein, a G protein-coupled receptor homologue, is a constituent of the virion and facilitates accumulation of immediate-early viral mRNA. Proc. Natl. Acad. Sci. USA 2001, 98, 3237–3242. [Google Scholar] [CrossRef]
- Schleiss, M.R.; McGregor, A.; Choi, K.Y.; Date, S.V.; Cui, X.; McVoy, M.A. Analysis of the nucleotide sequence of the guinea pig cytomegalovirus (GPCMV) genome. Virol. J. 2008, 5, 139. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.G.; Hancock, M.H.; Malouli, D.; Marshall, E.E.; Hughes, C.M.; Randall, K.T.; Morrow, D.; Ford, J.C.; Gilbride, R.M.; Selseth, A.N.; et al. Myeloid cell tropism enables MHC-E–restricted CD8+ T cell priming and vaccine efficacy by the RhCMV/SIV vaccine. Sci. Immunol. 2022, 7, eabn9301. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.G.; Zak, D.E.; Xu, G.; Ford, J.C.; Marshall, E.E.; Malouli, D.; Gilbride, R.M.; Hughes, C.M.; Ventura, A.B.; Ainslie, E.; et al. Prevention of tuberculosis in rhesus macaques by a cytomegalovirus-based vaccine. Nat. Med. 2018, 24, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Sharp, E.; Davis-Poynter, N.; Farrell, H. Analysis of the subcellular trafficking properties of murine cytomegalovirus M78, a 7 transmembrane receptor homologue. J. Gen. Virol. 2009, 90, 59–68. [Google Scholar] [CrossRef]
- Davis-Poynter, N.; Yunis, J.; Farrell, H.E. The cytoplasmic C-tail of the mouse cytomegalovirus 7 transmembrane receptor homologue, M78, regulates endocytosis of the receptor and modulates virus replication in different cell types. PLoS ONE 2016, 11, e0165066. [Google Scholar] [CrossRef]
- Yunis, J.; Farrell, H.E.; Bruce, K.; Lawler, C.; Sidenius, S.; Wyer, O.; Davis-Poynter, N.; Stevenson, P.G. Murine cytomegalovirus degrades MHC class II to colonize the salivary glands. PLoS Pathog. 2018, 14, e1006905. [Google Scholar] [CrossRef]
- Margulies, B.J.; Gibson, W. The chemokine receptor homologue encoded by US27 of human cytomegalovirus is heavily glycosylated and is present in infected human foreskin fibroblasts and enveloped virus particles. Virus Res. 2007, 123, 57–71. [Google Scholar] [CrossRef]
- Humby, M.S.; O’Connor, C.M. Human Cytomegalovirus US28 Is Important for Latent Infection of Hematopoietic Progenitor Cells. J. Virol. 2015, 90, 2959–2970. [Google Scholar] [CrossRef]
- Welch, A.R.; McGregor, L.M.; Gibson, W. Cytomegalovirus homologs of cellular G protein-coupled receptor genes are transcribed. J. Virol. 1991, 65, 3915–3918. [Google Scholar] [CrossRef]
- Balázs, Z.; Tombácz, D.; Szűcs, A.; Csabai, Z.; Megyeri, K.; Petrov, A.N.; Snyder, M.; Boldogkői, Z. Long-Read Sequencing of Human Cytomegalovirus Transcriptome Reveals RNA Isoforms Carrying Distinct Coding Potentials. Sci. Rep. 2017, 7, 15989. [Google Scholar] [CrossRef]
- Arnolds, K.L.; Lares, A.P.; Spencer, J.V. The US27 gene product of human cytomegalovirus enhances signaling of host chemokine receptor CXCR4. Virology 2013, 439, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Stegman, J.R.; Margulies, B.J. The human cytomegalovirus chemokine receptor homolog encoded by US27. Virus Genes 2017, 53, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Boeck, J.M.; Stowell, G.A.; O’Connor, C.M.; Spencer, J.V. The human cytomegalovirus US27 gene product constitutively activates antioxidant response element-mediated transcription through Gβγ, phosphoinositide 3-kinase, and nuclear respiratory factor 1. J. Virol. 2018, 92, e02185-18. [Google Scholar] [CrossRef]
- O’Connor, C.M.; Shenk, T. Human cytomegalovirus pUS27 G protein-coupled receptor homologue is required for efficient spread by the extracellular route but not for direct cell-to-cell spread. J. Virol. 2011, 85, 3700–3707. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.; Chou, C.; Li, H.; Hai, R.; Patterson, D.; Stolc, V.; Zhu, H.; Liu, F. Functional profiling of a human cytomegalovirus genome. Proc. Natl. Acad. Sci. USA 2003, 100, 14223–14228. [Google Scholar] [CrossRef]
- Boeck, J.M.; Spencer, J.V. Effect of human cytomegalovirus (HCMV) US27 on CXCR4 receptor internalization measured by fluorogen-activating protein (FAP) biosensors. PLoS ONE 2017, 12, e0172042. [Google Scholar] [CrossRef]
- Frank, T.; Reichel, A.; Larsen, O.; Stilp, A.-C.; Rosenkilde, M.M.; Stamminger, T.; Ozawa, T.; Tschammer, N. Attenuation of chemokine receptor function and surface expression as an immunomodulatory strategy employed by human cytomegalovirus is linked to vGPCR US28. Cell Commun. Signal. 2016, 14, 31. [Google Scholar] [CrossRef]
- Bianchi, M.E.; Mezzapelle, R. The chemokine receptor CXCR4 in cell proliferation and tissue regeneration. Front. Immunol. 2020, 11, 2109. [Google Scholar] [CrossRef]
- Gao, A.; Lin, Y.; Chai, Y.; Han, J.; Wu, L.; Ye, J. CXCL12/CXCR4 Axis promotes the chemotaxis and phagocytosis of B cells through the PI3K-AKT signaling pathway in an early vertebrate. J. Immunol. 2024, 213, 1676–1690. [Google Scholar] [CrossRef]
- Singh, P.; Mohammad, K.S.; Pelus, L.M. CXCR4 expression in the bone marrow microenvironment is required for hematopoietic stem and progenitor cell maintenance and early hematopoietic regeneration after myeloablation. Stem Cells 2020, 38, 849–859. [Google Scholar] [CrossRef]
- Sijmons, S.; Thys, K.; Mbong Ngwese, M.; Van Damme, E.; Dvorak, J.; Van Loock, M.; Li, G.; Tachezy, R.; Busson, L.; Aerssens, J. High-throughput analysis of human cytomegalovirus genome diversity highlights the widespread occurrence of gene-disrupting mutations and pervasive recombination. J. Virol. 2015, 89, 7673–7695. [Google Scholar] [CrossRef] [PubMed]
- Burg, J.S.; Ingram, J.R.; Venkatakrishnan, A.; Jude, K.M.; Dukkipati, A.; Feinberg, E.N.; Angelini, A.; Waghray, D.; Dror, R.O.; Ploegh, H.L. Structural basis for chemokine recognition and activation of a viral G protein–coupled receptor. Science 2015, 347, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.E.; Zagorski, W.A.; Brenneman, J.D.; Avery, D.; Miller, J.L.; O’Connor, C.M. US28 is a potent activator of phospholipase C during HCMV infection of clinically relevant target cells. PLoS ONE 2012, 7, e50524. [Google Scholar] [CrossRef] [PubMed]
- Krishna, B.A.; Miller, W.E.; O’Connor, C.M. US28: HCMV’s Swiss Army knife. Viruses 2018, 10, 445. [Google Scholar] [CrossRef]
- Casarosa, P.; Bakker, R.A.; Verzijl, D.; Navis, M.; Timmerman, H.; Leurs, R.; Smit, M.J. Constitutive signaling of the human cytomegalovirus-encoded chemokine receptor US28. J. Biol. Chem. 2001, 276, 1133–1137. [Google Scholar] [CrossRef]
- Fraile-Ramos, A.; Kledal, T.N.; Pelchen-Matthews, A.; Bowers, K.; Schwartz, T.W.; Marsh, M. The human cytomegalovirus US28 protein is located in endocytic vesicles and undergoes constitutive endocytosis and recycling. Mol. Biol. Cell 2001, 12, 1737–1749. [Google Scholar] [CrossRef]
- Bakker, R.A.; Casarosa, P.; Timmerman, H.; Smit, M.J.; Leurs, R. Constitutively active Gq/11-coupled receptors enable signaling by co-expressed Gi/o-coupled receptors. J. Biol. Chem. 2004, 279, 5152–5161. [Google Scholar] [CrossRef]
- Moepps, B.; Tulone, C.; Kern, C.; Minisini, R.; Michels, G.; Vatter, P.; Wieland, T.; Gierschik, P. Constitutive serum response factor activation by the viral chemokine receptor homologue pUS28 is differentially regulated by Gαq/11 and Gα16. Cell. Signal. 2008, 20, 1528–1537. [Google Scholar] [CrossRef]
- Minisini, R.; Tulone, C.; Lüske, A.; Michel, D.; Mertens, T.; Gierschik, P.; Moepps, B. Constitutive inositol phosphate formation in cytomegalovirus-infected human fibroblasts is due to expression of the chemokine receptor homologue pUS28. J. Virol. 2003, 77, 4489–4501. [Google Scholar] [CrossRef]
- Neote, K.; DiGregorio, D.; Mak, J.Y.; Horuk, R.; Schall, T.J. Molecular cloning, functional expression, and signaling characteristics of a CC chemokine receptor. Cell 1993, 72, 415–425. [Google Scholar] [CrossRef]
- Maussang, D.; Verzijl, D.; Van Walsum, M.; Leurs, R.; Holl, J.; Pleskoff, O.; Michel, D.; Van Dongen, G.A.; Smit, M.J. Human cytomegalovirus-encoded chemokine receptor US28 promotes tumorigenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 13068–13073. [Google Scholar] [CrossRef] [PubMed]
- Maussang, D.; Langemeijer, E.; Fitzsimons, C.P.; Stigter-van Walsum, M.; Dijkman, R.; Borg, M.K.; Slinger, E.; Schreiber, A.; Michel, D.; Tensen, C.P. The human cytomegalovirus–encoded chemokine receptor US28 promotes angiogenesis and tumor formation via cyclooxygenase-2. Cancer Res. 2009, 69, 2861–2869. [Google Scholar] [CrossRef] [PubMed]
- Slinger, E.; Maussang, D.; Schreiber, A.; Siderius, M.; Rahbar, A.; Fraile-Ramos, A.; Lira, S.A.; Söderberg-Nauclér, C.; Smit, M.J. HCMV-encoded chemokine receptor US28 mediates proliferative signaling through the IL-6–STAT3 axis. Sci. Signal. 2010, 3, ra58. [Google Scholar] [CrossRef] [PubMed]
- Langemeijer, E.V.; Slinger, E.; de Munnik, S.; Schreiber, A.; Maussang, D.; Vischer, H.; Verkaar, F.; Leurs, R.; Siderius, M.; Smit, M.J. Constitutive ss-catenin signaling by the viral chemokine receptor US28. PLoS ONE 2012, 7, e48935. [Google Scholar] [CrossRef]
- de Wit, R.H.; Mujić-Delić, A.; van Senten, J.R.; Fraile-Ramos, A.; Siderius, M.; Smit, M.J. Human cytomegalovirus encoded chemokine receptor US28 activates the HIF-1α/PKM2 axis in glioblastoma cells. Oncotarget 2016, 7, 67966. [Google Scholar] [CrossRef]
- Gao, J.-L.; Murphy, P.M. Human cytomegalovirus open reading frame US28 encodes a functional beta chemokine receptor. J. Biol. Chem. 1994, 269, 28539–28542. [Google Scholar] [CrossRef]
- Billstrom, M.A.; Johnson, G.L.; Avdi, N.J.; Worthen, G.S. Intracellular signaling by the chemokine receptor US28 during human cytomegalovirus infection. J. Virol. 1998, 72, 5535–5544. [Google Scholar] [CrossRef]
- Vieira, J.; Schall, T.J.; Corey, L.; Geballe, A.P. Functional analysis of the human cytomegalovirus US28 gene by insertion mutagenesis with the green fluorescent protein gene. J. Virol. 1998, 72, 8158–8165. [Google Scholar] [CrossRef]
- Melnychuk, R.M.; Streblow, D.N.; Smith, P.P.; Hirsch, A.J.; Pancheva, D.; Nelson, J.A. Human cytomegalovirus-encoded G protein-coupled receptor US28 mediates smooth muscle cell migration through Gα12. J. Virol. 2004, 78, 8382–8391. [Google Scholar] [CrossRef]
- Streblow, D.N.; Vomaske, J.; Smith, P.; Melnychuk, R.; Hall, L.; Pancheva, D.; Smit, M.; Casarosa, P.; Schlaepfer, D.D.; Nelson, J.A. Human cytomegalovirus chemokine receptor US28-induced smooth muscle cell migration is mediated by focal adhesion kinase and Src. J. Biol. Chem. 2003, 278, 50456–50465. [Google Scholar] [CrossRef]
- Vomaske, J.; Melnychuk, R.M.; Smith, P.P.; Powell, J.; Hall, L.; DeFilippis, V.; Früh, K.; Smit, M.; Schlaepfer, D.D.; Nelson, J.A. Differential ligand binding to a human cytomegalovirus chemokine receptor determines cell type–specific motility. PLoS Pathog. 2009, 5, e1000304. [Google Scholar] [CrossRef] [PubMed]
- Streblow, D.N.; Soderberg-Naucler, C.; Vieira, J.; Smith, P.; Wakabayashi, E.; Ruchti, F.; Mattison, K.; Altschuler, Y.; Nelson, J.A. The human cytomegalovirus chemokine receptor US28 mediates vascular smooth muscle cell migration. Cell 1999, 99, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Soroceanu, L.; Matlaf, L.; Bezrookove, V.; Harkins, L.; Martinez, R.; Greene, M.; Soteropoulos, P.; Cobbs, C.S. Human cytomegalovirus US28 found in glioblastoma promotes an invasive and angiogenic phenotype. Cancer Res. 2011, 71, 6643–6653. [Google Scholar] [CrossRef] [PubMed]
- Casarosa, P.; Waldhoer, M.; LiWang, P.J.; Vischer, H.F.; Kledal, T.; Timmerman, H.; Schwartz, T.W.; Smit, M.J.; Leurs, R. CC and CX3C chemokines differentially interact with the N terminus of the human cytomegalovirus-encoded US28 receptor. J. Biol. Chem. 2005, 280, 3275–3285. [Google Scholar] [CrossRef]
- Miller, W.E.; Houtz, D.A.; Nelson, C.D.; Kolattukudy, P.; Lefkowitz, R.J. G-protein-coupled receptor (GPCR) kinase phosphorylation and β-arrestin recruitment regulate the constitutive signaling activity of the human cytomegalovirus US28 GPCR. J. Biol. Chem. 2003, 278, 21663–21671. [Google Scholar] [CrossRef]
- Bergkamp, N.D.; van Senten, J.R.; Brink, H.J.; Bebelman, M.P.; van den Bor, J.; Çobanoğlu, T.S.; Dinkla, K.; Köster, J.; Klau, G.; Siderius, M. A virally encoded GPCR drives glioblastoma through feed-forward activation of the SK1-S1P1 signaling axis. Sci. Signal. 2023, 16, eade6737. [Google Scholar] [CrossRef]
- Bebelman, M.P.; Setiawan, I.M.; Bergkamp, N.D.; van Senten, J.R.; Crudden, C.; Bebelman, J.P.M.; Verweij, F.J.; van Niel, G.; Siderius, M.; Pegtel, D.M. Exosomal release of the virus-encoded chemokine receptor US28 contributes to chemokine scavenging. iScience 2023, 26, 107412. [Google Scholar] [CrossRef]
- Crawford, L.B.; Caposio, P.; Kreklywich, C.; Pham, A.H.; Hancock, M.H.; Jones, T.A.; Smith, P.P.; Yurochko, A.D.; Nelson, J.A.; Streblow, D.N. Human cytomegalovirus US28 ligand binding activity is required for latency in CD34+ hematopoietic progenitor cells and humanized NSG mice. mBio 2019, 10, 10-1128. [Google Scholar] [CrossRef]
- Krishna, B.A.; Wass, A.B.; Sridharan, R.; O’Connor, C.M. The Requirement for US28 During Cytomegalovirus Latency Is Independent of US27 and US29 Gene Expression. Front. Cell. Infect. Microbiol. 2020, 10, 186. [Google Scholar] [CrossRef]
- Zhu, D.; Pan, C.; Sheng, J.; Liang, H.; Bian, Z.; Liu, Y.; Trang, P.; Wu, J.; Liu, F.; Zhang, C.-Y.; et al. Human cytomegalovirus reprogrammes haematopoietic progenitor cells into immunosuppressive monocytes to achieve latency. Nat. Microbiol. 2018, 3, 503–513. [Google Scholar] [CrossRef]
- Elder, E.G.; Krishna, B.A.; Williamson, J.; Lim, E.Y.; Poole, E.; Sedikides, G.X.; Wills, M.; O’Connor, C.M.; Lehner, P.J.; Sinclair, J. Interferon-Responsive Genes Are Targeted during the Establishment of Human Cytomegalovirus Latency. mBio 2019, 10, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Elder, E.G.; Krishna, B.A.; Poole, E.; Perera, M.; Sinclair, J. Regulation of host and viral promoters during human cytomegalovirus latency via US28 and CTCF. J. Gen. Virol. 2021, 102, 001609. [Google Scholar] [CrossRef] [PubMed]
- Groves, I.J.; Matthews, S.M.; O’Connor, C.M. Host-encoded CTCF regulates human cytomegalovirus latency via chromatin looping. Proc. Natl. Acad. Sci. USA 2024, 121, e2315860121. [Google Scholar] [CrossRef] [PubMed]
- Aslam, Y.; Williamson, J.; Romashova, V.; Elder, E.; Krishna, B.; Wills, M.; Lehner, P.; Sinclair, J.; Poole, E. Human cytomegalovirus upregulates expression of HCLS1 resulting in increased cell motility and transendothelial migration during latency. iScience 2019, 20, 60–72. [Google Scholar] [CrossRef]
- Wu, S.-e.; Miller, W.E. The HCMV US28 vGPCR induces potent Gαq/PLC-β signaling in monocytes leading to increased adhesion to endothelial cells. Virology 2016, 497, 233–243. [Google Scholar] [CrossRef]
- Elder, E.; Sinclair, J. HCMV latency: What regulates the regulators? Med. Microbiol. Immunol. 2019, 208, 431–438. [Google Scholar] [CrossRef]
- Wass, A.B.; Krishna, B.A.; Herring, L.E.; Gilbert, T.S.; Nukui, M.; Groves, I.J.; Dooley, A.L.; Kulp, K.H.; Matthews, S.M.; Rotroff, D.M. Cytomegalovirus US28 regulates cellular EphA2 to maintain viral latency. Sci. Adv. 2022, 8, eadd1168. [Google Scholar] [CrossRef]
- Krishna, B.A.; Humby, M.S.; Miller, W.E.; O’Connor, C.M. Human cytomegalovirus G protein-coupled receptor US28 promotes latency by attenuating c-fos. Proc. Natl. Acad. Sci. USA 2019, 116, 1755–1764. [Google Scholar] [CrossRef]
- Waters, S.; Agostino, M.; Lee, S.; Ariyanto, I.; Kresoje, N.; Leary, S.; Munyard, K.; Gaudieri, S.; Gaff, J.; Irish, A. Sequencing directly from clinical specimens reveals genetic variations in HCMV-encoded chemokine receptor US28 that may influence antibody levels and interactions with human chemokines. Microbiol. Spectr. 2021, 9, e00020–e00021. [Google Scholar] [CrossRef]
- Sinclair, J.; Sissons, P. Latency and reactivation of human cytomegalovirus. J. Gen. Virol. 2006, 87, 1763–1779. [Google Scholar] [CrossRef]
- Liu, X.-f.; Swaminathan, S.; Yan, S.; Engelmann, F.; Abbott, D.A.; VanOsdol, L.A.; Heald-Sargent, T.; Qiu, L.; Chen, Q.; Iovane, A. A novel murine model of differentiation-mediated cytomegalovirus reactivation from latently infected bone marrow haematopoietic cells. J. Gen. Virol. 2019, 100, 1680–1694. [Google Scholar] [CrossRef] [PubMed]
- Seckert, C.K.; Renzaho, A.; Tervo, H.-M.; Krause, C.; Deegen, P.; Kühnapfel, B.; Reddehase, M.J.; Grzimek, N.K. Liver sinusoidal endothelial cells are a site of murine cytomegalovirus latency and reactivation. J. Virol. 2009, 83, 8869–8884. [Google Scholar] [CrossRef] [PubMed]
- Pollock, J.L.; Presti, R.M.; Paetzold, S.; IVth, H.W.V. Latent murine cytomegalovirus infection in macrophages. Virology 1997, 227, 168–179. [Google Scholar] [CrossRef]
- Brautigam, A.R.; Dutko, F.J.; Olding, L.B.; Oldstone, M.B. Pathogenesis of murine cytomegalovirus infection: The macrophage as a permissive cell for cytomegalovirus infection, replication and latency. J. Gen. Virol. 1979, 44, 349–359. [Google Scholar] [CrossRef]
- Cardin, R.D.; Schaefer, G.C.; Allen, J.R.; Davis-Poynter, N.J.; Farrell, H.E. The M33 chemokine receptor homolog of murine cytomegalovirus exhibits a differential tissue-specific role during in vivo replication and latency. J. Virol. 2009, 83, 7590–7601. [Google Scholar] [CrossRef] [PubMed]
- Alcendor, D.J.; Zong, J.; Dolan, A.; Gatherer, D.; Davison, A.J.; Hayward, G.S. Patterns of divergence in the vCXCL and vGPCR gene clusters in primate cytomegalovirus genomes. Virology 2009, 395, 21–32. [Google Scholar] [CrossRef]
- Hansen, S.G.; Strelow, L.I.; Franchi, D.C.; Anders, D.G.; Wong, S.W. Complete sequence and genomic analysis of rhesus cytomegalovirus. J. Virol. 2003, 77, 6620–6636. [Google Scholar] [CrossRef]
- Marsh, A.K.; Willer, D.O.; Ambagala, A.P.; Dzamba, M.; Chan, J.K.; Pilon, R.; Fournier, J.; Sandstrom, P.; Brudno, M.; MacDonald, K.S. Genomic sequencing and characterization of cynomolgus macaque cytomegalovirus. J. Virol. 2011, 85, 12995–13009. [Google Scholar] [CrossRef]
- Penfold, M.; Schmidt, T.; Dairaghi, D.; Barry, P.; Schall, T. Characterization of the rhesus cytomegalovirus US28 locus. J. Virol. 2003, 77, 10404–10413. [Google Scholar] [CrossRef]
- Arfelt, K.N.; Fares, S.; Rosenkilde, M.M. EBV, the human host, and the 7TM receptors: Defense or offense? Prog. Mol. Biol. Transl. Sci. 2015, 129, 395–427. [Google Scholar]
- Berg, C.; Spiess, K.; Lüttichau, H.R.; Rosenkilde, M.M. Biased small-molecule ligands for selective inhibition of HIV-1 cell entry via CCR5. Pharmacol. Res. Perspect. 2016, 4, e00262. [Google Scholar] [CrossRef] [PubMed]
- Fares, S.; Spiess, K.; Olesen, E.T.; Zuo, J.; Jackson, S.; Kledal, T.N.; Wills, M.R.; Rosenkilde, M.M. Distinct roles of extracellular domains in the epstein-barr virus-encoded BILF1 receptor for signaling and major histocompatibility complex class I downregulation. mBio 2019, 10, e01707-18. [Google Scholar] [CrossRef] [PubMed]
- Krishna, B.A.; Wills, M.R.; Sinclair, J.H. Advances in the treatment of cytomegalovirus. Br. Med. Bull. 2019, 131, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Chung, Y.H.; Lee, C. US28, a virally-encoded GPCR as an antiviral target for human cytomegalovirus infection. Biomol. Ther. 2017, 25, 69. [Google Scholar] [CrossRef]
- Lüttichau, H.; Schwartz, T. Validation of chemokine receptors as drug targets. Curr. Opin. Drug Discov. Dev. 2000, 3, 610–623. [Google Scholar]
- Rosenkilde, M.M.; Kledal, T.N. Targeting herpesvirus reliance of the chemokine system. Curr. Drug Targets 2006, 7, 103–118. [Google Scholar] [CrossRef]
- Smit, J.; Vink, C.; Verzijl, D.; Casarosa, P.; Bruggeman, A.; Leurs, R. Virally encoded G protein-coupled receptors: Targets for potentially innovative anti-viral drug development. Curr. Drug Targets 2003, 4, 431–441. [Google Scholar] [CrossRef]
- Spiess, K.; Jeppesen, M.G.; Malmgaard-Clausen, M.; Krzywkowski, K.; Kledal, T.N.; Rosenkilde, M.M. Novel Chemokine-Based Immunotoxins for Potent and Selective Targeting of Cytomegalovirus Infected Cells. J. Immunol. Res. 2017, 2017, 4069260. [Google Scholar] [CrossRef]
- Hulshof, J.W.; Vischer, H.F.; Verheij, M.H.; Fratantoni, S.A.; Smit, M.J.; de Esch, I.J.; Leurs, R. Synthesis and pharmacological characterization of novel inverse agonists acting on the viral-encoded chemokine receptor US28. Bioorg. Med. Chem. 2006, 14, 7213–7230. [Google Scholar] [CrossRef]
- Hassing, H.A.; Fares, S.; Larsen, O.; Pad, H.; Hauge, M.; Jones, R.M.; Schwartz, T.W.; Hansen, H.S.; Rosenkilde, M.M. Biased signaling of lipids and allosteric actions of synthetic molecules for GPR119. Biochem. Pharmacol. 2016, 119, 66–75. [Google Scholar] [CrossRef]
- Steen, A.; Larsen, O.; Thiele, S.; Rosenkilde, M.M. Biased and g protein-independent signaling of chemokine receptors. Front. Immunol. 2014, 5, 277. [Google Scholar] [CrossRef] [PubMed]
- De Groof, T.W.; Elder, E.G.; Lim, E.Y.; Heukers, R.; Bergkamp, N.D.; Groves, I.J.; Wills, M.; Sinclair, J.H.; Smit, M.J. Targeting the latent human cytomegalovirus reservoir for T-cell-mediated killing with virus-specific nanobodies. Nat. Commun. 2021, 12, 4436. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, J.; Poole, E.; Groves, I.; Owen, D.J.; Graham, S.C.; Sinclair, J.; Kelly, B.T. Repurposing an endogenous degradation domain for antibody-mediated disposal of cell-surface proteins. EMBO Rep. 2024, 25, 951–970. [Google Scholar] [CrossRef] [PubMed]
- Elder, E.; Krishna, B.; Williamson, J.; Aslam, Y.; Farahi, N.; Wood, A.; Romashova, V.; Roche, K.; Murphy, E.; Chilvers, E. Monocytes latently infected with human cytomegalovirus evade neutrophil killing. iScience 2019, 12, 13–26. [Google Scholar] [CrossRef]
- Spiess, K.; Jeppesen, M.G.; Malmgaard-Clausen, M.; Krzywkowski, K.; Dulal, K.; Cheng, T.; Hjortø, G.M.; Larsen, O.; Burg, J.S.; Jarvis, M.A. Rationally designed chemokine-based toxin targeting the viral G protein-coupled receptor US28 potently inhibits cytomegalovirus infection in vivo. Proc. Natl. Acad. Sci. USA 2015, 112, 8427–8432. [Google Scholar] [CrossRef]
- Krishna, B.A.; Spiess, K.; Poole, E.L.; Lau, B.; Voigt, S.; Kledal, T.N.; Rosenkilde, M.M.; Sinclair, J.H. Targeting the latent cytomegalovirus reservoir with an antiviral fusion toxin protein. Nat. Commun. 2017, 8, 14321. [Google Scholar] [CrossRef]
- Ribeiro, R.V.; Ku, T.; Wang, A.; Pires, L.; Ferreira, V.H.; Michaelsen, V.; Ali, A.; Galasso, M.; Moshkelgosha, S.; Gazzalle, A. Ex vivo treatment of cytomegalovirus in human donor lungs using a novel chemokine-based immunotoxin. J. Heart Lung Transplant. 2022, 41, 287–297. [Google Scholar] [CrossRef]
- Elder, E.G. Understanding and Exploiting Viral Protein US28 During Human Cytomegalovirus Latency. Doctoral Dissertation, University of Cambridge, Cambridge, UK, 2020. [Google Scholar]
- Hesselgesser, J.; Ng, H.P.; Liang, M.; Zheng, W.; May, K.; Bauman, J.G.; Monahan, S.; Islam, I.; Wei, G.P.; Ghannam, A. Identification and characterization of small molecule functional antagonists of the CCR1 chemokine receptor. J. Biol. Chem. 1998, 273, 15687–15692. [Google Scholar] [CrossRef]
- Tschammer, N. Allosteric modulation of the G protein-coupled US28 receptor of human cytomegalovirus: Are the small-weight inverse agonist of US28 ‘camouflaged’ agonists? Bioorg. Med. Chem. Lett. 2014, 24, 3744–3747. [Google Scholar] [CrossRef]
- Kralj, A.; Wetzel, A.; Mahmoudian, S.; Stamminger, T.; Tschammer, N.; Heinrich, M.R. Identification of novel allosteric modulators for the G-protein coupled US28 receptor of human cytomegalovirus. Bioorg. Med. Chem. Lett. 2011, 21, 5446–5450. [Google Scholar] [CrossRef]
- Vischer, H.F.; Hulshof, J.W.; Hulscher, S.; Fratantoni, S.A.; Verheij, M.H.; Victorina, J.; Smit, M.J.; de Esch, I.J.; Leurs, R. Identification of novel allosteric nonpeptidergic inhibitors of the human cytomegalovirus-encoded chemokine receptor US28. Bioorg. Med. Chem. 2010, 18, 675–688. [Google Scholar] [CrossRef] [PubMed]
- Kralj, A.; Nguyen, M.-T.; Tschammer, N.; Ocampo, N.; Gesiotto, Q.; Heinrich, M.R.; Phanstiel IV, O. Development of flavonoid-based inverse agonists of the key signaling receptor US28 of human cytomegalovirus. J. Med. Chem. 2013, 56, 5019–5032. [Google Scholar] [CrossRef] [PubMed]
- Kralj, A.; Kurt, E.; Tschammer, N.; Heinrich, M.R. Synthesis and biological evaluation of biphenyl amides that modulate the US28 receptor. ChemMedChem 2014, 9, 151–168. [Google Scholar] [CrossRef] [PubMed]
- Heukers, R.; Fan, T.S.; de Wit, R.H.; van Senten, J.R.; De Groof, T.W.; Bebelman, M.P.; Lagerweij, T.; Vieira, J.; de Munnik, S.M.; Smits-de Vries, L. The constitutive activity of the virally encoded chemokine receptor US28 accelerates glioblastoma growth. Oncogene 2018, 37, 4110–4121. [Google Scholar] [CrossRef]
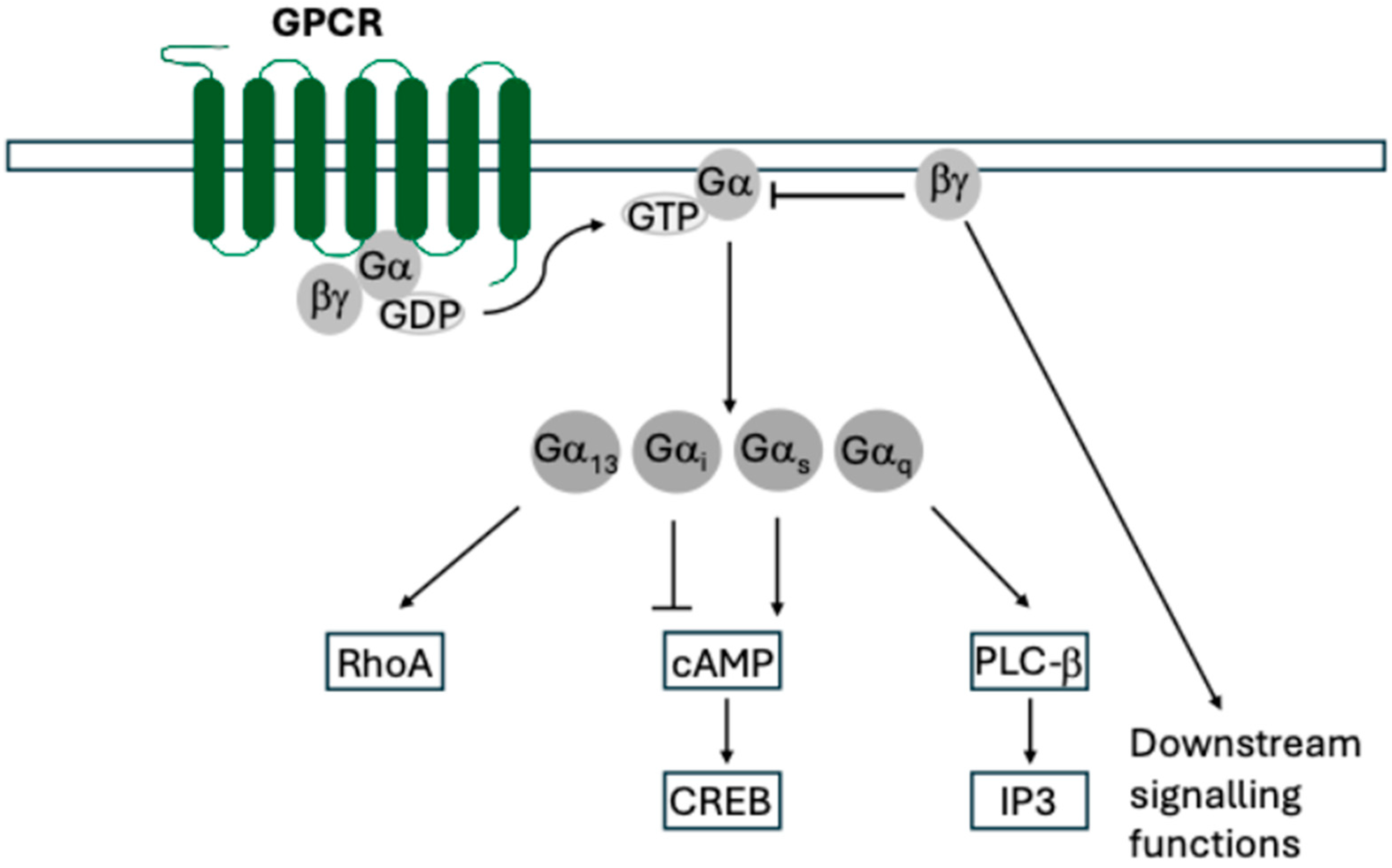

| Cell System | Ligand | Downstream Signalling and Phenotypes | References |
|---|---|---|---|
| Cos-7 cells | Constitutive but inhibited by CX3CL1 binding | PLC and NF-κB via Gαq | [87,88] |
| Cos-7 cells | Constitutive but antagonised by CCL5 binding | PLC and NF-κB via Gαo and Gαq/11 | [89] |
| Cos-7 cells | Constitutive | SRF via Gαo and Gαq11 (inhibited by Gα16) | [90] |
| Infected fibroblasts | Constitutive | PLC via Gαq | [91] |
| HEK293T | Constitutive | CREB/NFAT a via Gαq | [92] |
| NIH-3T3 and infected U373 | Constitutive | MAPK-induced VEGF-β secretion via Gαq | [93] |
| NIH-3T3 and infected fibroblasts | Constitutive | NF-κB-induced COX2 and VEGF via Gαq | [94] |
| NIH-3T3 and HEK293T, infected U373MG | Constitutive | NF-κB induction of IL6, VEGF secretion inducing JAK c/STAT3 | [95] |
| NIH-3T3 and HEK293T; infected fibroblasts and U373MG | Constitutive | β-catenin via both Gα12 and Gαq together | [96] |
| Infected HASMC, U373MG, HFFs, and HUVECs | Constitutive | PLC-β via Gαq and Gα11 in all cell types tested | [85] |
| U251 and NIH-3T3 | Constitutive | VEGF secretion and HIF1-α activation with Akt and PKM2 | [97] |
| K562 | CCL2 and CCL5 | Calcium release | [98] |
| HEK293T and infected HUVECs | CCL7 and CCL5 | Calcium release and MAPK via Gα16 | [99] |
| Infected fibroblasts | CCL2 and CCL5 | Calcium release | [100] |
| Infected arterial SMCs | CCL5 (inhibited by CX3CL1) | Chemotaxis via Gα12/13 | [101,102,103,104] |
| Mouse macrophages | CX3CL1 (inhibited by CCL5) | Chemotaxis via Gαq | [103] |
| Non-proliferating hippocampal cells and HUVECs | CCL5 | “Invasive phenotypes” via STAT3, AKT, ERK1/2, FAK, Src, and eNOS | [105] |
| Infected HASMC and HFFs | CCL5 | Calcium release via Gα12/13 | [85] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fares, S.; Krishna, B.A. Why Are Cytomegalovirus-Encoded G-Protein-Coupled Receptors Essential for Infection but Only Variably Conserved? Pathogens 2025, 14, 245. https://doi.org/10.3390/pathogens14030245
Fares S, Krishna BA. Why Are Cytomegalovirus-Encoded G-Protein-Coupled Receptors Essential for Infection but Only Variably Conserved? Pathogens. 2025; 14(3):245. https://doi.org/10.3390/pathogens14030245
Chicago/Turabian StyleFares, Suzan, and Benjamin A. Krishna. 2025. "Why Are Cytomegalovirus-Encoded G-Protein-Coupled Receptors Essential for Infection but Only Variably Conserved?" Pathogens 14, no. 3: 245. https://doi.org/10.3390/pathogens14030245
APA StyleFares, S., & Krishna, B. A. (2025). Why Are Cytomegalovirus-Encoded G-Protein-Coupled Receptors Essential for Infection but Only Variably Conserved? Pathogens, 14(3), 245. https://doi.org/10.3390/pathogens14030245