



The Characterisation of Lactococcus garvieae Isolated in an Outbreak of Septicaemic Disease in Farmed Sea Bass (Dicentrarchus labrax, Linnaues 1758) in Italy

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Case Presentation
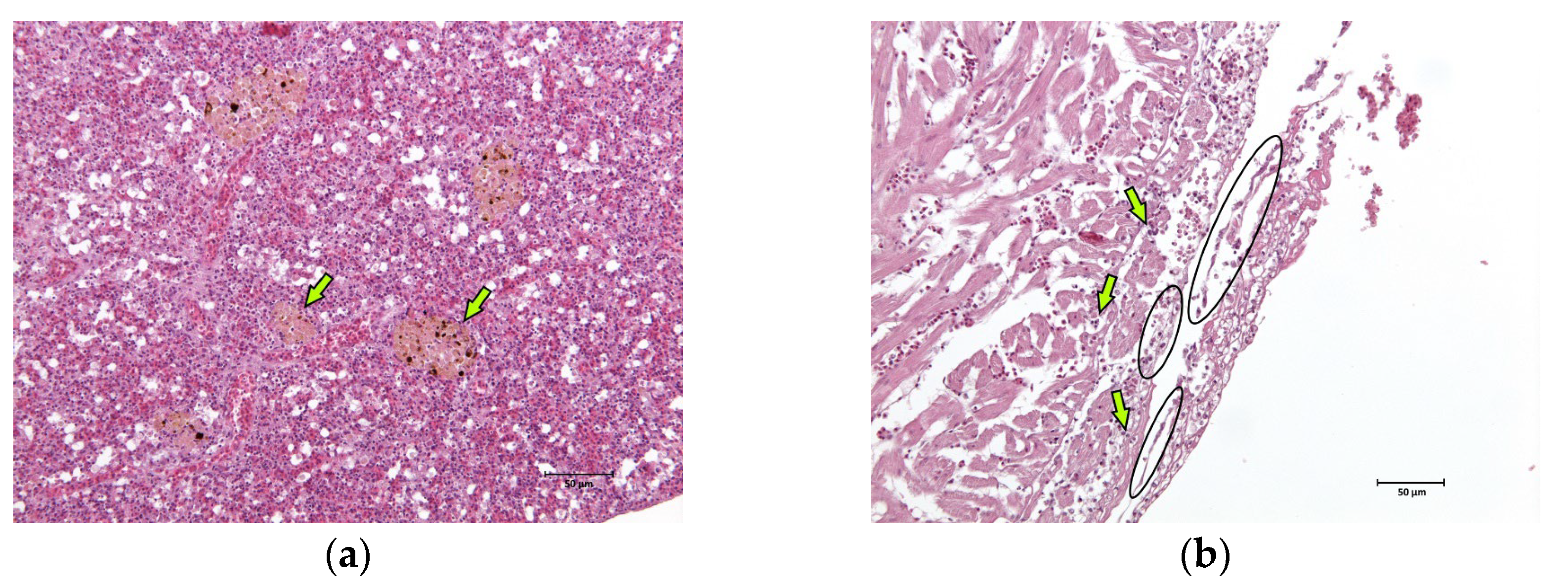
2.1. Anatomopathological Examinations
2.2. Bacteriological and Biochemical Tests
2.3. Molecular Analysis
2.4. Antimicrobial Therapy and Susceptibility Tests
3. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meyburgh, C.M.; Bragg, R.R.; Boucher, C.E. Lactococcus garvieae: An emerging bacterial pathogen of fish. Dis. Aquat. Organ. 2017, 123, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Soltani, M.; Nikbakht, G.; Mousavi, H.A.E.; Ahmadzadeh, N. Epizootic outbreaks of lactococcosis caused by in farmed rainbow trout (Oncorhynchus mykiss) in Iran. Bull. Eur. Assoc. Fish. Pathol. 2008, 28, 207–212. [Google Scholar]
- Ghittino, C.; Latini, M.; Agnetti, F.; Panzieri, C.; Lauro, L.; Ciappelloni, R.; Petracca, G. Emerging pathologies in aquaculture: Effects on production and food safety. Vet. Res. Commun. 2003, 27, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Soltani, M.; Baldisserotto, B.; Shekarabi, S.P.H.; Shafiei, S.; Bashiri, M. Lactococcosis a Re-Emerging Disease in Aquaculture: Disease Significant and Phytotherapy. Vet. Sci. 2021, 8, 181. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.J.; Pasnik, D.J.; Klesius, P.H.; Al-Ablani, S. First report of Streptococcus agalactiae and Lactococcus garvieae from a wild bottlenose dolphin (Tursiops truncatus). J. Wildl. Dis. 2006, 42, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Francés-Cuesta, C.; Ansari, I.; Fernández-Garáyzabal, J.F.; Gibello, A.; González-Candelas, F. Comparative genomics and evolutionary analysis of Lactococcus garvieae isolated from human endocarditis. Microb. Genom. 2022, 8, 000771. [Google Scholar] [CrossRef] [PubMed]
- Akmal, M.; Yoshida, T.; Nishiki, I. Complete Genome Sequence of Lactococcus garvieae MS210922A, Isolated from Farmed Greater Amberjack (Seriola dumerili) in Japan. Microbiol. Resour. Announc. 2022, 11, 1–2. [Google Scholar] [CrossRef]
- Kotzamanidis, C.; Malousi, A.; Bitchava, K.; Vafeas, G.; Chatzidimitriou, D.; Skoura, L.; Papadimitriou, E.; Chatzopoulou, F.; Zdragas, A. First Report of Isolation and Genome Sequence of L. petauri Strain from a Rainbow Trout Lactococcosis Outbreak. Curr. Microbiol. 2020, 77, 1089–1096. [Google Scholar] [CrossRef]
- Li, T.T.; Tian, W.L.; Gu, C.T. Elevation of Lactococcus lactis subsp. cremoris to the species level as Lactococcus cremoris sp. nov. and transfer of Lactococcus lactis subsp. tructae to Lactococcus cremoris as Lactococcus cremoris subsp. tructae comb. nov. Int. J. Syst. Evol. Microbiol. 2021, 71, 004727. [Google Scholar] [CrossRef]
- Pérez, T.; Balcázar, J.L.; Peix, A.; Valverde, A.; Velázquez, E.; de Blas, I.; Ruiz-Zarzuela, I. Lactococcus lactis subsp. tructae subsp. nov. isolated from the intestinal mucus of brown trout (Salmo trutta) and rainbow trout (Oncorhynchus mykiss). Int. J. Syst. Evol. Microbiol. 2011, 61, 1894–1898. [Google Scholar] [CrossRef]
- Akali, T.; Çiğdem, Ü.; Göken, Z. Pathological aspects of experimental infection of Lactococcus garvieae in European Sea Bass (Dicentrarchus labrax L.): Clinical, hematological, and histopathological parameters. Aquat. Res. 2022, 5, 219–229. [Google Scholar] [CrossRef]
- Tanekhy, M. Some study on bacterial infection in some cultured marine fish. J. Arab. Aquac. Soc. 2013, 8, 163–178. [Google Scholar]
- Arechavala-Lopez, P.; Sanchez-Jerez, P.; Bayle-Sempere, J.T.; Sfakianakis, D.G.; Somarakis, S. Morphological differences between wild and farmed Mediterranean fish. Hydrobiologia 2012, 679, 217–231. [Google Scholar] [CrossRef]
- FAO. FAO Fisheries and Aquaculture—FishStatJ—Software for Fishery and Aquaculture Statistical Time Series. Available online: https://www.fao.org/fishery/en/topic/166235?lang=en (accessed on 21 November 2023).
- Lucchetti, A. La Spigola o Branzino. Il Pesce 2008, 4, 1–7. [Google Scholar]
- Vandeputte, M.; Gagnaire, P.A.; Allal, F. The European sea bass: A key marine fish model in the wild and in aquaculture. Anim. Genet. 2019, 50, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Di Trapani, A.M.; Sgroi, F.; Testa, R.; Tudisca, S. Economic comparison between offshore and inshore aquaculture production systems of European sea bass in Italy. Aquaculture 2014, 434, 334–339. [Google Scholar] [CrossRef]
- Parisi, G.; Terova, G.; Gasco, L.; Piccolo, G.; Roncarati, A.; Moretti, V.M.; Centoducati, G.; Gatta, P.P.; Pais, A. Current status and future perspectives of Italian finfish aquaculture. Rev. Fish. Biol. Fish. 2014, 24, 15–73. [Google Scholar] [CrossRef]
- Zoli, M.; Rossi, L.; Costantini, M.; Bibbiani, C.; Fronte, B.; Brambilla, F.; Bacenetti, J. Quantification and characterization of the environmental impact of sea bream and sea bass production in Italy. Clean. Environ. Syst. 2023, 9, 100118. [Google Scholar] [CrossRef]
- Forchino, A.; Borja, A.; Brambilla, F.; Rodríguez, J.G.; Muxika, I.; Terova, G.; Saroglia, M. Evaluating the influence of off-shore cage aquaculture on the benthic ecosystem in Alghero Bay (Sardinia, Italy) using AMBI and M-AMBI. Ecol. Indic. 2011, 11, 1112–1122. [Google Scholar] [CrossRef]
- Grigorakis, K.; Rigos, G. Aquaculture effects on environmental and public welfare—The case of Mediterranean mariculture. Chemosphere 2011, 85, 899–919. [Google Scholar] [CrossRef]
- Sicuro, B.; Tarantola, M.; Valle, E. Italian aquaculture and the diffusion of alien species: Costs and benefits. Aquac. Res. 2016, 47, 3718–3728. [Google Scholar] [CrossRef]
- Mirto, S.; Pusceddu, A.; Gambi, C.; Holmer, M.; Danovaro, R. Ecological assessment of aquaculture impact in the Mediterranean Sea. In Proceedings of the Impact of Mariculture on Coastal Ecosystems, CIESM Workshop Monographs, Lisboa, Portugal, 21–24 February 2007; pp. 21–28. [Google Scholar]
- Nishizawa, T.; Mori, K.; Nakai, T.; Furusawa, I.; Muroga, K. Polymerase Chain-Reaction (PCR) Amplification of Rna of Striped Jack Nervous Necrosis Virus (SJNNV). Dis. Aquat. Organ. 1994, 18, 103–107. [Google Scholar] [CrossRef]
- Onuk, E.E.; Çiftçi, A. Phenotypic, genotypic characterisation and antimicrobial susceptibility determination of Lactococcus garvieae strains; Kafkas Üniversitesi: Kars, Turkey, 2013. [Google Scholar]
- Patel, J.B.; Leonard, D.G.B.; Pan, X.; Musser, J.M.; Berman, R.E.; Nachamkin, I. Sequence-based identification of Mycobacterium species using the MicroSeq 500 16S rDNA bacterial identification system. J. Clin. Microbiol. 2000, 38, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes de novo assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Olson, R.D.; Assaf, R.; Brettin, T.; Conrad, N.; Cucinell, C.; Davis, J.J.; Dempsey, D.M.; Dickerman, A.; Dietrich, E.M.; Kenyon, R.W.; et al. Introducing the Bacterial and Viral Bioinformatics Resource Center (BV-BRC): A resource combining PATRIC, IRD and ViPR. Nucleic Acids Res. 2023, 51, D678–D689. [Google Scholar] [CrossRef]
- Altinok, I.; Ozturk, R.C.; Ture, M. NGS analysis revealed that Lactococcus garvieae Lg-Per was Lactococcus petauri in Turkey. J. Fish. Dis. 2022, 45, 1839–1843. [Google Scholar] [CrossRef]
- Egger, R.C.; Rosa, J.C.C.; Resende, L.F.L.; de Padua, S.B.; Barbosa, F.D.; Zerbini, M.T.; Tavares, G.C.; Figueiredo, H.C.P. Emerging fish pathogens Lactococcus petauri and L. garvieae in Nile tilapia (Oreochromis niloticus). Aquaculture 2023, 565, 739093. [Google Scholar] [CrossRef]
- Ou, Y.J.; Ren, Q.Q.; Fang, S.T.; Wu, J.G.; Jiang, Y.X.; Chen, Y.R.; Zhong, Y.; Wang, D.D.; Zhang, G.X. Complete genome insights into Lactococcus petauri CF11 isolated from a healthy human gut using second- and third-generation sequencing. Front. Genet. 2020, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Stoppani, N.; Colussi, S.; Pastorino, P.; Prearo, M.; Sciuto, S.; Altinok, I.; Öztürk, R.Ç.; Ture, M.; Vela, A.I.; Blanco, M.D.; et al. 16S-23S rRNA Internal Transcribed Spacer Region (ITS) Sequencing: A Potential Molecular Diagnostic Tool for Differentiating. Microorganisms 2023, 11, 1320. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial-DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Ohbayashi, K.; Oinaka, D.; Hoai, T.D.; Yoshida, T.; Nishiki, I. PCR-mediated Identification of the Newly Emerging Pathogen Lactococcus garvieae Serotype II from Seriola quinqueradiata and S. dumerili. Fish. Pathol. 2017, 52, 46–49. [Google Scholar] [CrossRef][Green Version]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Morita, H.; Toh, H.; Oshima, K.; Yoshizaki, M.; Kawanishi, M.; Nakaya, K.; Suzuki, T.; Miyauchi, E.; Ishii, Y.; Tanabe, S.; et al. Complete Genome Sequence and Comparative Analysis of the Fish Pathogen. PLoS ONE 2011, 6, e104984. [Google Scholar] [CrossRef]
- Eraclio, G.; Ricci, G.; Quattrini, M.; Moroni, P.; Fortina, M.G. Detection of virulence-related genes in Lactococcus garvieae and their expression in response to different conditions. Folia Microbiol. 2018, 63, 291–298. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Proc. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- CLSI. M45—Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- CLSI. VET01—Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 5th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Eldar, A.; Ghittino, C.; Asanta, L.; Bozzetta, E.; Goria, M.; Prearo, M.; Bercovier, H. Enterococcus seriolicida is a junior synonym of Lactococcus garvieae, a causative agent of septicemia and meningoencephalitis in fish. Curr. Microbiol. 1996, 32, 85–88. [Google Scholar] [PubMed]
- Prieta, J.; Domenech, A.M.; Fernandez-Garaizabal, J.F. Lactococcosis de la trucha arco iris. Med. Vet. 1993, 10, 367–373. [Google Scholar]
- Algöet, M.; Bayley, A.E.; Roberts, E.G.; Feist, S.W.; Wheeler, R.W.; Verner-Jeffreys, D.W. Susceptibility of selected freshwater fish species to a UK Lactococcus garvieae isolate. J. Fish. Dis. 2009, 32, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Raissy, M.; Moumeni, M. Detection of antibiotic resistance genes in some Lactococcus garvieae strains isolated from infected rainbow trout. Iran. J. Fish. Sci. 2016, 15, 221–229. [Google Scholar]
- Sezgin, S.S.; Yilmaz, M.; Arslan, T.; Kubilay, A. Current antibiotic sensitivity of Lactococcus garvieae in rainbow trout (Oncorhynchus mykiss) farms from Southwestern Turkey. J. Agric. Sci. Tarim. Bilim. Derg. 2023, 29, 638–642. [Google Scholar] [CrossRef]
- Ceschia, G.; Giorgetti, G.; Giavenni, R.; Sarti, M. A new problem for Italian trout farms: Streptococcosis in rainbow trout (Oncorhynchus mykiss). Bull. Eur. Assoc. Fish. Pathol. 1992, 12, 71. [Google Scholar]
- Eldar, A.; Ghittino, C. Lactococcus garvieae and Streptococcus iniae infection in rainbow trout (Oncorhynchus mykiss): Similar but different diseases. Dis. Aquat. Organ. 1999, 36, 227–231. [Google Scholar] [CrossRef]
- Ghittino, P.; Prearo, M. Report of Streptococcosis in rainbow trout (Oncorhynchus mykiss) in Italy: Preliminary note. Boll. Soc. Ital. Patol. Ittica 1992, 8, 4–9. [Google Scholar]
- Pastorino, P.; Alonso, A.I.V.; Colussi, S.; Cavazza, G.; Menconi, V.; Mugetti, D.; Righetti, M.; Barbero, R.; Zuccaro, G.; Fernández-Garayzábal, J.F.; et al. A Summer Mortality Outbreak of Lactococcosis by Lactococcus garvieae in a Raceway System Affecting Farmed Rainbow Trout (Oncorhynchus mykiss) and Brook Trout (Salvelinus fontinalis). Animals 2019, 9, 1043. [Google Scholar] [CrossRef]
- Hine, M.; Adams, S.; Arthur, J.; Bartley, D.; Bondad-Reantaso, M.; Chávez, C.; Clausen, J.; Dalsgaard, A.; Flegel, T.; Gudding, R. Improving biosecurity: A necessity for aquaculture sustainability. In Proceedings of the Global Conference on Aquaculture 2010, Phuket, Thailand, 22–25 September 2010; pp. 437–494. [Google Scholar]
- Palić, D.; Scarfe, A.D.; Walster, C.I. A standardized approach for meeting national and international aquaculture biosecurity requirements for preventing, controlling, and eradicating infectious diseases. J. Appl. Aquacult. 2015, 27, 185–219. [Google Scholar] [CrossRef]
- Barnes, A.C.; Guyot, C.; Hansen, B.G.; Mackenzie, K.; Horne, M.T.; Ellis, A.E. Resistance to serum killing may contribute to differences in the abilities of capsulate and non-capsulated isolates of Lactococcus garvieae to cause disease in rainbow trout (Oncorhynchus mykiss L.). Fish. Shellfish. Immunol. 2002, 12, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, M.; Yoshida, T.; Yagashiro, S.; Kijima, M.; Yagyu, K.; Nakai, T.; Murakami, M.; Morita, H.; Suzuki, S. Differences between Lactococcus garvieae isolated from the genus Seriola in Japan and those isolated from other animals (trout, terrestrial animals from Europe) with regard to pathogenicity, phage susceptibility and genetic characterization. J. Appl. Microbiol. 2006, 101, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Amendola, S.; Maimone, F.; Pelino, V.; Pasini, A. New records of monthly temperature extremes as a signal of climate change in Italy. Int. J. Climatol. 2019, 39, 2491–2503. [Google Scholar] [CrossRef]
- Anderson, J.; Arblaster, K.; Bartley, J.; Cooper, T.; Kettunen, M.; Kaphengst, T.; Leipprand, A.; Laaser, C.; Umpfenbach, K.; Kuusisto, E. Climate change-induced water stress and its impact on natural and managed ecosystems. In Climate Change-Induced Water Stress and Its Impact on Natural and Managed Ecosystems; European Parliament: Luxembourg, 2006. [Google Scholar]
- Beaugrand, G.; Reid, P.C. Long-term changes in phytoplankton, zooplankton and salmon related to climate. Glob. Chang. Biol. 2003, 9, 801–817. [Google Scholar] [CrossRef]
- Tzanatos, E.; Raitsos, D.E.; Triantafyllou, G.; Somarakis, S.; Tsonis, A.A. Indications of a climate effect on Mediterranean fisheries. Clim. Change 2014, 122, 41–54. [Google Scholar] [CrossRef]
- Willis, K.J.; Bhagwat, S.A. Biodiversity and Climate Change. Science 2009, 326, 806–807. [Google Scholar] [CrossRef]
- Wolf, T.; Menne, B. Environment and Health Risks from Climate Change and Variability in Italy; World Health Organization, Regional Office for Europe: Copenhagen, Denmark, 2007. [Google Scholar]
- Chuah, L.O.; Effarizah, M.E.; Goni, A.M.; Rusul, G. Antibiotic Application and Emergence of Multiple Antibiotic Resistance (MAR) in Global Catfish Aquaculture. Curr. Environ. Health Rep. 2016, 3, 118–127. [Google Scholar] [CrossRef]
- Shao, Y.; Wang, Y.; Yuan, Y.; Xie, Y. A systematic review on antibiotics misuse in livestock and aquaculture and regulation implications in China. Sci. Total Environ. 2021, 798, 149205. [Google Scholar] [CrossRef]
- Türe, M.; Haliloglu, H.I.; Altuntas, C.; Boran, H.; Kutlu, I. Comparison of Experimental Susceptibility of Rainbow Trout (Oncorhynchus mykiss), Turbot (Psetta maxima), Black Sea Trout (Salmo trutta labrax) and Sea Bass (Dicentrarchus labrax) to Lactococcus garvieae. Turk. J. Fish. Aquat. Sci. 2014, 14, 507–513. [Google Scholar] [CrossRef]
- Vendrell, D.; Balcázar, J.L.; Ruiz-Zarzuela, I.; de Blas, I.; Gironés, O.; Múzquiz, J.L. Lactococcus garvieae in fish: A review. Comp. Immunol. Microbiol. Infect. Dis. 2006, 29, 177–198. [Google Scholar] [CrossRef]
- WHO. Critically Important Antimicrobials for Human Medicine, 6th Revision; WHO: Geneva, Switzerland, 2019; Available online: https://apps.who.int/iris/bitstream/handle/10665/312266/9789241515528-eng.pdf (accessed on 20 December 2023).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | LI296620A_23 | LI296620B_23 | LI329384_23 | BS000031_98 NO317338_20 TV252859_20 NO358429_21 PV358422_21 VR143326_21 NO346039_22 | SO223838_20 |
|---|---|---|---|---|---|
| Country of isolation | Italy | Italy | Italy | Italy | Italy |
| Year(s) of isolation | 2023 | 2023 | 2023 | 1998, 2020–2022 | 2020 |
| Farm Type | Inland saltwater | Inland saltwater | Offshore saltwater | Inland freshwater | Inland freshwater |
| Affected species | Sea bass | Sea bass | Sea bass | Rainbow trout | Rainbow trout |
| Mortality 1 | + | + | − | +++ | ++ |
| Gram | + | + | + | + | + |
| Morphology | Ovoid | Ovoid | Ovoid | Ovoid | Ovoid |
| Motility | − | − | − | − | − |
| Catalase | − | − | − | − | − |
| Oxidase | − | − | − | − | − |
| Oxidation (O)—Fermentation (F) | F | F | F | F | F |
| Citrate | − | − | − | − | − |
| DNase | − | − | − | − | − |
| Vogues-Proskauer | + | + | + | + | + |
| Esculin | + | + | + | + | + |
| Hippurate | − | − | − | + | + |
| Pyrrolidonyl arylamidase | + | + | + | + | + |
| α-galactosidase | − | − | − | − | − |
| β-glucuronidase | − | − | − | − | − |
| β-galactosidase | − | − | − | − | − |
| Phenylalanine ammonia lyase | − | − | − | − | − |
| Leucyl aminopeptidase | + | + | + | + | + |
| Arginine dihydrolase | + | + | - | + | + |
| Ribose | + | + | + | + | + |
| Arabinose | − | − | − | − | − |
| Mannitol | + | + | − | + | + |
| Sorbitol | − | − | − | − | − |
| Lactose | − | − | − | − | − |
| Trehalose | + | + | + | + | + |
| Inulin | − | − | − | − | − |
| Raffinose | − | − | − | − | − |
| Starch | − | − | + | − | − |
| Glycogen | − | − | − | − | − |
| β-haemolysis | − | − | − | − | − |
| API CODE | 5,143,110 | 5,143,110 | 5,143,011 | 7,143,110 | 7,143,110 |
| Strain | hlyIII 1 | Fbp 1 | Pva 1 | bsh1 1 | bsh2 1 |
|---|---|---|---|---|---|
| LI296620A_23 | + | + | + | − | − |
| LI296620B_23 | + | + | + | − | − |
| LI329384_23 | + | + | + | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salogni, C.; Bertasio, C.; Accini, A.; Gibelli, L.R.; Pigoli, C.; Susini, F.; Podavini, E.; Scali, F.; Varisco, G.; Alborali, G.L. The Characterisation of Lactococcus garvieae Isolated in an Outbreak of Septicaemic Disease in Farmed Sea Bass (Dicentrarchus labrax, Linnaues 1758) in Italy. Pathogens 2024, 13, 49. https://doi.org/10.3390/pathogens13010049
Salogni C, Bertasio C, Accini A, Gibelli LR, Pigoli C, Susini F, Podavini E, Scali F, Varisco G, Alborali GL. The Characterisation of Lactococcus garvieae Isolated in an Outbreak of Septicaemic Disease in Farmed Sea Bass (Dicentrarchus labrax, Linnaues 1758) in Italy. Pathogens. 2024; 13(1):49. https://doi.org/10.3390/pathogens13010049
Chicago/Turabian StyleSalogni, Cristian, Cristina Bertasio, Adelchi Accini, Lucia Rita Gibelli, Claudio Pigoli, Francesca Susini, Eleonora Podavini, Federico Scali, Giorgio Varisco, and Giovanni Loris Alborali. 2024. "The Characterisation of Lactococcus garvieae Isolated in an Outbreak of Septicaemic Disease in Farmed Sea Bass (Dicentrarchus labrax, Linnaues 1758) in Italy" Pathogens 13, no. 1: 49. https://doi.org/10.3390/pathogens13010049
APA StyleSalogni, C., Bertasio, C., Accini, A., Gibelli, L. R., Pigoli, C., Susini, F., Podavini, E., Scali, F., Varisco, G., & Alborali, G. L. (2024). The Characterisation of Lactococcus garvieae Isolated in an Outbreak of Septicaemic Disease in Farmed Sea Bass (Dicentrarchus labrax, Linnaues 1758) in Italy. Pathogens, 13(1), 49. https://doi.org/10.3390/pathogens13010049