

Utility of an In-Vitro Micro-Neutralizing Test in Comparison to a Plaque Reduction Neutralization Test for Dengue Virus, Japanese Encephalitis Virus, and Zika Virus Serology and Drug Screening

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Monoclonal Antibodies, Serum Samples, and Therapeutic Drug Candidates
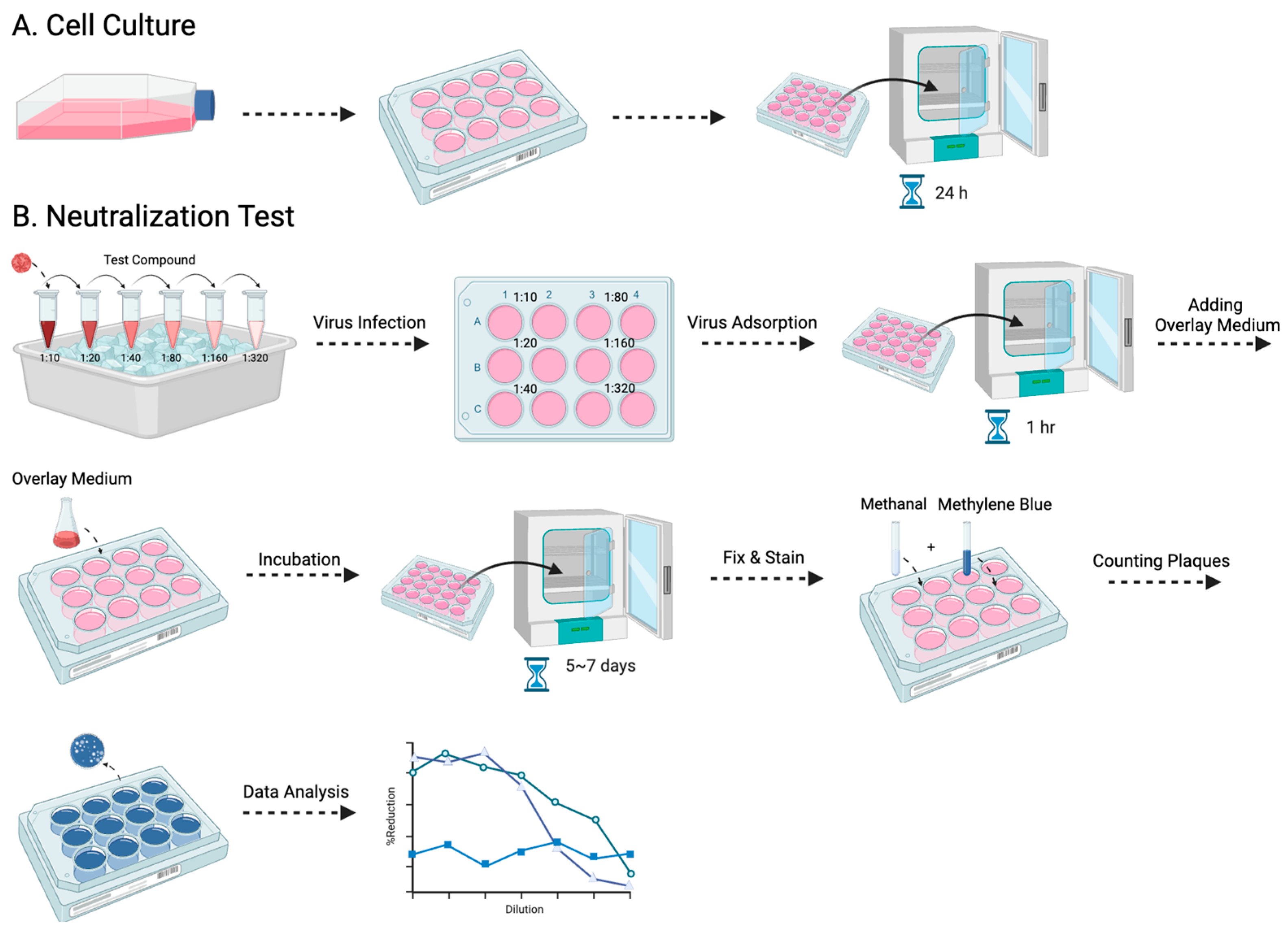
2.3. Plaque Reduction Neutralization Test
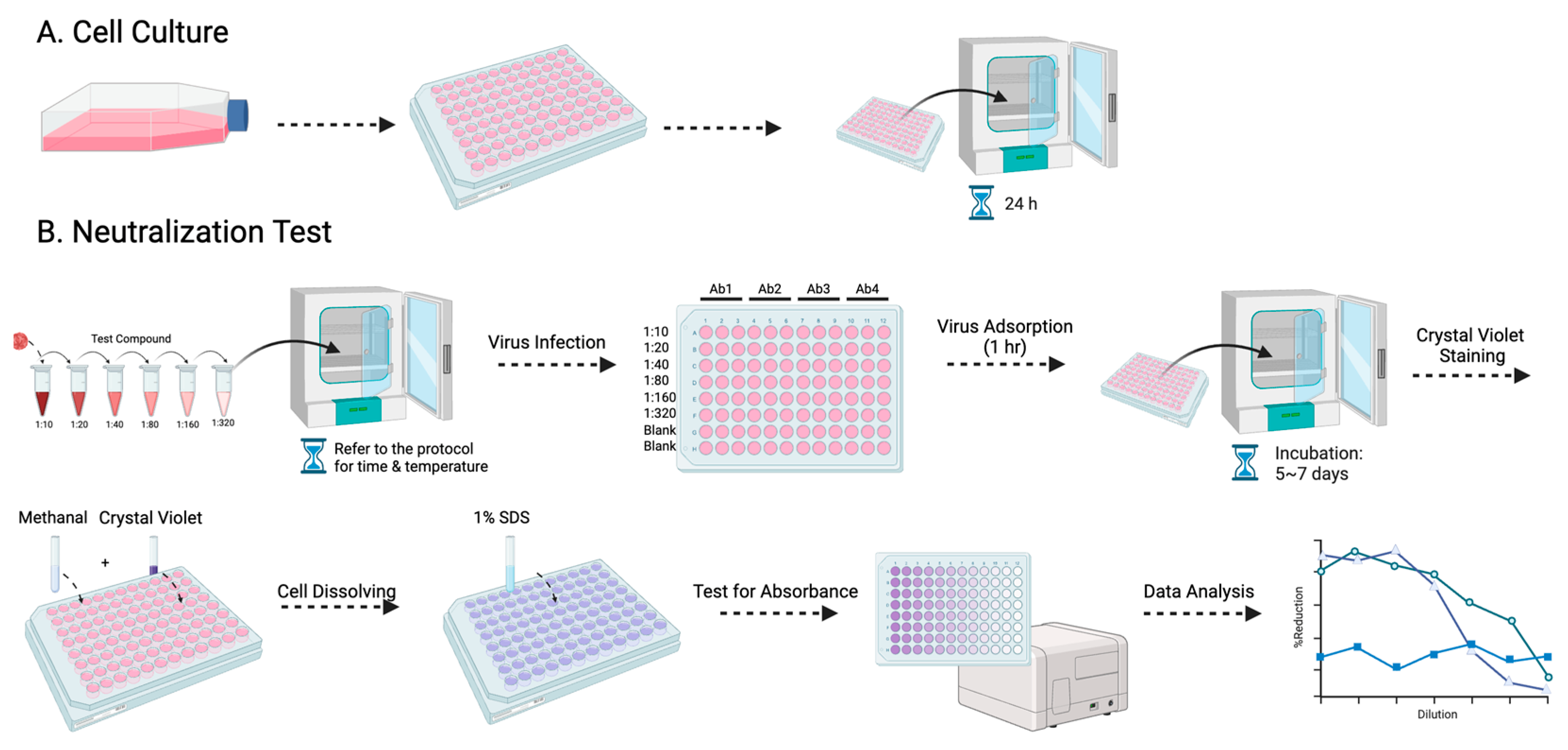
2.4. Micro-Neutralization Test
2.5. Data Analysis
3. Results
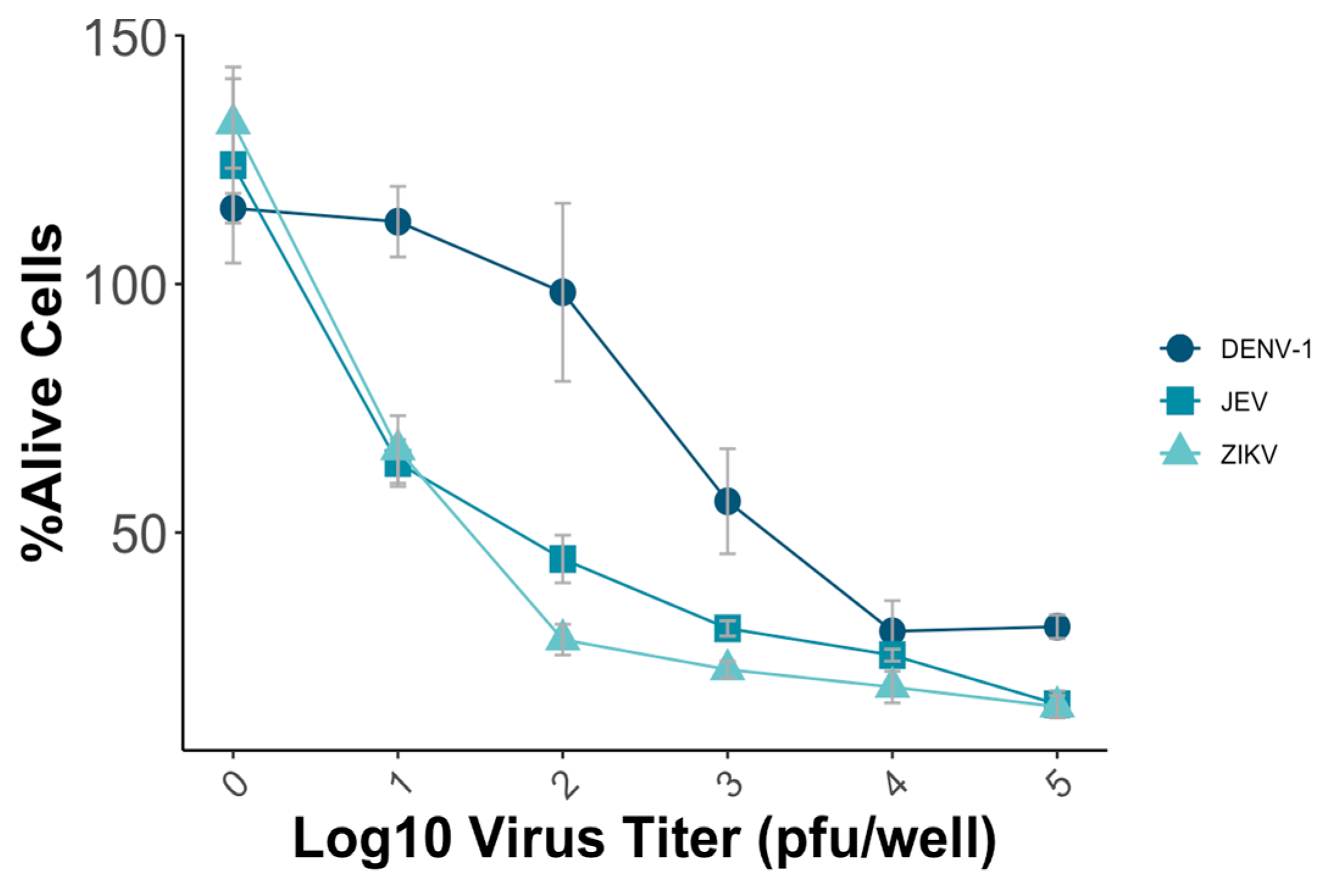
3.1. Evaluation of the Microneutralization Test
3.2. Determination of the MNT Titers Using Monoclonal Antibodies
3.3. Determination of the MNT Titers Using Serum Samples
3.4. Determination of the MNT Titers Using Antiviral Drug Candidates
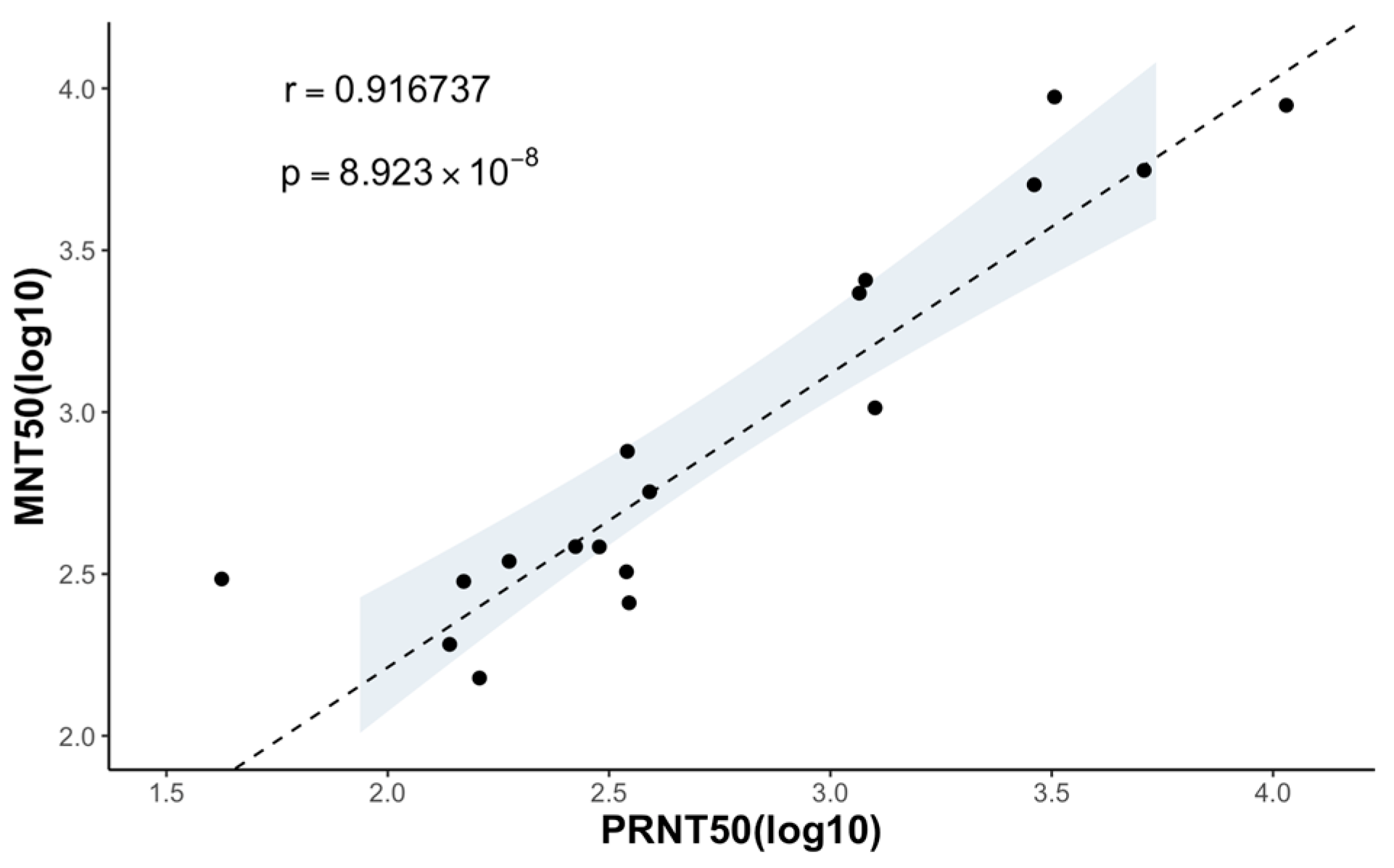
3.5. Comparison of the MNT to the PRNT
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pierson, T.C.; Diamond, M.S. The Continued Threat of Emerging Flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The Global Distribution and Burden of Dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Daep, C.A.; Muñoz-Jordán, J.L.; Eugenin, E.A. Flaviviruses, an Expanding Threat in Public Health: Focus on Dengue, West Nile, and Japanese Encephalitis Virus. J. Neurovirol. 2014, 20, 539–560. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C.; Diamond, M.S. The Emergence of Zika Virus and Its New Clinical Syndromes. Nature 2018, 560, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-J.S.; Higgs, S.; Horne, K.M.; Vanlandingham, D.L. Flavivirus-Mosquito Interactions. Viruses 2014, 6, 4703–4730. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Quispe, C.; Herrera-Bravo, J.; Sarkar, C.; Sharma, R.; Garg, N.; Fredes, L.I.; Martorell, M.; Alshehri, M.M.; Sharifi-Rad, J.; et al. Production, Transmission, Pathogenesis, and Control of Dengue Virus: A Literature-Based Undivided Perspective. BioMed Res. Int. 2021, 2021, 4224816. [Google Scholar] [CrossRef]
- Mulvey, P.; Duong, V.; Boyer, S.; Burgess, G.; Williams, D.T.; Dussart, P.; Horwood, P.F. The Ecology and Evolution of Japanese Encephalitis Virus. Pathogens 2021, 10, 1534. [Google Scholar] [CrossRef]
- Vasilakis, N.; Weaver, S.C. Flavivirus Transmission Focusing on Zika. Curr. Opin. Virol. 2017, 22, 30–35. [Google Scholar] [CrossRef]
- Mackenzie, J.S.; Gubler, D.J.; Petersen, L.R. Emerging Flaviviruses: The Spread and Resurgence of Japanese Encephalitis, West Nile and Dengue Viruses. Nat. Med. 2004, 10, S98–S109. [Google Scholar] [CrossRef]
- Sikka, V.; Chattu, V.K.; Popli, R.K.; Galwankar, S.C.; Kelkar, D.; Sawicki, S.G.; Stawicki, S.P.; Papadimos, T.J. The Emergence of Zika Virus as a Global Health Security Threat: A Review and a Consensus Statement of the INDUSEM Joint Working Group (JWG). J. Glob. Infect. Dis. 2016, 8, 3–15. [Google Scholar] [CrossRef]
- Michael, B.N.; Axel, K.; John, E.; Chusak, P.; Jose, L.S.M.; Renu, D.D.; Raman, V.; Olaf, H. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control: New Edition; WHO Guidelines Approved by the Guidelines Review Committee; World Health Organization: Geneva, Switzerland, 2009; ISBN 978-92-4-154787-1. [Google Scholar]
- Kuwayama, M.; Ito, M.; Takao, S.; Shimazu, Y.; Fukuda, S.; Miyazaki, K.; Kurane, I.; Takasaki, T. Japanese Encephalitis Virus in Meningitis Patients, Japan. Emerg. Infect. Dis. 2005, 11, 471–473. [Google Scholar] [CrossRef] [PubMed]
- Marano, G.; Pupella, S.; Vaglio, S.; Liumbruno, G.M.; Grazzini, G. Zika Virus and the Never-Ending Story of Emerging Pathogens and Transfusion Medicine. Blood Transfus. 2016, 14, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Boldescu, V.; Behnam, M.A.M.; Vasilakis, N.; Klein, C.D. Broad-Spectrum Agents for Flaviviral Infections: Dengue, Zika and Beyond. Nat. Rev. Drug Discov. 2017, 16, 565–586. [Google Scholar] [CrossRef] [PubMed]
- Russell, P.K.; Nisalak, A.; Sukhavachana, P.; Vivona, S. A Plaque Reduction Test for Dengue Virus Neutralizing Antibodies. J. Immunol. Baltim. Md. 1950 1967, 99, 285–290. [Google Scholar] [CrossRef]
- Kuno, G. Serodiagnosis of Flaviviral Infections and Vaccinations in Humans. Adv. Virus Res. 2003, 61, 3–65. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Xie, X.; Ren, P.; Loeffelholz, M.J.; Yang, Y.; Furuya, A.; Dupuis, A.P.; Kramer, L.D.; Wong, S.J.; Shi, P.-Y. A Rapid Zika Diagnostic Assay to Measure Neutralizing Antibodies in Patients. EBioMedicine 2017, 17, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Moi, M.L.; Lim, C.-K.; Kotaki, A.; Takasaki, T.; Kurane, I. Development of an Antibody-Dependent Enhancement Assay for Dengue Virus Using Stable BHK-21 Cell Lines Expressing Fc GammaRIIA. J. Virol. Methods 2010, 163, 205–209. [Google Scholar] [CrossRef]
- Lim, S.P. Dengue Drug Discovery: Progress, Challenges and Outlook. Antivir. Res. 2019, 163, 156–178. [Google Scholar] [CrossRef]
- Bernatchez, J.A.; Tran, L.T.; Li, J.; Luan, Y.; Siqueira-Neto, J.L.; Li, R. Drugs for the Treatment of Zika Virus Infection. J. Med. Chem. 2020, 63, 470–489. [Google Scholar] [CrossRef]
- Felicetti, T.; Manfroni, G.; Cecchetti, V.; Cannalire, R. Broad-Spectrum Flavivirus Inhibitors: A Medicinal Chemistry Point of View. ChemMedChem 2020, 15, 2391–2419. [Google Scholar] [CrossRef]
- Samrat, S.K.; Xu, J.; Li, Z.; Zhou, J.; Li, H. Antiviral Agents against Flavivirus Protease: Prospect and Future Direction. Pathogens 2022, 11, 293. [Google Scholar] [CrossRef] [PubMed]
- Crance, J.M.; Scaramozzino, N.; Jouan, A.; Garin, D. Interferon, Ribavirin, 6-Azauridine and Glycyrrhizin: Antiviral Compounds Active against Pathogenic Flaviviruses. Antivir. Res. 2003, 58, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C.; Fremont, D.H.; Kuhn, R.J.; Diamond, M.S. Structural Insights into the Mechanisms of Antibody-Mediated Neutralization of Flavivirus Infection: Implications for Vaccine Development. Cell Host Microbe 2008, 4, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Katzelnick, L.C.; Montoya, M.; Gresh, L.; Balmaseda, A.; Harris, E. Neutralizing Antibody Titers against Dengue Virus Correlate with Protection from Symptomatic Infection in a Longitudinal Cohort. Proc. Natl. Acad. Sci. USA 2016, 113, 728–733. [Google Scholar] [CrossRef] [PubMed]
- Van Gessel, Y.; Klade, C.S.; Putnak, R.; Formica, A.; Krasaesub, S.; Spruth, M.; Cena, B.; Tungtaeng, A.; Gettayacamin, M.; Dewasthaly, S. Correlation of Protection against Japanese Encephalitis Virus and JE Vaccine (IXIARO®) Induced Neutralizing Antibody Titers. Vaccine 2011, 29, 5925–5931. [Google Scholar] [CrossRef] [PubMed]
- Maciejewski, S.; Ruckwardt, T.J.; Morabito, K.M.; Foreman, B.M.; Burgomaster, K.E.; Gordon, D.N.; Pelc, R.S.; DeMaso, C.R.; Ko, S.-Y.; Fisher, B.E.; et al. Distinct Neutralizing Antibody Correlates of Protection among Related Zika Virus Vaccines Identify a Role for Antibody Quality. Sci. Transl. Med. 2020, 12, eaaw9066. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Hansen, M.B.; Nielsen, S.E.; Berg, K. Re-Examination and Further Development of a Precise and Rapid Dye Method for Measuring Cell Growth/Cell Kill. J. Immunol. Methods 1989, 119, 203–210. [Google Scholar] [CrossRef]
- Gu, L.; Schneller, S.W.; Li, Q. Assays for the Identification of Novel Antivirals against Bluetongue Virus. J. Vis. Exp. JoVE 2013, 80, 50820. [Google Scholar] [CrossRef]
- Thomas, S.J.; Nisalak, A.; Anderson, K.B.; Libraty, D.H.; Kalayanarooj, S.; Vaughn, D.W.; Putnak, R.; Gibbons, R.V.; Jarman, R.; Endy, T.P. Dengue Plaque Reduction Neutralization Test (PRNT) in Primary and Secondary Dengue Virus Infections: How Alterations in Assay Conditions Impact Performance. Am. J. Trop. Med. Hyg. 2009, 81, 825–833. [Google Scholar] [CrossRef]
- Bewley, K.R.; Coombes, N.S.; Gagnon, L.; McInroy, L.; Baker, N.; Shaik, I.; St-Jean, J.R.; St-Amant, N.; Buttigieg, K.R.; Humphries, H.E.; et al. Quantification of SARS-CoV-2 Neutralizing Antibody by Wild-Type Plaque Reduction Neutralization, Microneutralization and Pseudotyped Virus Neutralization Assays. Nat. Protoc. 2021, 16, 3114–3140. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ma, C.; Wang, J. Cytopathic Effect Assay and Plaque Assay to Evaluate in Vitro Activity of Antiviral Compounds Against Human Coronaviruses 229E, OC43, and NL63. Bio Protoc. 2022, 12, e4314. [Google Scholar] [CrossRef] [PubMed]
- Amanat, F.; White, K.M.; Miorin, L.; Strohmeier, S.; McMahon, M.; Meade, P.; Liu, W.; Albrecht, R.A.; Simon, V.; Martinez-Sobrido, L.; et al. An In Vitro Microneutralization Assay for SARS-CoV-2 Serology and Drug Screening. Curr. Protoc. Microbiol. 2020, 58, e108. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | mAb 12D11 | mAb 4G2 | mAb B247 | mAb B376 | ||||
|---|---|---|---|---|---|---|---|---|
| MNT50 1 | PRNT50 2 | MNT50 | PRNT50 | MNT50 | PRNT50 | MNT50 | PRNT50 | |
| DENV-1 | 640 | 1280 | 160 | 160 | <10 3 | <10 | 1280 | 2560 |
| JEV | <10 | <10 | <10 | <10 | <10 | <10 | <10 | <10 |
| ZIKV | <10 | <10 | <10 | <10 | <10 | <10 | <10 | <10 |
| Sample Code | DENV-1 | |
|---|---|---|
| MNT50 1 | PRNT50 2 | |
| 1 | 10,240 | 2560 |
| 2 | 5260 | 2560 |
| 3 | 10,240 | 10,240 |
| 4 | 2560 | 1280 |
| 5 | 320 | 160 |
| 6 | 1280 | 1280 |
| 7 | 5260 | 5260 |
| 8 | 320 | 320 |
| 9 | 320 | 40 |
| 10 | 320 | 320 |
| 11 | 640 | 320 |
| 12 | 640 | 320 |
| 13 | 320 | 320 |
| 14 | 320 | 320 |
| 15 | 320 | 160 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haga, K.; Chen, Z.; Himeno, M.; Majima, R.; Moi, M.L. Utility of an In-Vitro Micro-Neutralizing Test in Comparison to a Plaque Reduction Neutralization Test for Dengue Virus, Japanese Encephalitis Virus, and Zika Virus Serology and Drug Screening. Pathogens 2024, 13, 8. https://doi.org/10.3390/pathogens13010008
Haga K, Chen Z, Himeno M, Majima R, Moi ML. Utility of an In-Vitro Micro-Neutralizing Test in Comparison to a Plaque Reduction Neutralization Test for Dengue Virus, Japanese Encephalitis Virus, and Zika Virus Serology and Drug Screening. Pathogens. 2024; 13(1):8. https://doi.org/10.3390/pathogens13010008
Chicago/Turabian StyleHaga, Kazumi, Zhenying (Nancy) Chen, Misao Himeno, Ryuichi Majima, and Meng Ling Moi. 2024. "Utility of an In-Vitro Micro-Neutralizing Test in Comparison to a Plaque Reduction Neutralization Test for Dengue Virus, Japanese Encephalitis Virus, and Zika Virus Serology and Drug Screening" Pathogens 13, no. 1: 8. https://doi.org/10.3390/pathogens13010008
APA StyleHaga, K., Chen, Z., Himeno, M., Majima, R., & Moi, M. L. (2024). Utility of an In-Vitro Micro-Neutralizing Test in Comparison to a Plaque Reduction Neutralization Test for Dengue Virus, Japanese Encephalitis Virus, and Zika Virus Serology and Drug Screening. Pathogens, 13(1), 8. https://doi.org/10.3390/pathogens13010008