
Assessing the Epidemiology of Rotavirus A, B, C and H in Diarrheic Pigs of Different Ages in Northern Italy

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Development of RV Multiplex RT-qPCR Assays
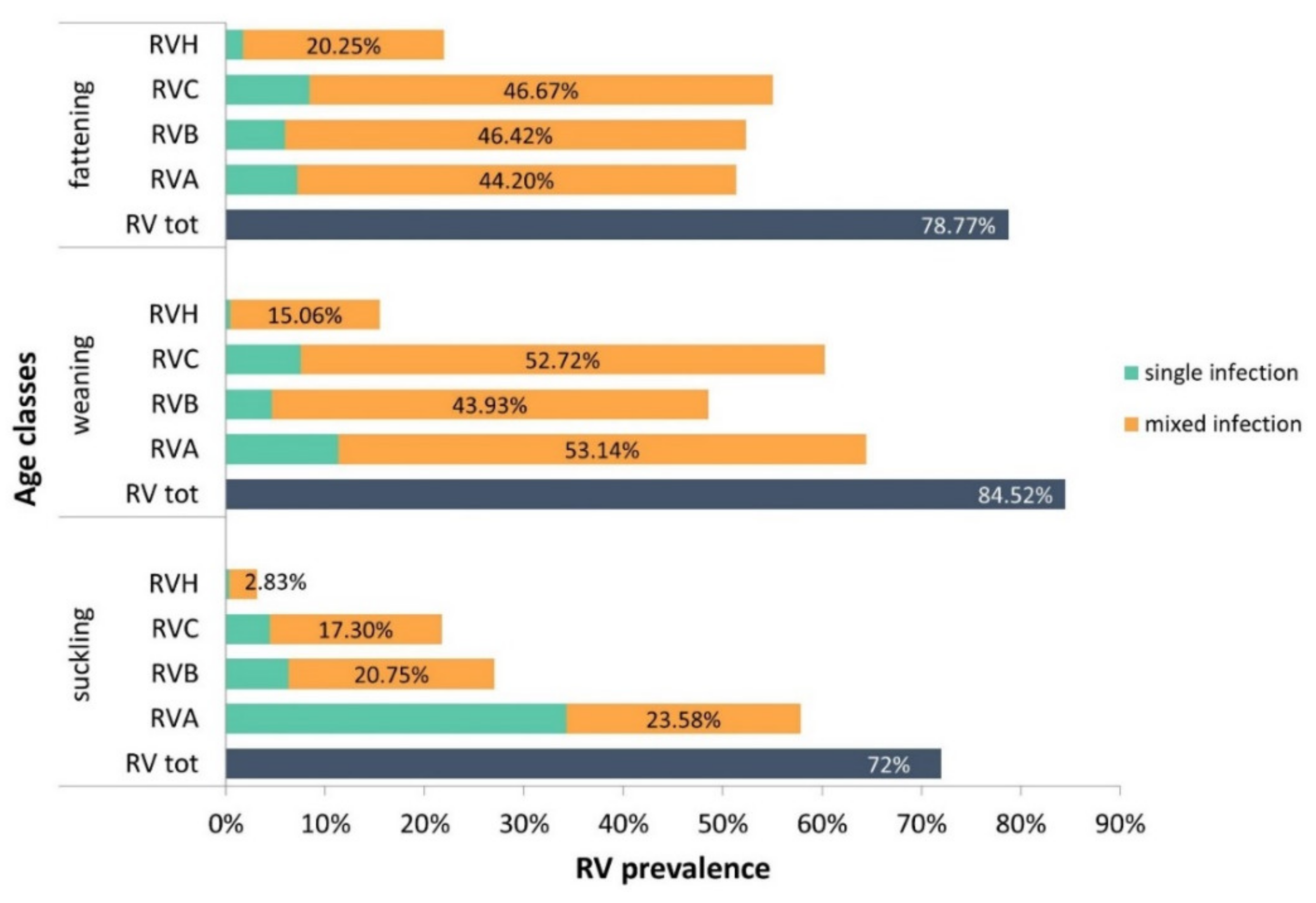
2.2. RV Group Distribution among the Age Classes
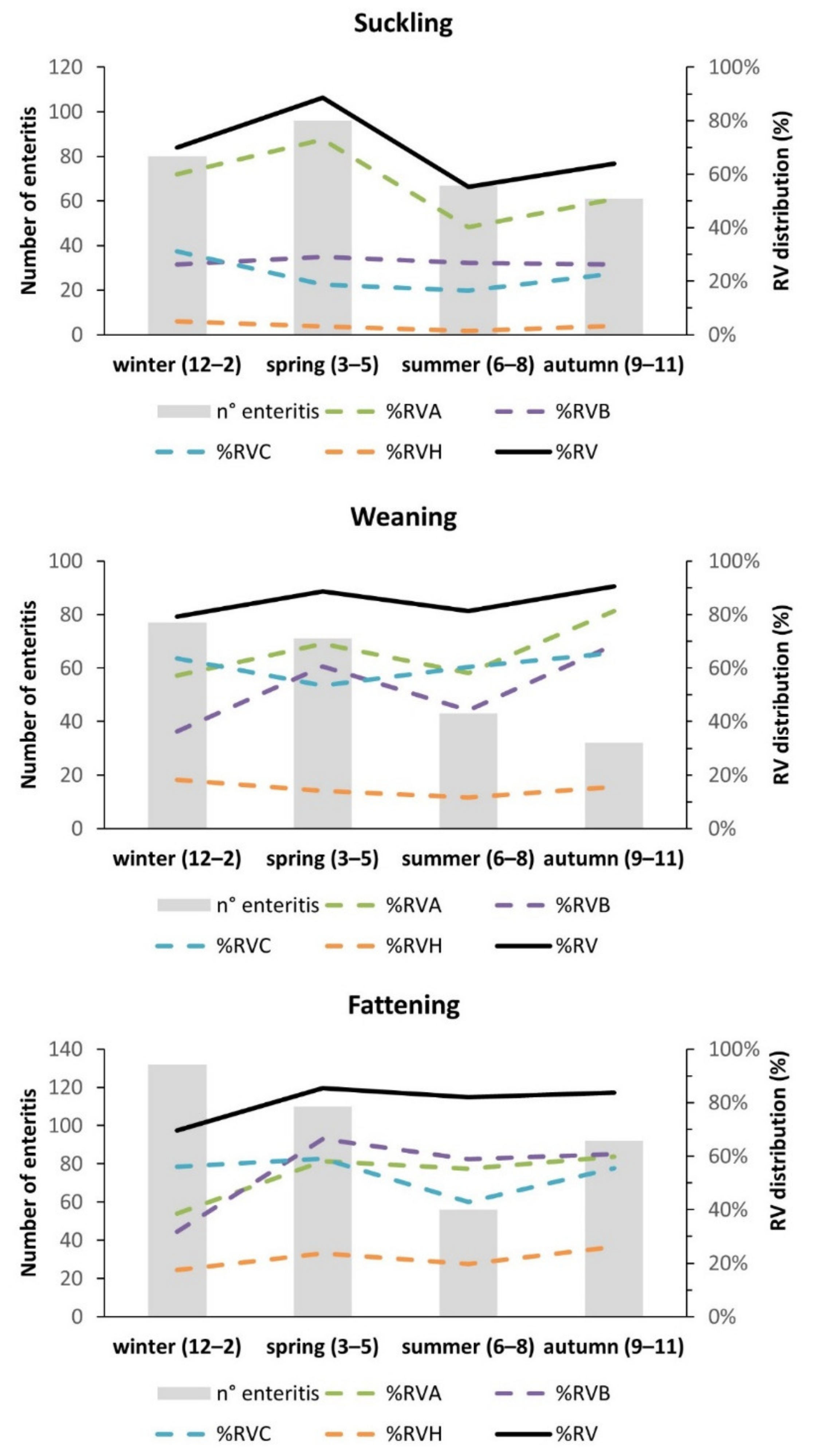
2.3. Seasonal Distribution of RV among Age Classes
2.4. Co-Infections of RV with Other Pathogens
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. Quantitative RT-qPCR for Detection of RVs
4.3. RV RT-qPCR Performances
4.4. Bacterial and Viral Agent Detection
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matthijnssens, J.; Otto, P.H.; Ciarlet, M.; Desselberger, U.; Van Ranst, M.; Johne, R. VP6-Sequence-Based Cutoff Values as a Criterion for Rotavirus Species Demarcation. Arch. Virol. 2012, 157, 1177–1182. [Google Scholar] [CrossRef]
- Bányai, K.; Kemenesi, G.; Budinski, I.; Földes, F.; Zana, B.; Marton, S.; Varga-Kugler, R.; Oldal, M.; Kurucz, K.; Jakab, F. Candidate New Rotavirus Species in Schreiber’s Bats, Serbia. Infect. Genet. Evol. 2017, 48, 19–26. [Google Scholar] [CrossRef]
- Kindler, E.; Trojnar, E.; Heckel, G.; Otto, P.H.; Johne, R. Analysis of Rotavirus Species Diversity and Evolution Including the Newly Determined Full-Length Genome Sequences of Rotavirus F and G. Infect. Genet. Evol. 2013, 14, 58–67. [Google Scholar] [CrossRef]
- Bányai, K.; Estes, M.K.; Martella, V.; Parashar, U.D. Viral Gastroenteritis. Lancet Lond. Engl. 2018, 392, 175–186. [Google Scholar] [CrossRef]
- Dóró, R.; Farkas, S.L.; Martella, V.; Bányai, K. Zoonotic Transmission of Rotavirus: Surveillance and Control. Expert Rev. Anti Infect. Ther. 2015, 13, 1337–1350. [Google Scholar] [CrossRef]
- Martella, V.; Bányai, K.; Matthijnssens, J.; Buonavoglia, C.; Ciarlet, M. Zoonotic Aspects of Rotaviruses. Vet. Microbiol. 2010, 140, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Cowley, D.; Donato, C.M.; Roczo-Farkas, S.; Kirkwood, C.D. Emergence of a Novel Equine-like G3P[8] Inter-Genogroup Reassortant Rotavirus Strain Associated with Gastroenteritis in Australian Children. J. Gen. Virol. 2016, 97, 403–410. [Google Scholar] [CrossRef]
- Katsuda, K.; Kohmoto, M.; Kawashima, K.; Tsunemitsu, H. Frequency of Enteropathogen Detection in Suckling and Weaned Pigs with Diarrhea in Japan. J. Vet. Diagn. Investig. 2006, 18, 350–354. [Google Scholar] [CrossRef]
- Marthaler, D.; Homwong, N.; Rossow, K.; Culhane, M.; Goyal, S.; Collins, J.; Matthijnssens, J.; Ciarlet, M. Rapid Detection and High Occurrence of Porcine Rotavirus A, B, and C by RT-QPCR in Diagnostic Samples. J. Virol. Methods 2014, 209, 30–34. [Google Scholar] [CrossRef]
- Wakuda, M. Porcine Rotavirus Closely Related to Novel Group of Human Rotaviruses. Emerg. Infect. Dis. 2011. [Google Scholar] [CrossRef]
- Smitalova, R.; Rodak, L.; Smid, B.; Psikal, I. Detection of Nongroup a Rotaviruses in Faecal Samples of Pigs in the Czech Republic. Veterinární Medicína 2009, 54, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Saif, L.J.; Jiang, B. Nongroup A Rotaviruses of Humans and Animals. In Rotaviruses; Ramig, R.F., Ed.; Current Topics in Microbiology and Immunology; Springer Berlin Heidelberg: Berlin, Heidelberg, 1994; Volume 185, pp. 339–371. ISBN 978-3-642-78258-9. [Google Scholar]
- Otto, P.H.; Rosenhain, S.; Elschner, M.C.; Hotzel, H.; Machnowska, P.; Trojnar, E.; Hoffmann, K.; Johne, R. Detection of Rotavirus Species A, B and C in Domestic Mammalian Animals with Diarrhoea and Genotyping of Bovine Species A Rotavirus Strains. Vet. Microbiol. 2015, 179, 168–176. [Google Scholar] [CrossRef]
- Chasey, D.; Bridger, J.C.; McCrae, M.A. A New Type of Atypical Rotavirus in Pigs. Arch. Virol. 1986, 89, 235–243. [Google Scholar] [CrossRef]
- Amimo, J.O.; Vlasova, A.N.; Saif, L.J. Detection and Genetic Diversity of Porcine Group A Rotaviruses in Historic (2004) and Recent (2011 and 2012) Swine Fecal Samples in Ohio: Predominance of the G9P[13] Genotype in Nursing Piglets. J. Clin. Microbiol. 2013, 51, 1142–1151. [Google Scholar] [CrossRef] [Green Version]
- Amimo, J.O.; Junga, J.O.; Ogara, W.O.; Vlasova, A.N.; Njahira, M.N.; Maina, S.; Okoth, E.A.; Bishop, R.P.; Saif, L.J.; Djikeng, A. Detection and Genetic Characterization of Porcine Group A Rotaviruses in Asymptomatic Pigs in Smallholder Farms in East Africa: Predominance of P[8] Genotype Resembling Human Strains. Vet. Microbiol. 2015, 175, 195–210. [Google Scholar] [CrossRef]
- Monini, M.; Zaccaria, G.; Ianiro, G.; Lavazza, A.; Vaccari, G.; Ruggeri, F.M. Full-Length Genomic Analysis of Porcine Rotavirus Strains Isolated from Pigs with Diarrhea in Northern Italy. Infect. Genet. Evol. 2014, 25, 4–13. [Google Scholar] [CrossRef]
- Marthaler, D.; Rossow, K.; Culhane, M.; Collins, J.; Goyal, S.; Ciarlet, M.; Matthijnssens, J. Identification, Phylogenetic Analysis and Classification of Porcine Group C Rotavirus VP7 Sequences from the United States and Canada. Virology 2013, 446, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Amimo, J.O.; Vlasova, A.N.; Saif, L.J. Prevalence and Genetic Heterogeneity of Porcine Group C Rotaviruses in Nursing and Weaned Piglets in Ohio, USA and Identification of a Potential New VP4 Genotype. Vet. Microbiol. 2013, 164, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Theuns, S.; Vyt, P.; Desmarets, L.M.B.; Roukaerts, I.D.M.; Heylen, E.; Zeller, M.; Matthijnssens, J.; Nauwynck, H.J. Presence and Characterization of Pig Group A and C Rotaviruses in Feces of Belgian Diarrheic Suckling Piglets. Virus Res. 2016, 213, 172–183. [Google Scholar] [CrossRef]
- Tuanthap, S.; Phupolphan, C.; Luengyosluechakul, S.; Duang-in, A.; Theamboonlers, A.; Wattanaphansak, S.; Vongpunsawad, S.; Amonsin, A.; Poovorawan, Y. Porcine Rotavirus C in Pigs with Gastroenteritis on Thai Swine Farms, 2011–2016. PeerJ 2018, 6, e4724. [Google Scholar] [CrossRef] [Green Version]
- Martella, V.; Bányai, K.; Lorusso, E.; Bellacicco, A.L.; Decaro, N.; Camero, M.; Bozzo, G.; Moschidou, P.; Arista, S.; Pezzotti, G. Prevalence of Group C Rotaviruses in Weaning and Post-Weaning Pigs with Enteritis. Vet. Microbiol. 2007, 123, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Kuga, K.; Miyazaki, A.; Suzuki, T.; Takagi, M.; Hattori, N.; Katsuda, K.; Mase, M.; Sugiyama, M.; Tsunemitsu, H. Genetic Diversity and Classification of the Outer Capsid Glycoprotein VP7 of Porcine Group B Rotaviruses. Arch. Virol. 2009, 154, 1785–1795. [Google Scholar] [CrossRef]
- Molinari, B.L.D.; Possatti, F.; Lorenzetti, E.; Alfieri, A.F.; Alfieri, A.A. Unusual Outbreak of Post-Weaning Porcine Diarrhea Caused by Single and Mixed Infections of Rotavirus Groups A., B, C, and H. Vet. Microbiol. 2016, 193, 125–132. [Google Scholar] [CrossRef]
- Vidal, A.; Martín-Valls, G.E.; Tello, M.; Mateu, E.; Martín, M.; Darwich, L. Prevalence of Enteric Pathogens in Diarrheic and Non-Diarrheic Samples from Pig Farms with Neonatal Diarrhea in the North East of Spain. Vet. Microbiol. 2019, 237, 108419. [Google Scholar] [CrossRef]
- Shepherd, F.; Murtaugh, M.; Chen, F.; Culhane, M.; Marthaler, D. Longitudinal Surveillance of Porcine Rotavirus B Strains from the United States and Canada and In Silico Identification of Antigenically Important Sites. Pathogens 2017, 6, 64. [Google Scholar] [CrossRef] [Green Version]
- Marthaler, D.; Rossow, K.; Culhane, M.; Goyal, S.; Collins, J.; Matthijnssens, J.; Nelson, M.; Ciarlet, M. Widespread Rotavirus H in Commercially Raised Pigs, United States. Emerg. Infect. Dis. 2014, 20, 1203–1206. [Google Scholar] [CrossRef] [Green Version]
- Molinari, B.L.D.; Lorenzetti, E.; Otonel, R.A.A.; Alfieri, A.F.; Alfieri, A.A. Species H Rotavirus Detected in Piglets with Diarrhea, Brazil, 2012. Emerg. Infect. Dis. 2014, 20, 1019–1022. [Google Scholar] [CrossRef]
- Song, D.S.; Kang, B.K.; Oh, J.S.; Ha, G.W.; Yang, J.S.; Moon, H.J.; Jang, Y.-S.; Park, B.K. Multiplex Reverse Transcription-PCR for Rapid Differential Detection of Porcine Epidemic Diarrhea Virus, Transmissible Gastroenteritis Virus, and Porcine Group a Rotavirus. J. Vet. Diagn. Investig. 2006, 18, 278–281. [Google Scholar] [CrossRef] [Green Version]
- Moutelíková, R.; Prodělalová, J.; Dufková, L. Diversity of VP7, VP4, VP6, NSP2, NSP4, and NSP5 Genes of Porcine Rotavirus C: Phylogenetic Analysis and Description of Potential New VP7, VP4, VP6, and NSP4 Genotypes. Arch. Virol. 2015, 160, 1715–1727. [Google Scholar] [CrossRef]
- Chandler-Bostock, R.; Hancox, L.R.; Nawaz, S.; Watts, O.; Iturriza-Gomara, M.; Mellits, K.M. Genetic Diversity of Porcine Group A Rotavirus Strains in the UK. Vet. Microbiol. 2014, 173, 27–37. [Google Scholar] [CrossRef]
- Molinari, B.L.D.; Alfieri, A.F.; Alfieri, A.A. Genetic Variability of VP6, VP7, VP4, and NSP4 Genes of Porcine Rotavirus Group H Detected in Brazil. Virus Res. 2015, 197, 48–53. [Google Scholar] [CrossRef]
- Luppi, A.; Gibellini, M.; Gin, T.; Vangroenweghe, F.; Vandenbroucke, V.; Bauerfeind, R.; Bonilauri, P.; Labarque, G.; Hidalgo, Á. Prevalence of Virulence Factors in Enterotoxigenic Escherichia Coli Isolated from Pigs with Post-Weaning Diarrhoea in Europe. Porc. Health Manag. 2016, 2, 20. [Google Scholar] [CrossRef] [Green Version]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post Weaning Diarrhea in Pigs: Risk Factors and Non-Colistin-Based Control Strategies. Acta Vet. Scand. 2017, 59, 31. [Google Scholar] [CrossRef] [Green Version]
- Homwong, N.; Diaz, A.; Rossow, S.; Ciarlet, M.; Marthaler, D. Three-Level Mixed-Effects Logistic Regression Analysis Reveals Complex Epidemiology of Swine Rotaviruses in Diagnostic Samples from North America. PLoS ONE 2016, 11, e0154734. [Google Scholar] [CrossRef] [Green Version]
- Bertasio, C.; Giacomini, E.; Lazzaro, M.; Perulli, S.; Papetti, A.; Lavazza, A.; Lelli, D.; Alborali, G.; Boniotti, M.B. Porcine Epidemic Diarrhea Virus Shedding and Antibody Response in Swine Farms: A Longitudinal Study. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.F.; Ward, G.E.; Murtaugh, M.P.; Lin, G.; Gebhart, C.J. Enhanced Detection of Intracellular Organism of Swine Proliferative Enteritis, Ileal Symbiont Intracellularis, in Feces by Polymerase Chain Reaction. J. Clin. Microbiol. 1993, 31, 2611–2615. [Google Scholar] [CrossRef] [Green Version]
- Willems, H.; Reiner, G. A Multiplex Real-Time PCR for the Simultaneous Detection and Quantitation of Brachyspira Hyodysenteriae, Brachyspira Pilosicoli and Lawsonia Intracellularis in Pig Faeces. Berl. Munch. Tierarztl. Wochenschr. 2010, 123, 205–209. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 1 August 2020).

{kind=link}
{kind=link}
| RVA-RVB Assay | RVC-RVH Assay | |||
|---|---|---|---|---|
| Test | RVA | RVB | RVC | RVH |
| Slope § | −3.3896 ± 0.0495 | −3.428 ± 0.027 | −3.309 ± 0.019 | −3.263 ± 0.045 |
| Calibration curve (R2) § | 0.9996 ± 0.0001 | 0.999 ± 0.0002 | 0.999 ± 0.001 | 0.998 ± 0.001 |
| PCR efficiency (%) § | 97.093 ± 1.943 | 95.770 ± 1.035 | 100.6 ± 0.839 | 102.6 ± 1.950 |
| Specificity (%) | 100 | 100 | 100 | 100 |
| LOD (genomic copies) ‡ | 100 | 100 | 100 | 100 |
| RV Group | χ2 | p-Value | S vs. W | S vs. F | W vs. F |
|---|---|---|---|---|---|
| RV (all groups) | 12.68 | 0.002 * | 0.0005 * | 0.035 * | 0.070 |
| RVA total infections | 10.71 | 0.005 * | 0.259 | 0.190 | 0.001 * |
| RVA single infections | 99.85 | <0.0001 * | <0.0001 * | <0.0001 * | 0.070 |
| RVA mixed infections | 55.99 | <0.0001 * | <0.0001 * | <0.0001 * | 0.028 |
| RVB total infections | 50.44 | <0.0001 * | <0.0001 * | <0.0001 * | 0.619 |
| RVB single infections | 0.78 | 0.679 | 0.390 | 0.839 | 0.474 |
| RVB mixed infections | 56.11 | <0.0001 * | <0.0001 * | <0.0001 * | 0.540 |
| RVC total infections | 108.61 | <0.0001 * | <0.0001 * | <0.0001 * | 0.404 |
| RVC single infections | 4.67 | 0.097 | 0.136 | 0.323 | 0.461 |
| RVC mixed infections | 48.12 | <0.0001 * | <0.0001 * | <0.0001 * | 0.100 |
| RVH total infections | 52.51 | <0.0001 * | <0.0001 * | <0.0001 * | 0.113 |
| RVH single infections † | N.D | N.D | 1 | 0.085 | 0.269 |
| RVH mixed infections | 48.18 | <0.0001 * | <0.0001 * | <0.0001 * | 0.100 |
| Suckling (n = 110) | Weaning (n = 41) | Fattening (n = 163) | ||||
|---|---|---|---|---|---|---|
| n | % | n | % | n | % | |
| Single infections | 10 | 9.1% | 9 | 22.0% | 7 | 4.3% |
| RV | 6 | 5.5% | 6 | 14.6% | 2 | 1.2% |
| E.coli | 2 | 1.8% | 2 | 4.9% | 5 | 3.1% |
| PEDV | 1 | 0.9% | 1 | 2.4% | 0 | 0.0% |
| C.difficile | 1 | 0.9% | 0 | 0.0% | - | - |
| C.perfrigens | 0 | 0.0% | 0 | 0.0% | - | - |
| B.pilosicoli | - | - | - | - | 0 | 0.0% |
| B.hyodisenteriae | - | - | - | - | 0 | 0.0% |
| L.intracellularis | - | - | - | 0 | 0.0% | |
| RV mixed infections | 75 | 68.2% | 26 | 63.4% | 139 | 85.3% |
| RV+E.coli | 66 | 60.0% | 17 | 41.5% | 132 | 81.0% |
| RV+PEDV | 6 | 5.5% | 9 | 22.0% | 26 | 16.0% |
| RV+C.difficile | 28 | 25.5% | 4 | 9.8% | - | - |
| RV+C.perfrigens | 32 | 29.1% | 6 | 14.6% | - | - |
| RV+B.pilosicoli | - | - | - | - | 7 | 4.3% |
| RV+B.hyodisenteriae | - | - | - | - | 27 | 16.6% |
| RV+L. intracellularis | - | - | - | - | 32 | 19.6% |
| non RV co-infections | 23 | 20.9% | 5 | 12.2% | 17 | 10.4% |
| negatives | 2 | 1.8% | 1 | 2.4% | 0 | 0.0% |
| RT-qPCR Assays | 5′–3′ Sequence | Conc. (nM) |
|---|---|---|
| RVA-RVB assay | RVA_VP6_for CACCTTCAAGAGARGATAAYTTRCAA | 500 |
| RVA_VP6_rev TCGGATACCAGGTRKTTAGCCT | 500 | |
| RVA_VP6_probe FAM-TCCATTAGAAGCATGTTGAT-MGB | 200 | |
| RVB_VP6_for TRTGGKGWCARAARATAGCRAT | 500 | |
| RVB_VP6_rev ACCTYTCGAAGCACTYCCWTT | 500 | |
| RVB_VP6_probe VIC-TGATCCGGCGTCRGCT-MGB | 100 | |
| RVC-RVH assay | RVC_VP6_for TGTAGCATGATTCACGAATGGGT | 500 |
| RVC_VP6_rev ACATTTCATCCTCCTGGGGATC | 500 | |
| RVC_VP6_probe CY5-GCGTAGGGGCAAATGCGCATGA-BHQ2 | 100 | |
| RVH_VP6_for CCACCACAATTMGTTCAYTGGTC | 750 | |
| RVH_VP6_rev TCCCAGTGCGTGACCAGAT | 750 | |
| RVH_VP6_probe FAM-GCATGTTTAATTGCAGCYTATTC-MGB | 200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrari, E.; Salogni, C.; Martella, V.; Alborali, G.L.; Scaburri, A.; Boniotti, M.B. Assessing the Epidemiology of Rotavirus A, B, C and H in Diarrheic Pigs of Different Ages in Northern Italy. Pathogens 2022, 11, 467. https://doi.org/10.3390/pathogens11040467
Ferrari E, Salogni C, Martella V, Alborali GL, Scaburri A, Boniotti MB. Assessing the Epidemiology of Rotavirus A, B, C and H in Diarrheic Pigs of Different Ages in Northern Italy. Pathogens. 2022; 11(4):467. https://doi.org/10.3390/pathogens11040467
Chicago/Turabian StyleFerrari, Elena, Cristian Salogni, Vito Martella, Giovanni Loris Alborali, Alessandra Scaburri, and Maria Beatrice Boniotti. 2022. "Assessing the Epidemiology of Rotavirus A, B, C and H in Diarrheic Pigs of Different Ages in Northern Italy" Pathogens 11, no. 4: 467. https://doi.org/10.3390/pathogens11040467
APA StyleFerrari, E., Salogni, C., Martella, V., Alborali, G. L., Scaburri, A., & Boniotti, M. B. (2022). Assessing the Epidemiology of Rotavirus A, B, C and H in Diarrheic Pigs of Different Ages in Northern Italy. Pathogens, 11(4), 467. https://doi.org/10.3390/pathogens11040467