
Kudoa septempunctata Spores Cause Acute Gastroenteric Symptoms in Mouse and Musk Shrew Models as Evidenced In Vitro in Human Colon Cells

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Ethics
2.2. Kudoa septempunctata Collection
2.3. Correlative Three-Dimensional (3-D) Quantitative Phase Imaging (QPI)
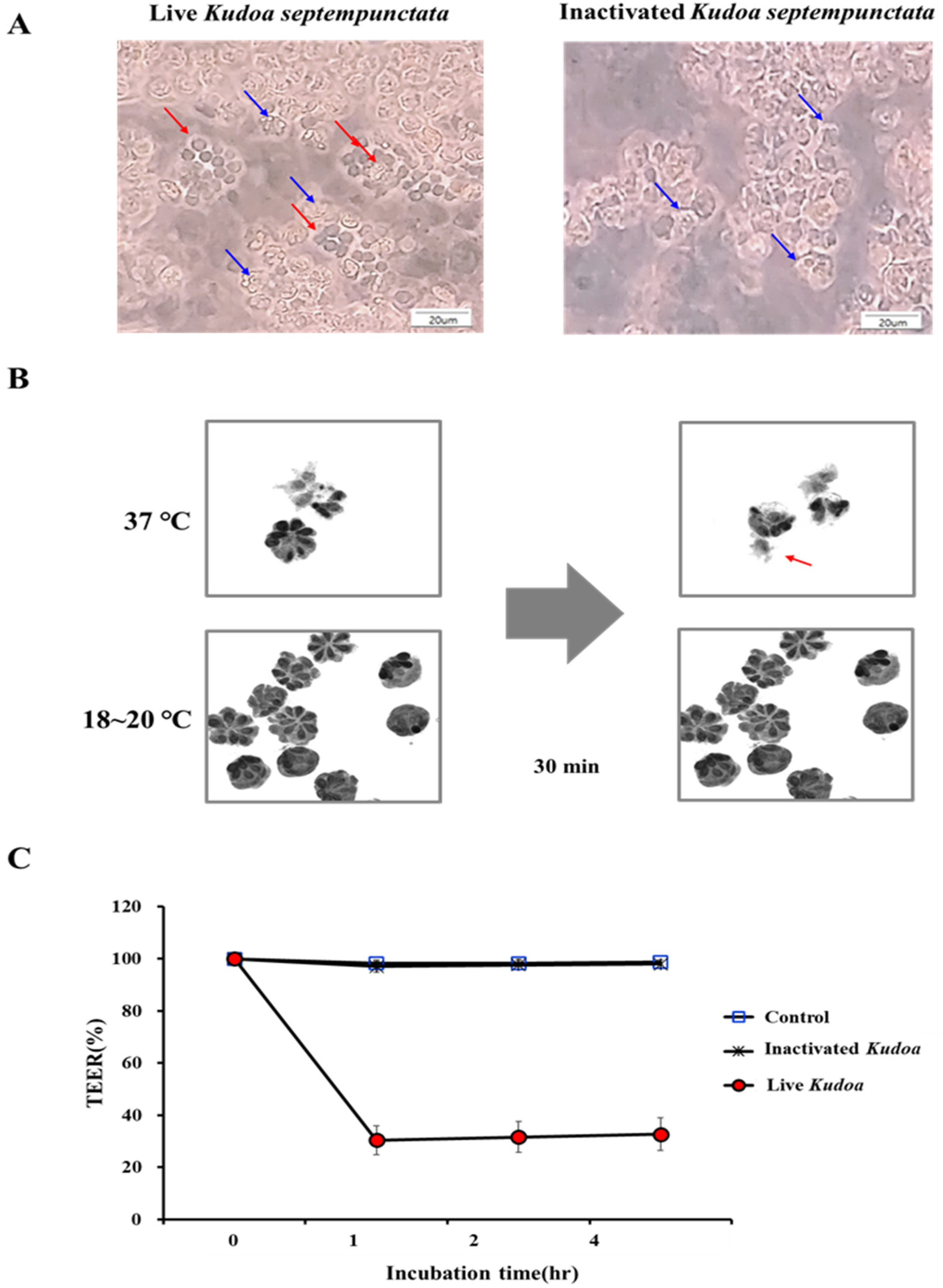
2.4. TEER Assay for Spore Activity
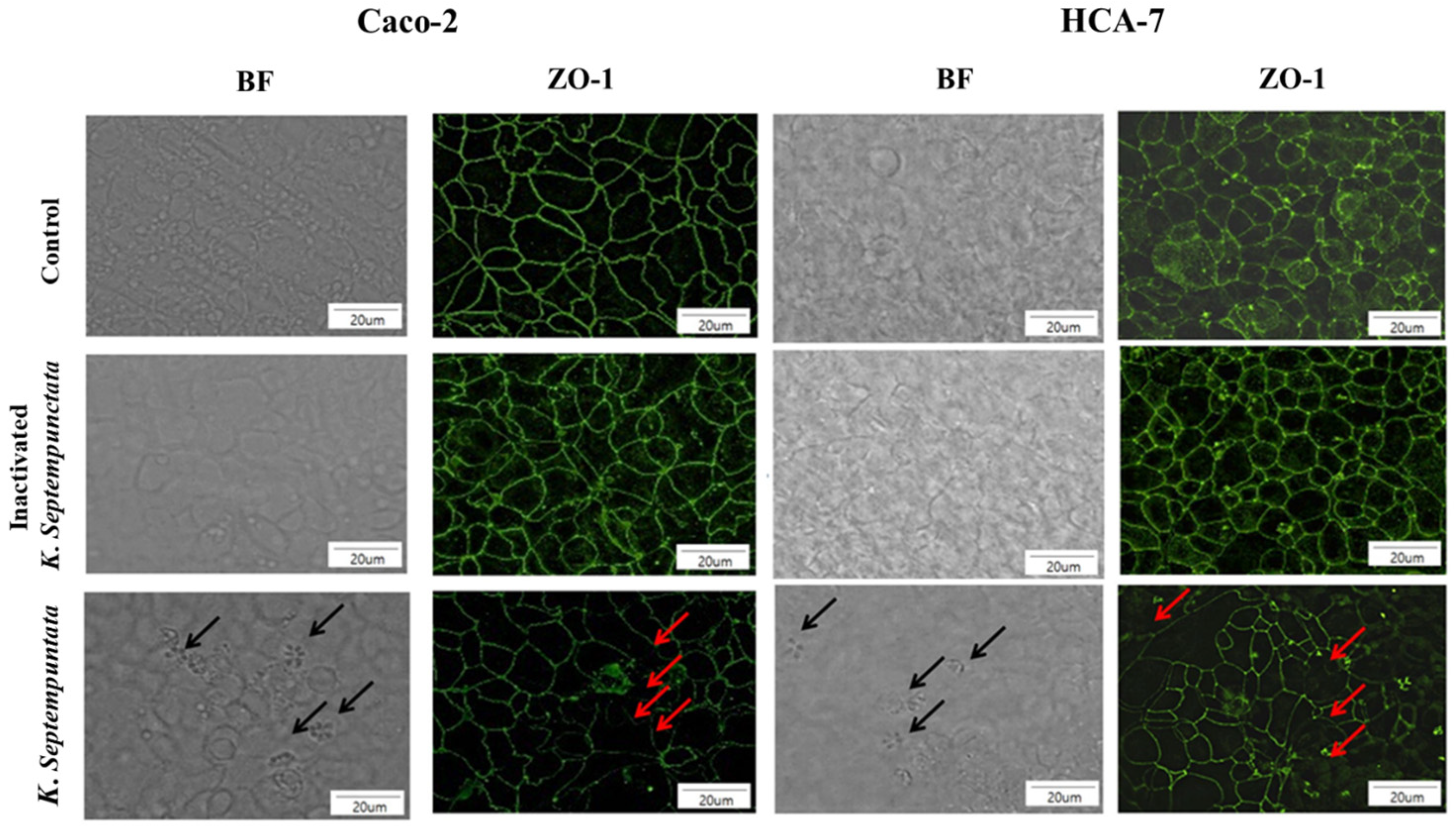
2.5. Immunofluorescence Assay on Zonula occludens-1 (ZO-1) in Cell-Line Cells
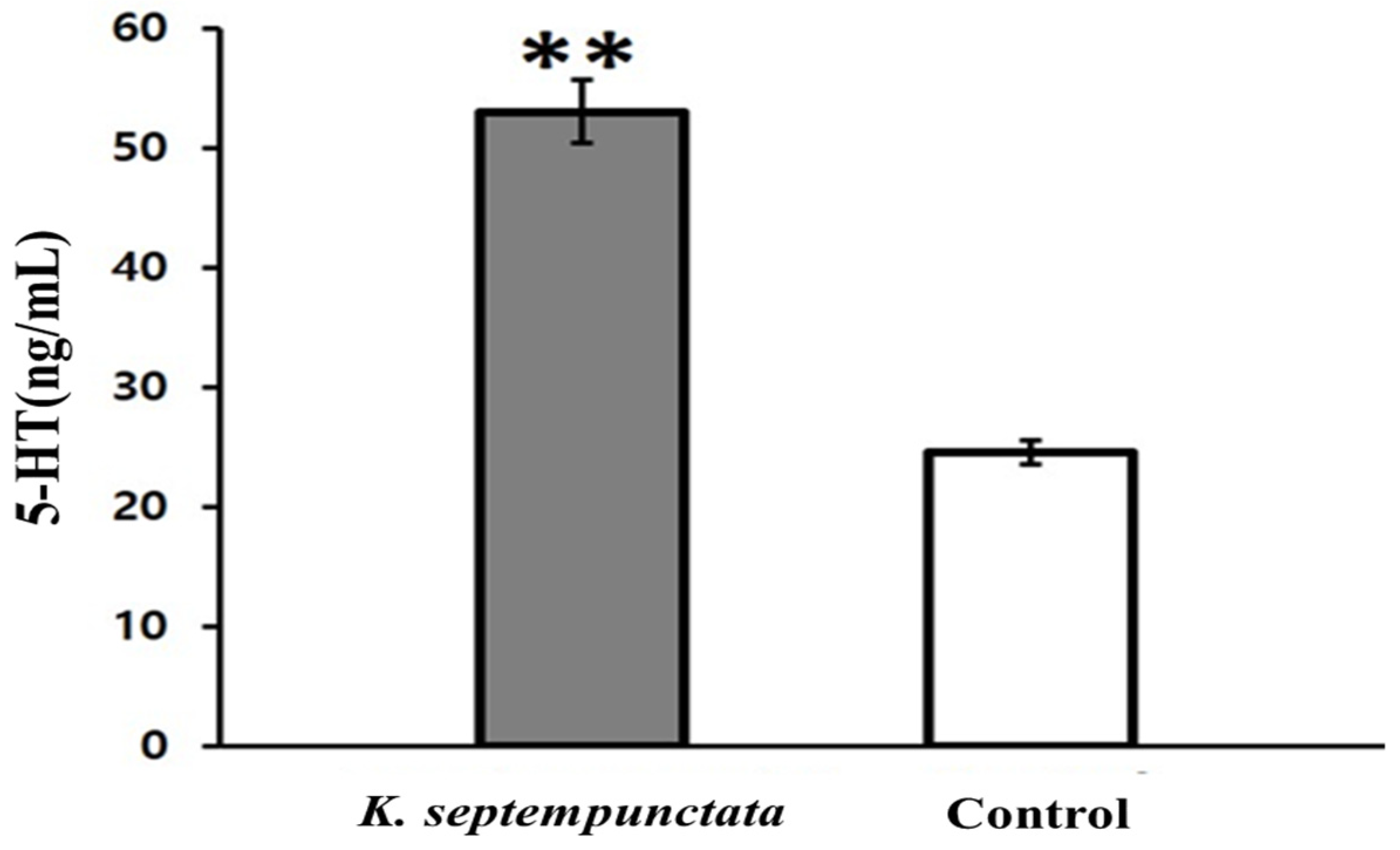
2.6. Assay on the Serotonin Production of Caco-2 Cells
2.7. Suckling Mouse Provocation for Diarrheal Response
2.8. Experimental Design for the House Musk Shrews to Determine Emetic Response to K. septempunctata
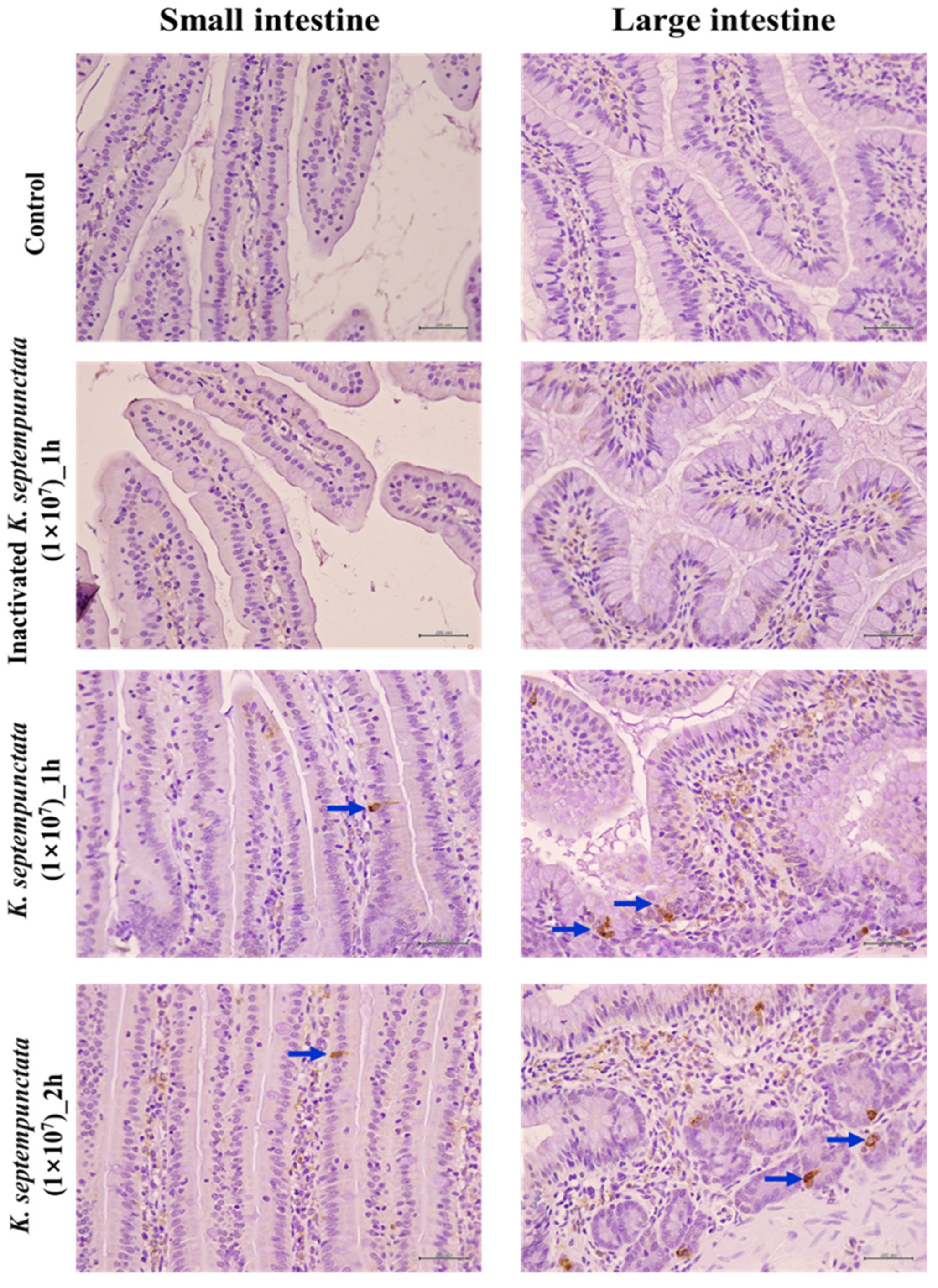
2.9. Immunohistochemical Localization of Serotonin in the Intestinal Tissues
2.10. Statistical Analysis
3. Results
3.1. Kudoa septempunctata Sporoplasm Altered the Transepithelial Electrical Resistance of the Caco-2 Cell Monolayer
3.2. K. septempunctata Disturbed Tight Junction ZO-1 Protein Arrangement
3.3. Diarrheal Responses of ddY and ICR Suckling Mice to K. septempunctata
3.4. Emetic Response to K. septempunctata in House Musk Shrews
3.5. Serotonin (5-HT) Produced In Vitro and In Vivo by K. septempunctata
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matsukane, Y.; Sato, H.; Tanaka, S.; Kamata, Y.; Sugita-Konishi, Y. Kudoa septempunctata n. sp. (Myxosporea: Multivalvulida) from an Aquacultured Olive Flounder (Paralichthys olivaceus) Imported from Korea. Parasitol. Res. 2010, 107, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, F.; Ogasawara, Y.; Kato, K.; Sekizuka, T.; Nozaki, T.; Sugita-Konishi, Y.; Ohnishi, T.; Kuroda, M. Genetic Variants of Kudoa septempunctata (Myxozoa: Multivalvulida), a Flounder Parasite Causing Foodborne Disease. J. Fish Dis. 2016, 39, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Chase, J.C.; Dawson-Coates, J.A.; Haddow, J.D.; Stewart, M.H.; Haines, L.R.; Whitaker, D.J.; Ken, M.L.; Olafson, R.W.; Pearson, T.W. Analysis of Kudoa thyrsites (Myxozoa: Myxosporea) Spore Antigens Using Monoclonal Antibodies. Dis. Aquat. Organ. 2001, 45, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Kawai, T.; Jinnai, M.; Ohnishi, T.; Sugita-Konishi, Y.; Kumeda, Y. Detection of Kudoa septempunctata 18S Ribosomal DNA in Patient Fecal Samples from Novel Food-Borne Outbreaks Caused by Consumption of Raw Olive Flounder (Paralichthys olivaceus). J. Clin. Microbiol. 2012, 50, 2964–2968. [Google Scholar] [CrossRef]
- Kawai, T.; Sekizuka, T.; Yahata, Y.; Kuroda, M.; Kumeda, Y.; Iijima, Y.; Kamata, Y.; Sugita-Konishi, Y.; Ohnishi, T. Identification of Kudoa septempunctata as the Causative Agent of Novel Food Poisoning Outbreaks in Japan by Consumption of Paralichthys olivaceus in Raw Fish. Clin. Infect. Dis. 2012, 54, 1046–1052. [Google Scholar] [CrossRef]
- Iwashita, Y.; Kamijo, Y.; Nakahashi, S.; Shindo, A.; Yokoyama, K.; Yamamoto, A.; Omori, Y.; Ishikura, K.; Fujioka, M.; Hatada, T.; et al. Food Poisoning Associated with Kudoa septempunctata. J. Emerg. Med. 2013, 44, 943–945. [Google Scholar] [CrossRef]
- Suzuki, J.; Murata, R.; Yokoyama, H.; Sadamasu, K.; Kai, A. Detection Rate of Diarrhoea-Causing Kudoa Hexapunctata in Pacific Bluefin Tuna Thunnus orientalis from Japanese Waters. Int. J. Food Microbiol. 2015, 194, 1–6. [Google Scholar] [CrossRef]
- Yokoyama, H.; Funaguma, N.; Kobayashi, S. In Vitro Inactivation of Kudoa septempunctata Spores Infecting the Muscle of Olive Flounder Paralichthys olivaceus. Foodborne Pathog. Dis. 2016, 13, 21–27. [Google Scholar] [CrossRef]
- Jang, Y.; Ahn, M.; Bang, H.; Kang, B. Effects of Kudoa septempunctata Genotype ST3 Isolate from Korea on ddY Suckling Mice. Parasite 2016, 23, 18. [Google Scholar] [CrossRef][Green Version]
- Ahn, M.; Woo, H.; Kang, B.; Jang, Y.; Shin, T. Effect of Oral Administration of Kudoa septempunctata Genotype ST3 in Adult BALB/C Mice. Parasite 2015, 22, 35. [Google Scholar] [CrossRef][Green Version]
- Guo, Q.; Jia, L.; Qin, J.; Li, H.; Gu, Z. Myxozoans and Our Dinner Table: Pathogenicity Studies of Myxobolus Honghuensis (Myxosporea: Bivalvulida) Using a Suckling Mice Model. Foodborne Pathog. Dis. 2015, 12, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, T.; Kikuchi, Y.; Furusawa, H.; Kamata, Y.; Sugita-Konishi, Y. Kudoa septempunctata Invasion Increases the Permeability of Human Intestinal Epithelial Monolayer. Foodborne Pathog. Dis. 2013, 10, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.; Ko, H.J.; Kim, J.; Jang, Y.; Shin, T. Evaluation of the Inflammatory Response to Kudoa septempunctata Genotype ST3 Isolated from Olive Flounder (Paralichthys olivaceus) in Caco-2 Cells. Parasite 2018, 25, 12. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, T.; Oyama, R.; Furusawa, H.; Ohba, N.; Kamata, Y.; Sugita-Konishi, Y. Kudoa septempunctata Was Recognised by Toll-Like Receptor 2 Produced by a RAW 264 Macrophage-Like Cell Line. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2013, 30, 1365–1369. [Google Scholar] [CrossRef]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER Measurement Techniques for In Vitro Barrier Model Systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef]
- Shin, J.H.; Yang, J.P.; Seo, S.H.; Kim, S.G.; Kim, E.M.; Ham, D.W.; Shin, E.H. Immune-Triggering Effect of the Foodborne Parasite Kudoa septempunctata through the C-Type Lectin Mincle in HT29 Cells. BMB Rep. 2020, 53, 478–483. [Google Scholar] [CrossRef]
- Yamamoto, S.; Kawano, F.; Yokoyama, H.; Kobayashi, S. Effects of Kudoa septempunctata Infections in a Human Intestinal Epithelial Model (Caco-2): A DNA Microarray Study. Biosci. Biotechnol. Biochem. 2020, 84, 1030–1038. [Google Scholar] [CrossRef]
- Shin, S.; Kim, K.; Yoon, J.; Park, Y. Active Illumination Using a Digital Micromirror Device for Quantitative Phase Imaging. Opt. Lett. 2015, 40, 5407–5410. [Google Scholar] [CrossRef]
- Fedchenko, N.; Reifenrath, J. Different Approaches for Interpretation and Reporting of Immunohistochemistry Analysis Results in the Bone Tissue—A Review. Diagn. Pathol. 2014, 9, 221. [Google Scholar] [CrossRef]
- Samadi, N.; Klems, M.; Untersmayr, E. The Role of Gastrointestinal Permeability in Food Allergy. Ann. Allergy Asthma Immunol. 2018, 121, 168–173. [Google Scholar] [CrossRef]
- König, J.; Wells, J.; Cani, P.D.; García-Ródenas, C.L.; MacDonald, T.; Mercenier, A.; Whyte, J.; Troost, F.; Brummer, R.J. Human Intestinal Barrier Function in Health and Disease. Clin. Transl. Gastroenterol. 2016, 7, e196. [Google Scholar] [CrossRef] [PubMed]
- Ventura, M.T.; Polimeno, L.; Amoruso, A.C.; Gatti, F.; Annoscia, E.; Marinaro, M.; Di Leo, E.; Matino, M.G.; Buquicchio, R.; Bonini, S.; et al. Intestinal Permeability in Patients with Adverse Reactions to Food. Dig. Liver Dis. 2006, 38, 732–736. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Ryu, S.; Lee, H. Foodborne Illness Outbreaks in Gyeonggi Province, Korea, Following Seafood Consumption Potentially Caused by Kudoa septempunctata between 2015 and 2016. Osong Public Health Res. Perspect. 2018, 9, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Yahata, Y.; Sugita-Konishi, Y.; Ohnishi, T.; Toyokawa, T.; Nakamura, N.; Taniguchi, K.; Okabe, N. Kudoa septempunctata-Induced Gastroenteritis in Humans after Flounder Consumption in Japan: A Case-Controlled Study. Jpn. J. Infect. Dis. 2015, 68, 119–123. [Google Scholar] [CrossRef]
- Tachibana, T.; Watari, T. Kudoa septempunctata Infection: An Underdiagnosed Pathogen of Acute Gastrointestinal Symptoms. QJM Int. J. Med. 2020, 113, 43–44. [Google Scholar] [CrossRef]
- Lytton, S.D.; Fischer, W.; Nagel, W.; Haas, R.; Beck, F.X. Production of Ammonium by Helicobacter pylori Mediates Occludin Processing and Disruption of Tight Junctions in Caco-2 Cells. Microbiology 2005, 151, 3267–3276. [Google Scholar] [CrossRef]
- Dickman, K.G.; Hempson, S.J.; Anderson, J.; Lippe, S.; Zhao, L.; Burakoff, R.; Shaw, R.D. Rotavirus Alters Paracellular Permeability and Energy Metabolism in Caco-2 Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G757–G766. [Google Scholar] [CrossRef]
- Carballeda-Sangiao, N.; Sánchez-Alonso, I.; Navas, A.; Arcos, S.C.; de Palencia, P.F.; Careche, M.; González-Muñoz, M. Anisakis simplex Products Impair Intestinal Epithelial Barrier Function and Occludin and Zonula occludens-1 Localisation in Differentiated Caco-2 Cells. PLOS Negl. Trop. Dis. 2020, 14, e0008462. [Google Scholar] [CrossRef]
- El-Matbouli, M.; Hoffmann, R.W. Light and Electron Microscopic Studies on the Chronological Development of Myxobolus cerebralis to the Actinosporean Stage in Tubifex Tubifex. Int. J. Parasitol. 1998, 28, 195–217. [Google Scholar] [CrossRef]
- Guttman, J.A.; Finlay, B.B. Tight Junctions as Targets of Infectious Agents. Biochim. Biophys. Acta 2009, 1788, 832–841. [Google Scholar] [CrossRef]
- Viswanathan, V.K.; Hodges, K.; Hecht, G. Enteric Infection Meets Intestinal Function: How Bacterial Pathogens Cause Diarrhoea. Nat. Rev. Microbiol. 2009, 7, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Field, M. Intestinal Ion Transport and the Pathophysiology of Diarrhea. J. Clin. Investig. 2003, 111, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Hoque, K.M.; Chakraborty, S.; Sheikh, I.A.; Woodward, O.M. New Advances in the Pathophysiology of Intestinal Ion Transport and Barrier Function in Diarrhea and the Impact on Therapy. Expert Rev. Anti. Infect. Ther. 2012, 10, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Vogelmann, R.; Amieva, M.R.; Falkow, S.; Nelson, W.J. Breaking into the Epithelial Apical-Junctional Complex--News from Pathogen Hackers. Curr. Opin. Cell Biol. 2004, 16, 86–93. [Google Scholar] [CrossRef]
- Kumar, A.; Chatterjee, I.; Anbazhagan, A.N.; Jayawardena, D.; Priyamvada, S.; Alrefai, W.A.; Sun, J.; Borthakur, A.; Dudeja, P.K. Cryptosporidium parvum Disrupts Intestinal Epithelial Barrier Function via Altering Expression of Key Tight Junction and Adherens Junction Proteins. Cell. Microbiol. 2018, 20, e12830. [Google Scholar] [CrossRef]
- Buret, A.G.; Mitchell, K.; Muench, D.G.; Scott, K.G. Giardia lamblia Disrupts Tight Junctional ZO-1 and Increases Permeability in Non-transformed Human Small Intestinal Epithelial Monolayers: Effects of Epidermal Growth Factor. Parasitology 2002, 125, 11–19. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms Regulating Intestinal Barrier Integrity and Its Pathological Implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef]
- Rao, M.; Gershon, M.D. The Bowel and beyond: The Enteric Nervous System in Neurological Disorders. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 517–528. [Google Scholar] [CrossRef]
- Bertrand, P.P.; Kunze, W.A.; Furness, J.B.; Bornstein, J.C. The Terminals of Myenteric Intrinsic Primary Afferent Neurons of the Guinea-Pig Ileum Are Excited by 5-Hydroxytryptamine Acting at 5-hydroxytryptamine-3 Receptors. Neuroscience 2000, 101, 459–469. [Google Scholar] [CrossRef]
- Hansen, M.B.; Witte, A.B. The Role of Serotonin in Intestinal Luminal Sensing and Secretion. Acta Physiol. 2008, 193, 311–323. [Google Scholar] [CrossRef]
- Spohn, S.N.; Mawe, G.M. Non-conventional Features of Peripheral Serotonin Signalling—The Gut and Beyond. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.B. The Enteric Nervous System I: Organisation and Classification. Pharmacol. Toxicol. 2003, 92, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Hagbom, M.; Istrate, C.; Engblom, D.; Karlsson, T.; Rodriguez-Diaz, J.; Buesa, J.; Taylor, J.A.; Loitto, V.M.; Magnusson, K.E.; Ahlman, H.; et al. Rotavirus Stimulates Release of Serotonin (5-HT) from Human Enterochromaffin Cells and Activates Brain Structures Involved in Nausea and Vomiting. PLOS Pathog. 2011, 7, e1002115. [Google Scholar] [CrossRef] [PubMed]
- Sugita-Konishi, Y.; Sato, H.; Ohnishi, T. Novel Foodborne Disease Associated with Consumption of Raw Fish, Olive Flounder (Paralichthys olivaceus). J. Food Saf. 2014, 2, 141–150. [Google Scholar] [CrossRef]
- Terry, N.; Margolis, K.G. Serotonergic Mechanisms Regulating the GI Tract: Experimental Evidence and Therapeutic Relevance. Handb. Exp. Pharmacol. 2017, 239, 319–342. [Google Scholar] [CrossRef]
- Margolis, K.G.; Stevanovic, K.; Li, Z.; Yang, Q.M.; Oravecz, T.; Zambrowicz, B.; Jhaver, K.G.; Diacou, A.; Gershon, M.D. Pharmacological Reduction of Mucosal but Not Neuronal Serotonin Opposes Inflammation in Mouse Intestine. Gut 2014, 63, 928–937. [Google Scholar] [CrossRef]
- Kim, J.J.; Wang, H.; Terc, J.D.; Zambrowicz, B.; Yang, Q.M.; Khan, W.I. Blocking Peripheral Serotonin Synthesis by Telotristat Etiprate (LX1032/LX1606) Reduces Severity of Both Chemical- and Infection-Induced Intestinal Inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G455–G465. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| K. septempunctata Spores Administered. | No. of Diarrhea/Mouse Tested | No. of Diarrheal Incidents | Duration of Diarrhea (min) |
|---|---|---|---|
| Live 2 × 106 | 8/10 *** | 3.3 | 115.0 ± 23.9 |
| Live 1 × 105 | 4/10 | 1.3 | 119.0 ± 32.8 |
| Heat-inactivated 2 × 106 | 0/10 | _ | _ |
| PBS | 1/10 | 1 | 254 |
| K. septempunctata Spores Administered | No. of Diarrhea/Mouse Tested | No. of Diarrheal Incidents | p-Value |
|---|---|---|---|
| Live 2 × 106 | 10/10 | 4.3 | 0.0001 *** |
| Live 2 × 105 | 8/10 | 2.4 | 0.0006 *** |
| Live 2 × 104 | 1/5 | 0.4 | 0.555 |
| Live 2 × 103 | 1/5 | 0.4 | 0.555 |
| Live 2 × 102 | 0/5 | 0 | - |
| Heat-inactivated 2 × 106 | 0/5 | 0.1 | - |
| PBS | 1/10 | 0.1 | - |
| K. septempunctata Spores Administered | No. of Musk Shrews (Emesis/Tested) | No. of Emetic Events | Duration of Emesis (min) |
|---|---|---|---|
| Live 1 × 107 | 6/7 *** | 4.0 ± 1.2 | 30.0 ± 5.8 |
| Live 1 × 106 | 4/8 | 3.0 ± 1.0 | 23.5 ± 2.5 |
| Heat-inactivated 1 × 107 | 0/8 | 0 | 0 |
| PBS | 0/2 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, S.-H.; Kwon, J.-Y.; Lee, S.-O.; Lee, H.-I.; Hong, S.-J.; Ju, J.-W. Kudoa septempunctata Spores Cause Acute Gastroenteric Symptoms in Mouse and Musk Shrew Models as Evidenced In Vitro in Human Colon Cells. Pathogens 2023, 12, 739. https://doi.org/10.3390/pathogens12050739
Hong S-H, Kwon J-Y, Lee S-O, Lee H-I, Hong S-J, Ju J-W. Kudoa septempunctata Spores Cause Acute Gastroenteric Symptoms in Mouse and Musk Shrew Models as Evidenced In Vitro in Human Colon Cells. Pathogens. 2023; 12(5):739. https://doi.org/10.3390/pathogens12050739
Chicago/Turabian StyleHong, Sung-Hee, Ji-Young Kwon, Soon-Ok Lee, Hee-Il Lee, Sung-Jong Hong, and Jung-Won Ju. 2023. "Kudoa septempunctata Spores Cause Acute Gastroenteric Symptoms in Mouse and Musk Shrew Models as Evidenced In Vitro in Human Colon Cells" Pathogens 12, no. 5: 739. https://doi.org/10.3390/pathogens12050739
APA StyleHong, S.-H., Kwon, J.-Y., Lee, S.-O., Lee, H.-I., Hong, S.-J., & Ju, J.-W. (2023). Kudoa septempunctata Spores Cause Acute Gastroenteric Symptoms in Mouse and Musk Shrew Models as Evidenced In Vitro in Human Colon Cells. Pathogens, 12(5), 739. https://doi.org/10.3390/pathogens12050739