


Glycerol Monolaurate Inhibits Wild-Type African Swine Fever Virus Infection in Porcine Macrophages



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell and Virus Preparations
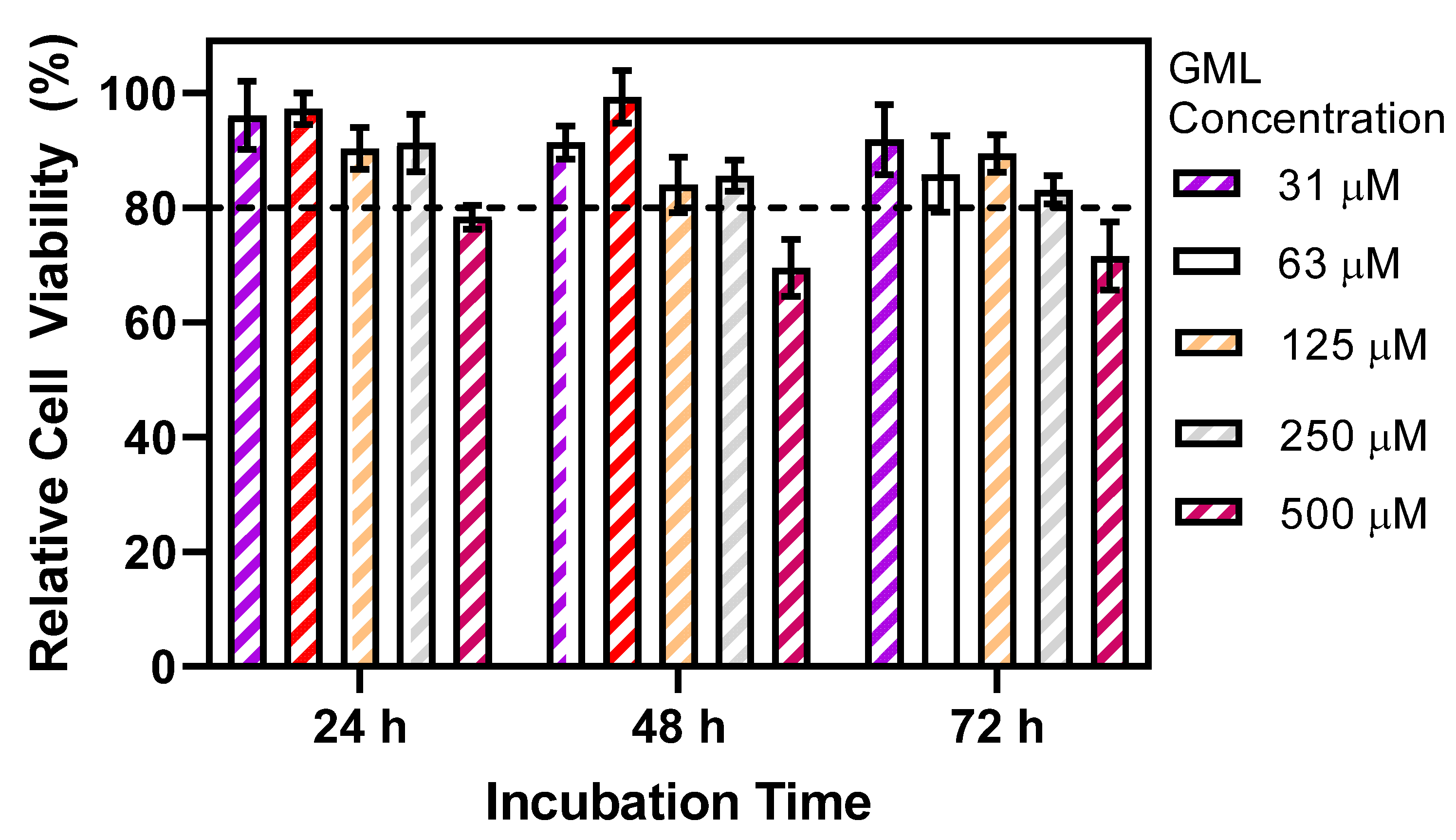
2.2. Cytotoxicity Assay
2.3. Antiviral Assay
2.4. Statistical Analysis
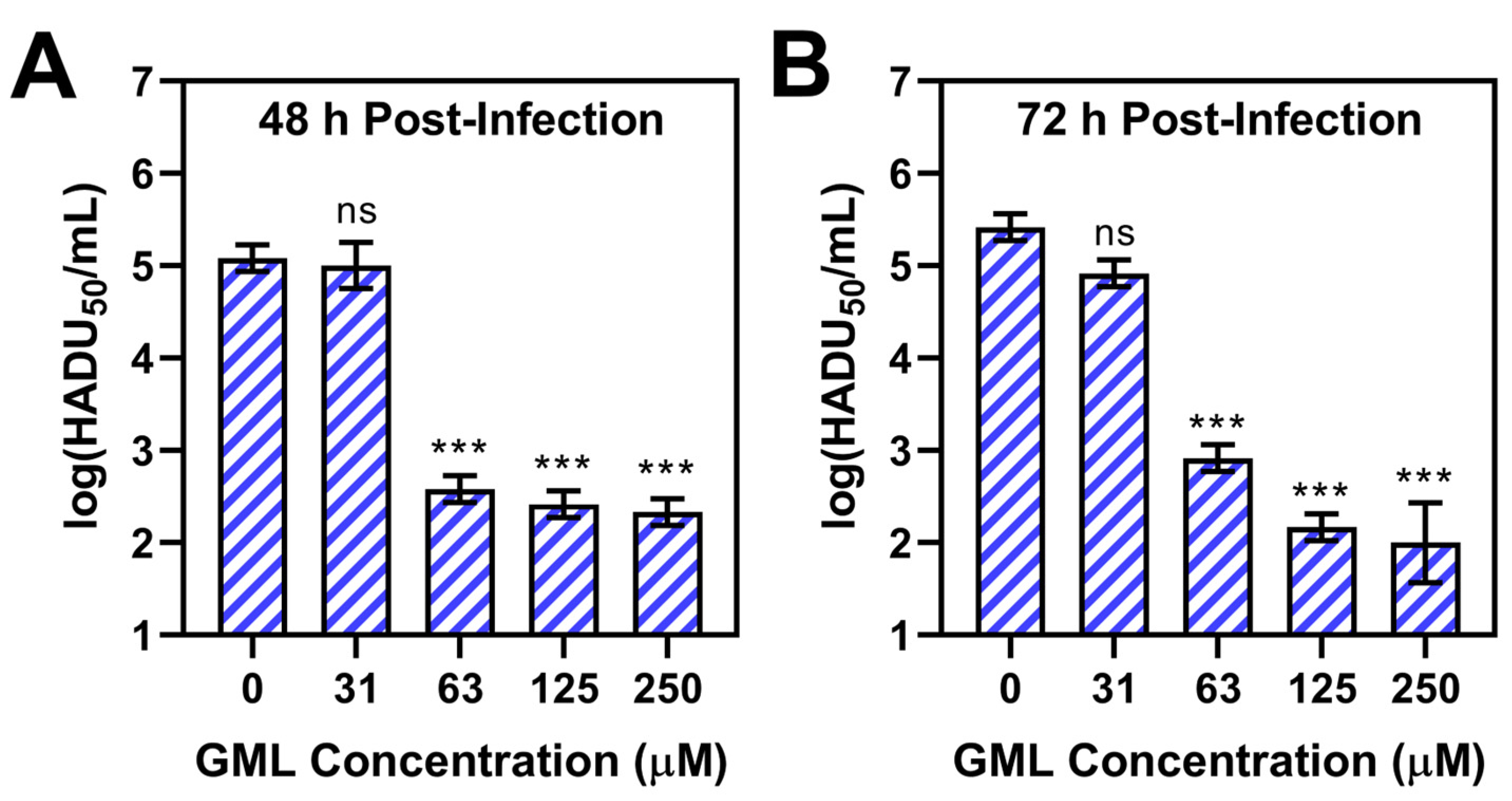
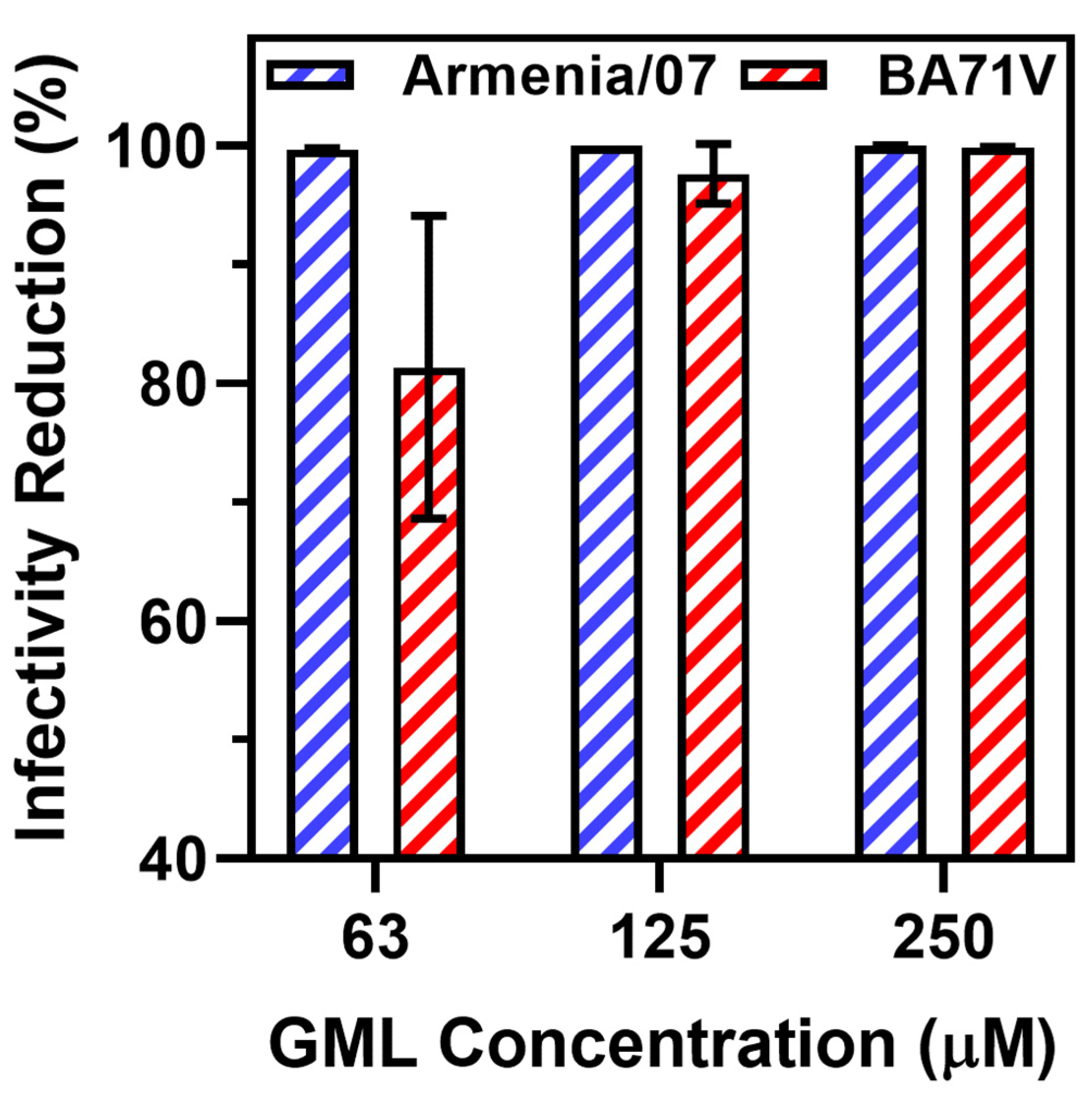
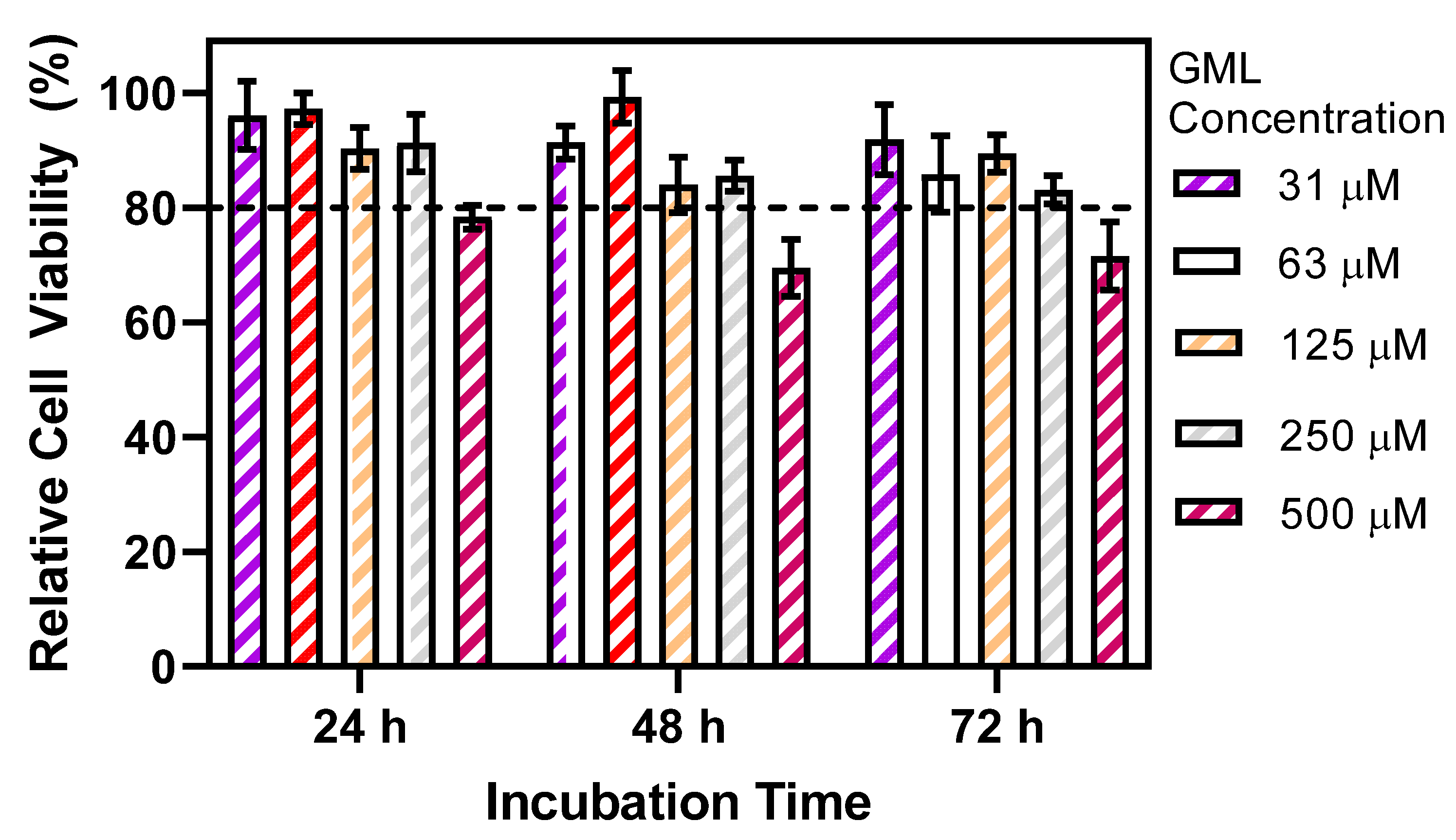
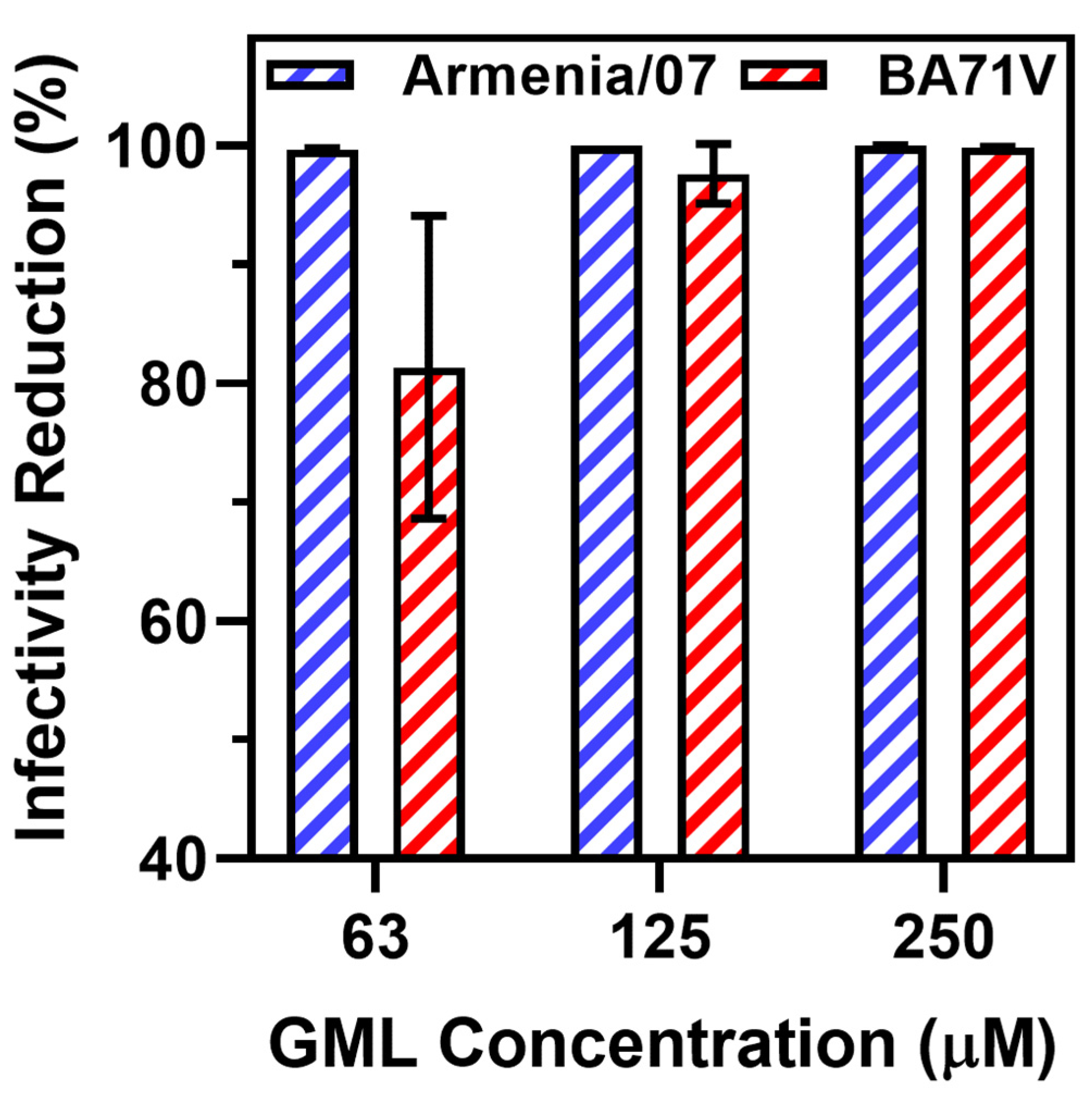
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galindo, I.; Alonso, C. African swine fever virus: A review. Viruses 2017, 9, 103. [Google Scholar] [CrossRef] [PubMed]
- Gaudreault, N.N.; Madden, D.W.; Wilson, W.C.; Trujillo, J.D.; Richt, J.A. African swine fever virus: An emerging DNA arbovirus. Front. Vet. Sci. 2020, 7, 215. [Google Scholar] [CrossRef] [PubMed]
- You, S.; Liu, T.; Zhang, M.; Zhao, X.; Dong, Y.; Wu, B.; Wang, Y.; Li, J.; Wei, X.; Shi, B. African swine fever outbreaks in China led to gross domestic product and economic losses. Nat. Food 2021, 2, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Arabyan, E.; Kotsynyan, A.; Hakobyan, A.; Zakaryan, H. Antiviral agents against African swine fever virus. Virus Res. 2019, 270, 197669. [Google Scholar] [CrossRef]
- Penrith, M.L.; Kivaria, F.M. One hundred years of African swine fever in Africa: Where have we been, where are we now, where are we going? Transbound. Emerg. Dis. 2022, 69, e1179–e1200. [Google Scholar] [CrossRef]
- Barongo, M.B.; Bishop, R.P.; Fèvre, E.M.; Knobel, D.L.; Ssematimba, A. A mathematical model that simulates control options for African swine fever virus (ASFV). PLoS ONE 2016, 11, e0158658. [Google Scholar] [CrossRef]
- Mutua, F.; Dione, M. The context of application of biosecurity for control of African swine fever in smallholder pig systems: Current gaps and recommendations. Front. Vet. Sci. 2021, 8, 689811. [Google Scholar] [CrossRef]
- Beato, M.S.; D’Errico, F.; Iscaro, C.; Petrini, S.; Giammarioli, M.; Feliziani, F. Disinfectants against African swine fever: An updated review. Viruses 2022, 14, 1384. [Google Scholar] [CrossRef]
- Niederwerder, M.C.; Stoian, A.M.; Rowland, R.R.; Dritz, S.S.; Petrovan, V.; Constance, L.A.; Gebhardt, J.T.; Olcha, M.; Jones, C.K.; Woodworth, J.C. Infectious dose of African swine fever virus when consumed naturally in liquid or feed. Emerg. Infect. Dis. 2019, 25, 891. [Google Scholar] [CrossRef]
- Phillips, F.C.; Rubach, J.K.; Poss, M.J.; Anam, S.; Goyal, S.M.; Dee, S.A. Monoglyceride reduces viability of porcine epidemic diarrhoea virus in feed and prevents disease transmission to post-weaned piglets. Transbound. Emerg. Dis. 2022, 69, 121–127. [Google Scholar] [CrossRef]
- Śmiechowska, M.; Newerli-Guz, J.; Skotnicka, M. Spices and seasoning mixes in European Union—Innovations and ensuring safety. Foods 2021, 10, 2289. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhao, D.; Wang, J.; Zhang, Y.; Wang, M.; Gao, Y.; Li, F.; Wang, J.; Bu, Z.; Rao, Z.; et al. Architecture of African swine fever virus and implications for viral assembly. Science 2019, 366, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Vigant, F.; Santos, N.C.; Lee, B. Broad-spectrum antivirals against viral fusion. Nat. Rev. Microbiol. 2015, 13, 426–437. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jeon, W.-Y.; Sut, T.N.; Cho, N.-J.; Jackman, J.A. Stopping membrane-enveloped viruses with nanotechnology strategies: Toward antiviral drug development and pandemic preparedness. ACS Nano 2021, 15, 125–148. [Google Scholar] [CrossRef] [PubMed]
- Hernáez, B.; Guerra, M.; Salas, M.L.; Andrés, G. African swine fever virus undergoes outer envelope disruption, capsid disassembly and inner envelope fusion before core release from multivesicular endosomes. PLoS Pathog. 2016, 12, e1005595. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.M.; Moreno, L.T.; Alejo, A.; Lacasta, A.; Rodríguez, F.; Salas, M.L. Genome sequence of African swine fever virus BA71, the virulent parental strain of the nonpathogenic and tissue-culture adapted BA71V. PLoS ONE 2015, 10, e0142889. [Google Scholar] [CrossRef]
- Jackman, J.A.; Boyd, R.D.; Elrod, C.C. Medium-chain fatty acids and monoglycerides as feed additives for pig production: Towards gut health improvement and feed pathogen mitigation. J. Anim. Sci. Biotechnol. 2020, 11, 44. [Google Scholar] [CrossRef]
- Jackman, J.A.; Lavergne, T.A.; Elrod, C.C. Antimicrobial monoglycerides for swine and poultry applications. Front. Anim. Sci. 2022, 3, 1019320. [Google Scholar] [CrossRef]
- Hakobyan, A.; Galindo, I.; Nañez, A.; Arabyan, E.; Karalyan, Z.; Chistov, A.A.; Streshnev, P.P.; Korshun, V.A.; Alonso, C.; Zakaryan, H. Rigid amphipathic fusion inhibitors demonstrate antiviral activity against African swine fever virus. J. Gen. Virol. 2018, 99, 148–156. [Google Scholar] [CrossRef]
- Jackman, J.A.; Hakobyan, A.; Zakaryan, H.; Elrod, C.C. Inhibition of African swine fever virus in liquid and feed by medium-chain fatty acids and glycerol monolaurate. J. Anim. Sci. Biotechnol. 2020, 11, 114. [Google Scholar] [CrossRef]
- Niederwerder, M.C.; Dee, S.; Diel, D.G.; Stoian, A.M.M.; Constance, L.A.; Olcha, M.; Petrovan, V.; Patterson, G.; Cino-Ozuna, A.G.; Rowland, R.R.R. Mitigating the risk of African swine fever virus in feed with anti-viral chemical additives. Transbound. Emerg. Dis. 2021, 68, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Palowski, A. The Use of a Risk-Free In Situ Non-Animal (RISNA) Surrogate Assay for Evaluating Inactivation Strategies of African Swine Fever Virus in Feed Ingredients in Real-World Demonstrations. Master’s Thesis, University of Minnesota, Minneapolis, MN, USA, 2021. [Google Scholar]
- Palowski, A.; Balestreri, C.; Urriola, P.E.; van de Ligt, J.L.G.; Sampedro, F.; Dee, S.; Shah, A.; Yancy, H.F.; Shurson, G.C.; Schroeder, D.C. Survival of a surrogate African swine fever virus-like algal virus in feed matrices using a 23-day commercial United States truck transport model. Front. Microbiol. 2022, 13, 1059118. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhu, L.; Zhao, X.; Liu, J.; Cheng, H.; Zhang, L.; Tang, H.; Sun, X.; Hu, Y.; Xu, Z. Effects of oral of administration of monoglycide laurate on virus load and inflammation in PEDV infected porcine. Front. Vet. Sci. 2022, 9, 98038. [Google Scholar] [CrossRef] [PubMed]
- Su, B.; Wang, Y.; Jian, S.; Tang, H.; Deng, H.; Zhu, L.; Zhao, X.; Liu, J.; Cheng, H.; Zhang, L.; et al. In vitro and in vivo antiviral activity of monolaurin against Seneca Valley virus. Front. Vet. Sci. 2023, 10, 980187. [Google Scholar] [CrossRef] [PubMed]
- Arabyan, E.; Hakobyan, A.; Hakobyan, T.; Grigoryan, R.; Izmailyan, R.; Avetisyan, A.; Karalyan, Z.; Jackman, J.A.; Ferreira, F.; Elrod, C.C.; et al. Flavonoid library screening reveals kaempferol as a potential antiviral agent against African swine fever virus. Front. Microbiol. 2021, 12, 736780. [Google Scholar] [CrossRef]
- Carrascosa, A.L.; Bustos, M.J.; de Leon, P. Methods for growing and titrating African swine fever virus: Field and laboratory samples. Curr. Protoc. Cell Biol. 2011, 53, 26.14.1–26.14.25. [Google Scholar] [CrossRef]
- Kabara, J.J.; Swieczkowski, D.M.; Conley, A.J.; Truant, J.P. Fatty acids and derivatives as antimicrobial agents. Antimicrob. Agents Chemother. 1972, 2, 23–28. [Google Scholar] [CrossRef]
- Welch, J.L.; Xiang, J.; Okeoma, C.M.; Schlievert, P.M.; Stapleton, J.T. Glycerol monolaurate, an analogue to a factor secreted by lactobacillus, is virucidal against enveloped viruses, including HIV-1. mBio 2020, 11, 10–1128. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Kim, M.C.; Cho, N.-J. Spectrum of membrane morphological responses to antibacterial fatty acids and related surfactants. Langmuir 2015, 31, 10223–10232. [Google Scholar] [CrossRef]
- Hierholzer, J.C.; Kabara, J.J. In vitro effects of monolaurin compounds on enveloped RNA and DNA viruses. J. Food Saf. 1982, 4, 1–12. [Google Scholar] [CrossRef]
- Rowlands, R.J.; Michaud, V.; Heath, L.; Hutchings, G.; Oura, C.; Vosloo, W.; Dwarka, R.; Onashvili, T.; Albina, E.; Dixon, L.K. African swine fever virus isolate, Georgia, 2007. Emerg. Infect. Dis. 2008, 14, 1870. [Google Scholar] [CrossRef] [PubMed]
- Sunwoo, S.-Y.; Pérez-Núñez, D.; Morozov, I.; Sánchez, E.G.; Gaudreault, N.N.; Trujillo, J.D.; Mur, L.; Nogal, M.; Madden, D.; Urbaniak, K.; et al. DNA-protein vaccination strategy does not protect from challenge with African swine fever virus Armenia 2007 strain. Vaccines 2019, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.K.; Park, S.; Ma, G.J.; Kolahdouzan, K.; Zhdanov, V.P.; Jackman, J.A.; Cho, N.-J. Competing interactions of fatty acids and monoglycerides trigger synergistic phospholipid membrane remodeling. J. Phys. Chem. Lett. 2020, 11, 4951–4957. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.W.; Jeon, W.-Y.; Yoon, B.K.; Jackman, J.A. Mechanistic evaluation of antimicrobial lipid interactions with tethered lipid bilayers by electrochemical impedance spectroscopy. Sensors 2022, 22, 3712. [Google Scholar] [CrossRef]
- Yang, L.; Wen, J.; Zhang, Y.; Liu, Z.; Luo, Z.; Xu, L.; Lai, S.; Tang, H.; Sun, X.; Hu, Y.; et al. The antiviral activity of caprylic monoglyceride against porcine reproductive and respiratory syndrome virus in vitro and in vivo. Molecules 2022, 27, 7263. [Google Scholar] [CrossRef]
- Vigant, F.; Hollmann, A.; Lee, J.; Santos, N.C.; Jung, M.E.; Lee, B. The rigid amphipathic fusion inhibitor dUY11 acts through photosensitization of viruses. J. Virol. 2014, 88, 1849–1853. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Park, S.; Mokrzecka, N.; Cho, N.-J. Characterizing the membrane-disruptive behavior of dodecylglycerol using supported lipid bilayers. Langmuir 2019, 35, 3568–3575. [Google Scholar] [CrossRef]
- Farcet, J.-B.; Karbiener, M.; Zelger, L.; Kindermann, J.; Kreil, T.R. Detergent-mediated virus inactivation in biotechnological matrices: More than just CMC. Int. J. Mol. Sci. 2023, 24, 7920. [Google Scholar] [CrossRef]
- Tan, J.Y.B.; Yoon, B.K.; Cho, N.-J.; Lovrić, J.; Jug, M.; Jackman, J.A. Lipid nanoparticle technology for delivering biologically active fatty acids and monoglycerides. Int. J. Mol. Sci. 2021, 22, 9664. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jackman, J.A.; Arabyan, E.; Zakaryan, H.; Elrod, C.C. Glycerol Monolaurate Inhibits Wild-Type African Swine Fever Virus Infection in Porcine Macrophages. Pathogens 2023, 12, 1193. https://doi.org/10.3390/pathogens12101193
Jackman JA, Arabyan E, Zakaryan H, Elrod CC. Glycerol Monolaurate Inhibits Wild-Type African Swine Fever Virus Infection in Porcine Macrophages. Pathogens. 2023; 12(10):1193. https://doi.org/10.3390/pathogens12101193
Chicago/Turabian StyleJackman, Joshua A., Erik Arabyan, Hovakim Zakaryan, and Charles C. Elrod. 2023. "Glycerol Monolaurate Inhibits Wild-Type African Swine Fever Virus Infection in Porcine Macrophages" Pathogens 12, no. 10: 1193. https://doi.org/10.3390/pathogens12101193
APA StyleJackman, J. A., Arabyan, E., Zakaryan, H., & Elrod, C. C. (2023). Glycerol Monolaurate Inhibits Wild-Type African Swine Fever Virus Infection in Porcine Macrophages. Pathogens, 12(10), 1193. https://doi.org/10.3390/pathogens12101193