
Impact of Vaccination on Rotavirus Genotype Diversity: A Nearly Two-Decade-Long Epidemiological Study before and after Rotavirus Vaccine Introduction in Sicily, Italy



 ,
,  and
and 
Abstract
1. Introduction
2. Results
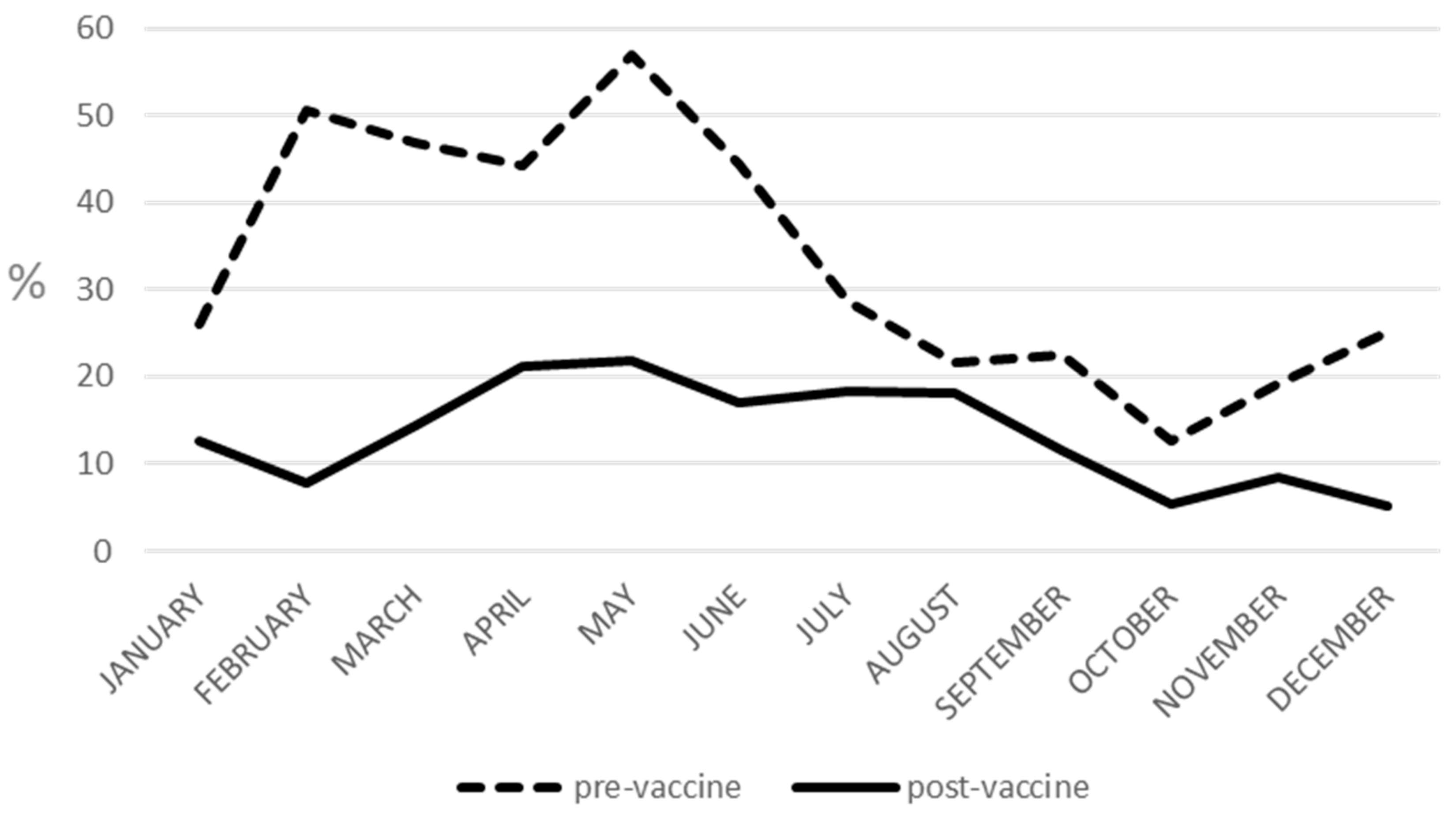
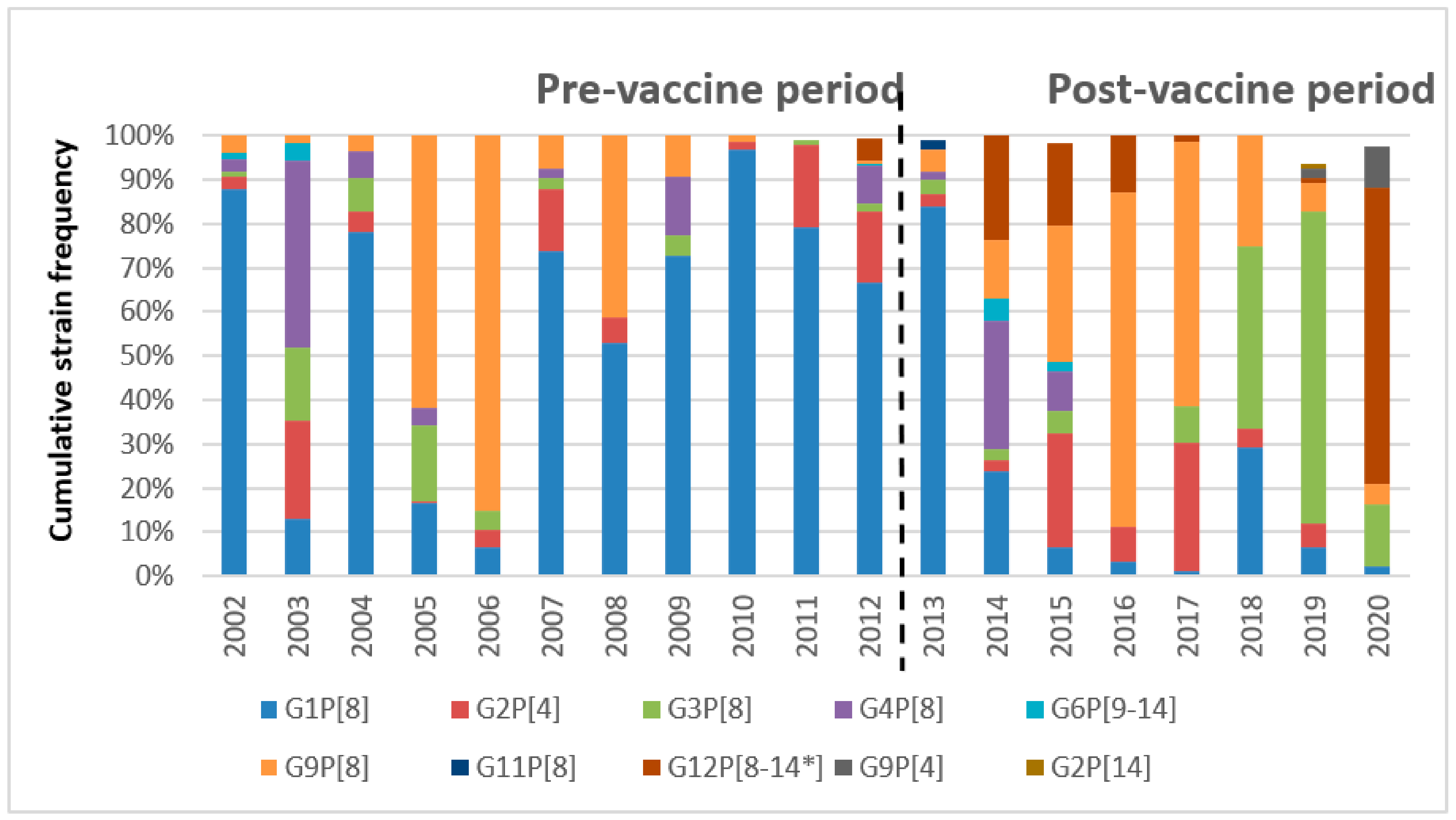
2.1. Seasonality and G/P Temporal Distribution
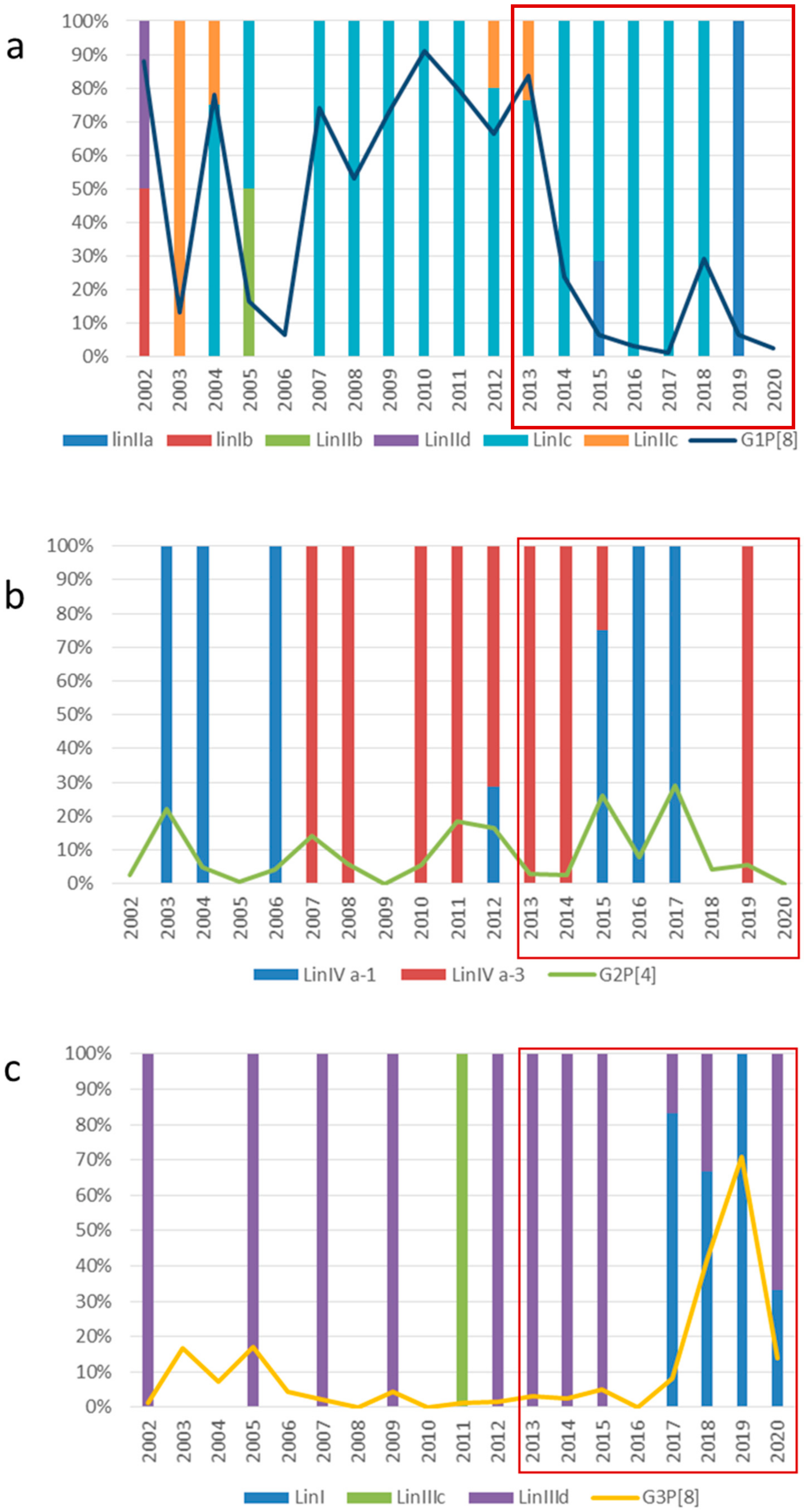
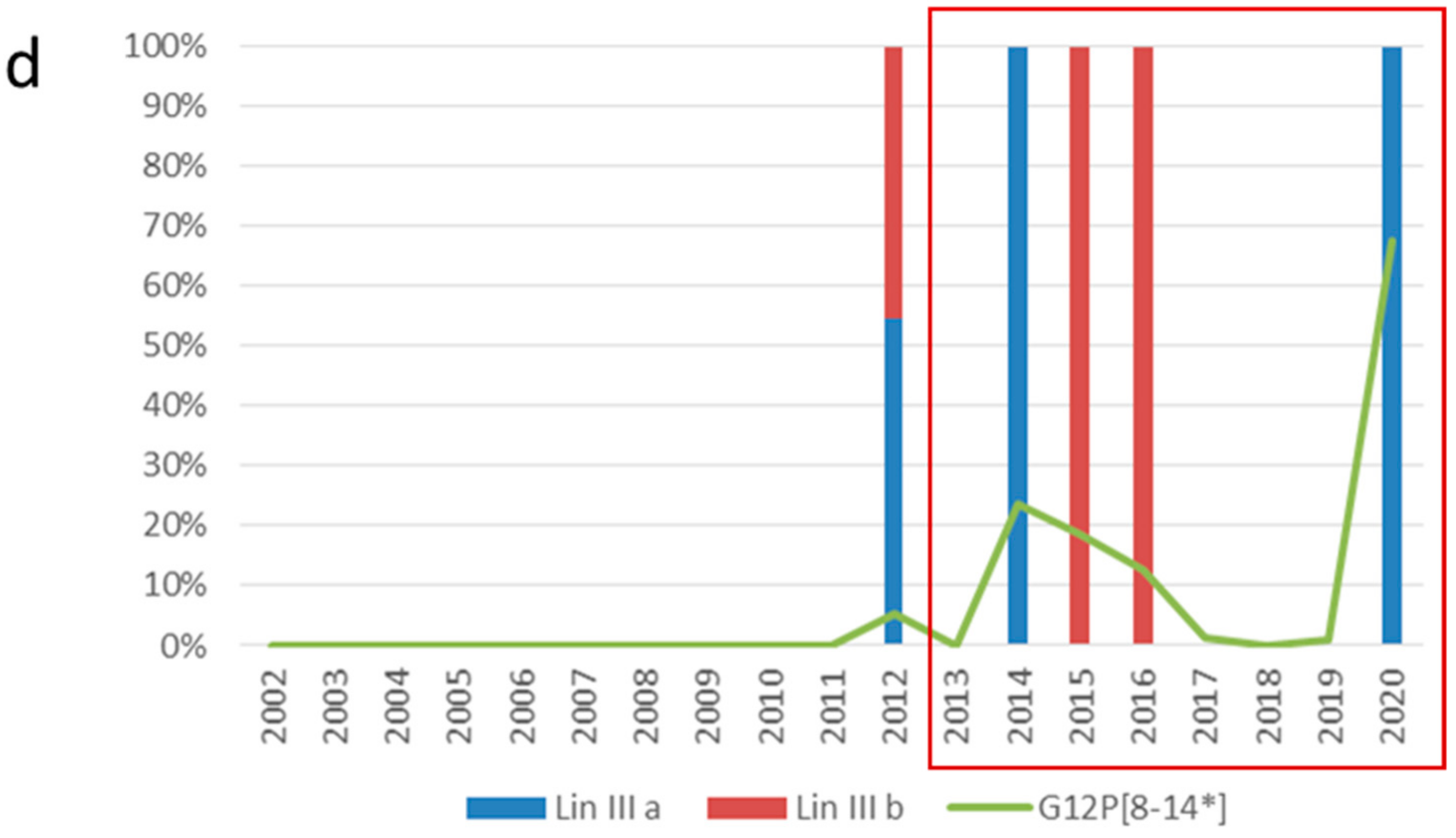
2.2. VP7 and VP4 Phylogenetic Analyses
2.3. Comparative Analysis of Neutralizing Epitopes of the VP7 and VP4 Proteins
3. Discussion
4. Material and Methods
4.1. Study Population
4.2. G and P Genotyping and Phylogenetic Analyses
4.3. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Parashar, U.D. Global, Regional, and National Estimates of Rotavirus Mortality in Children <5 Years of Age, 2000–2013. Clin. Infect. Dis. 2016, 62, S96–S105. [Google Scholar] [CrossRef] [PubMed]
- Mast, T.C.; DeMuro-Mercon, C.; Kelly, C.M.; Floyd, L.E.; Walter, E.B. The Impact of Rotavirus Gastroenteritis on the Family. BMC Pediatr. 2009, 9, 11. [Google Scholar] [CrossRef]
- Ciarlet, M.; Schodel, F. Development of a Rotavirus Vaccine: Clinical Safety, Immunogenicity, and Efficacy of the Pentavalent Rotavirus Vaccine, RotaTeq. Vaccine 2009, 27, G72–G81. [Google Scholar] [CrossRef] [PubMed]
- Zeller, M.; Rahman, M.; Heylen, E.; De Coster, S.; De Vos, S.; Arijs, I.; Novo, L.; Verstappen, N.; Van Ranst, M.; Matthijnssens, J. Rotavirus Incidence and Genotype Distribution before and after National Rotavirus Vaccine Introduction in Belgium. Vaccine 2010, 28, 7507–7513. [Google Scholar] [CrossRef] [PubMed]
- Vesikari, T.; Matson, D.O.; Dennehy, P.; Van Damme, P.; Santosham, M.; Rodriguez, Z.; Dallas, M.J.; Heyse, J.F.; Goveia, M.G.; Black, S.B.; et al. Safety and Efficacy of a Pentavalent Human-Bovine (WC3) Reassortant Rotavirus Vaccine. N. Engl. J. Med. 2006, 354, 23–33. [Google Scholar] [CrossRef]
- Ruiz-Palacios, G.M.; Pérez-Schael, I.; Velázquez, F.R.; Abate, H.; Breuer, T.; Clemens, S.C.; Cheuvart, B.; Espinoza, F.; Gillard, P.; Innis, B.L.; et al. Safety and Efficacy of an Attenuated Vaccine against Severe Rotavirus Gastroenteritis. N. Engl. J. Med. 2006, 354, 11–22. [Google Scholar] [CrossRef]
- Estes, M.K.; Greenber, H.B. Rotaviruses. In Fields Virology, 6th ed.; Lippincott, Williams and Wilkins: Philadelphia, PA, USA, 2013; Volume 2. [Google Scholar]
- Matthijnssens, J.; Ciarlet, M.; Heiman, E.; Arijs, I.; Delbeke, T.; McDonald, S.M.; Palombo, E.A.; Iturriza-Gomara, M.; Maes, P.; Patton, J.T.; et al. Full Genome-Based Classification of Rotaviruses Reveals a Common Origin between Human Wa-Like and Porcine Rotavirus Strains and Human DS-1-like and Bovine Rotavirus Strains. J. Virol. 2008, 82, 3204–3219. [Google Scholar] [CrossRef]
- Matthijnssens, J.; Ciarlet, M.; Rahman, M.; Attoui, H.; Banyai, K.; Estes, M.K.; Gentsch, J.R.; Iturriza-Gomara, M.; Kirkwood, C.D.; Martella, V.; et al. Recommendations for the Classification of Group A Rotaviruses Using All 11 Genomic RNA Segments. Arch. Virol. 2008, 153, 1621–1629. [Google Scholar] [CrossRef]
- Papp, H.; Laszlo, B.; Jakab, F.; Ganesh, B.; De Grazia, S.; Matthijnssens, J.; Ciarlet, M.; Martella, V.; Banyai, K. Review of Group A Rotavirus Strains Reported in Swine and Cattle. Vet. Microbiol. 2013, 165, 190–199. [Google Scholar] [CrossRef]
- Martella, V.; Bányai, K.; Matthijnssens, J.; Buonavoglia, C.; Ciarlet, M. Zoonotic Aspects of Rotaviruses. Vet. Microbiol. 2010, 140, 246–255. [Google Scholar] [CrossRef]
- Steele, A.D.; Victor, J.C.; Carey, M.E.; Tate, J.E.; Atherly, D.E.; Pecenka, C.; Diaz, Z.; Parashar, U.D.; Kirkwood, C.D. Experiences with Rotavirus Vaccines: Can We Improve Rotavirus Vaccine Impact in Developing Countries? Hum. Vaccines Immunother. 2019, 15, 1215–1227. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Van Ranst, M. Genotype Constellation and Evolution of Group A Rotaviruses Infecting Humans. Curr. Opin. Virol. 2012, 2, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Joelsson, D.B.; Warakomski, D.J.; Zhou, T.; Mathis, P.K.; van Maanen, M.H.; Ranheim, T.S.; Ciarlet, M. Molecular and Biological Characterization of the 5 Human-Bovine Rotavirus (WC3)-Based Reassortant Strains of the Pentavalent Rotavirus Vaccine, RotaTeq. Virology 2010, 403, 111–127. [Google Scholar] [CrossRef]
- Zeller, M.; Patton, J.T.; Heylen, E.; De Coster, S.; Ciarlet, M.; Van Ranst, M.; Matthijnssens, J. Genetic Analyses Reveal Differences in the VP7 and VP4 Antigenic Epitopes between Human Rotaviruses Circulating in Belgium and Rotaviruses in Rotarix and RotaTeq. J. Clin. Microbiol. 2012, 50, 966–976. [Google Scholar] [CrossRef]
- Roczo-Farkas, S.; Kirkwood, C.D.; Cowley, D.; Barnes, G.L.; Bishop, R.F.; Bogdanovic-Sakran, N.; Boniface, K.; Donato, C.M.; Bines, J.E. The Impact of Rotavirus Vaccines on Genotype Diversity: A Comprehensive Analysis of 2 Decades of Australian Surveillance Data. J. Infect. Dis. 2018, 218, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Chaudhary, S.; Bubber, P.; Ray, P. Epidemiology and Genetic Diversity of Group A Rotavirus in Acute Diarrhea Patients in Pre-Vaccination Era in Himachal Pradesh, India. Vaccine 2019, 37, 5350–5356. [Google Scholar] [CrossRef]
- Tate, J.E.; Rheingans, R.D.; O’Reilly, C.E.; Obonyo, B.; Burton, D.C.; Tornheim, J.A.; Adazu, K.; Jaron, P.; Ochieng, B.; Kerin, T.; et al. Rotavirus Disease Burden and Impact and Cost-Effectiveness of a Rotavirus Vaccination Program in Kenya. J. Infect. Dis. 2009, 200, S76–S84. [Google Scholar] [CrossRef]
- Desai, R.; Parashar, U.D.; Lopman, B.; de Oliveira, L.H.; Clark, A.D.; Sanderson, C.F.B.; Tate, J.E.; Matus, C.R.; Andrus, J.K.; Patel, M.M. Potential Intussusception Risk versus Health Benefits from Rotavirus Vaccination in Latin America. Clin. Infect. Dis. 2012, 54, 1397–1405. [Google Scholar] [CrossRef]
- Hanquet, G.; Ducoffre, G.; Vergison, A.; Neels, P.; Sabbe, M.; Van Damme, P.; Van Herck, K. Impact of Rotavirus Vaccination on Laboratory Confirmed Cases in Belgium. Vaccine 2011, 29, 4698–4703. [Google Scholar] [CrossRef]
- Braeckman, T.; Van Herck, K.; Raes, M.; Vergison, A.; Sabbe, M.; Van Damme, P. Rotavirus Vaccines in Belgium: Policy and Impact. Pediatr. Infect. Dis. J. 2011, 30, S21–S24. [Google Scholar] [CrossRef]
- Luchs, A.; Cilli, A.; Morillo, S.G.; Carmona, R.D.C.C.; Timenetsky, M.D.C.S.T. Rotavirus Genotypes Circulating In Brazil, 2007–2012: Implications For The Vaccine Program. Rev. Inst. Med. Trop. Sao Paulo 2015, 57, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Hungerford, D.; Smith, K.; Tucker, A.; Iturriza-Gómara, M.; Vivancos, R.; McLeonard, C.; Cunliffe, N.A.; French, N. Population Effectiveness of the Pentavalent and Monovalent Rotavirus Vaccines: A Systematic Review and Meta-Analysis of Observational Studies. BMC Infect. Dis. 2017, 17, 569. [Google Scholar] [CrossRef] [PubMed]
- Bibera, G.L.; Chen, J.; Pereira, P.; Benninghoff, B. Dynamics of G2P[4] Strain Evolution and Rotavirus Vaccination: A Review of Evidence for Rotarix. Vaccine 2020, 38, 5591–5600. [Google Scholar] [CrossRef]
- Varghese, T.; Alokit Khakha, S.; Giri, S.; Nair, N.P.; Badur, M.; Gathwala, G.; Chaudhury, S.; Kaushik, S.; Dash, M.; Mohakud, N.K.; et al. Rotavirus Strain Distribution before and after Introducing Rotavirus Vaccine in India. Pathogens 2021, 10, 416. [Google Scholar] [CrossRef] [PubMed]
- Girish Kumar, C.P.; Giri, S.; Chawla-Sarkar, M.; Gopalkrishna, V.; Chitambar, S.D.; Ray, P.; Venkatasubramanian, S.; Borkakoty, B.; Roy, S.; Bhat, J.; et al. Epidemiology of Rotavirus Diarrhea among Children Less than 5 Years Hospitalized with Acute Gastroenteritis Prior to Rotavirus Vaccine Introduction in India. Vaccine 2020, 38, 8154–8160. [Google Scholar] [CrossRef]
- Thanh, H.D.; Tran, V.T.; Lim, I.; Kim, W. Emergence of Human G2P[4] Rotaviruses in the Post-Vaccination Era in South Korea: Footprints of Multiple Interspecies Re-Assortment Events. Sci. Rep. 2018, 8, 6011. [Google Scholar] [CrossRef]
- Gikonyo, J.N.; Mbatia, B.; Okanya, P.W.; Obiero, G.F.O.; Sang, C.; Steele, D.; Nyangao, J. Post-Vaccine Rotavirus Genotype Distribution in Nairobi County, Kenya. Int. J. Infect. Dis. 2020, 100, 434–440. [Google Scholar] [CrossRef]
- De Grazia, S.; Bonura, F.; Colomba, C.; Cascio, A.; Di Bernardo, F.; Collura, A.; Terranova, D.M.; Martella, V.; Giammanco, G.M. Data Mining from a 27-Years Rotavirus Surveillance in Palermo, Italy. Infect. Genet. Evol. 2014, 28, 377–384. [Google Scholar] [CrossRef]
- Dan Hungerford, Miren Iturriza-Gómara Eurorotanet Annual Report. 2020. Available online: https://Www.Eurorotanet.Com/Wp-Content/Uploads/2021/11/EuroRotaNet_report-2020_20211115_Final_v1.0.Pdf2021 (accessed on 19 March 2022).
- Arista, S.; Vizzi, E.; Migliore, M.C.; Di Rosa, E.; Cascio, A. High Incidence of G9P[8] Rotavirus Infections in Italian Children during the Winter Season 1999-2000. Eur. J. Epidemiol. 2003, 18, 711–714. [Google Scholar] [CrossRef]
- Giammanco, G.M.; Bonura, F.; DI Bernardo, F.; Cascio, A.; Ferrera, G.; Dones, P.; Saporito, L.; Collura, A.; Terranova, D.M.; Valenzise, M.; et al. Introduction and Prolonged Circulation of G12 Rotaviruses in Sicily. Epidemiol. Infect. 2016, 144, 1943–1950. [Google Scholar] [CrossRef][Green Version]
- Bonura, F.; Bányai, K.; Mangiaracina, L.; Bonura, C.; Martella, V.; Giammanco, G.M.; De Grazia, S. Emergence in 2017–2019 of Novel Reassortant Equine-like G3 Rotavirus Strains in Palermo, Sicily. Transbound. Emerg. Dis. 2021, 69, 813–835. [Google Scholar] [CrossRef] [PubMed]
- Carvalho-Costa, F.A.; de Assis, R.M.S.; Fialho, A.M.; Araújo, I.T.; Silva, M.F.; Gómez, M.M.; Andrade, J.S.; Rose, T.L.; Fumian, T.M.; Volotão, E.M.; et al. The Evolving Epidemiology of Rotavirus A Infection in Brazil a Decade after the Introduction of Universal Vaccination with Rotarix®. BMC Pediatr. 2019, 19, 42. [Google Scholar] [CrossRef] [PubMed]
- Hungerford, D.; Allen, D.J.; Nawaz, S.; Collins, S.; Ladhani, S.; Vivancos, R.; Iturriza-Gómara, M. Impact of Rotavirus Vaccination on Rotavirus Genotype Distribution and Diversity in England, September 2006 to August 2016. Euro Surveill. 2019, 24, 1700774. [Google Scholar] [CrossRef] [PubMed]
- Arista, S.; Giammanco, G.M.; De Grazia, S.; Ramirez, S.; Lo Biundo, C.; Colomba, C.; Cascio, A.; Martella, V. Heterogeneity and Temporal Dynamics of Evolution of G1 Human Rotaviruses in a Settled Population. J. Virol. 2006, 80, 10724–10733. [Google Scholar] [CrossRef] [PubMed]
- Donato, C.M.; Cowley, D.; Snelling, T.L.; Akopov, A.; Kirkness, E.F.; Kirkwood, C.D. Characterization of a G1P[8] Rotavirus Causing an Outbreak of Gastroenteritis in the Northern Territory, Australia, in the Vaccine Era. Emerg. Microbes Infect. 2014, 3, e47. [Google Scholar] [CrossRef] [PubMed]
- Harastani, H.H.; Reslan, L.; Sabra, A.; Ali, Z.; Hammadi, M.; Ghanem, S.; Hajar, F.; Matar, G.M.; Dbaibo, G.S.; Zaraket, H. Genetic Diversity of Human Rotavirus A Among Hospitalized Children Under-5 Years in Lebanon. Front. Immunol. 2020, 11, 317. [Google Scholar] [CrossRef]
- Wangchuk, S.; Mitui, M.T.; Tshering, K.; Yahiro, T.; Bandhari, P.; Zangmo, S.; Dorji, T.; Tshering, K.; Matsumoto, T.; Nishizono, A.; et al. Dominance of Emerging G9 and G12 Genotypes and Polymorphism of VP7 and VP4 of Rotaviruses from Bhutanese Children with Severe Diarrhea Prior to the Introduction of Vaccine. PLoS ONE 2014, 9, e110795. [Google Scholar] [CrossRef]
- Iturriza-Gomara, M.; Green, J.; Brown, D.W.; Desselberger, U.; Gray, J.J. Comparison of Specific and Random Priming in the Reverse Transcriptase Polymerase Chain Reaction for Genotyping Group A Rotaviruses. J Virol. Methods 1999, 78, 93–103. [Google Scholar] [CrossRef]
- Pang, X.; Lee, B.; Chui, L.; Preiksaitis, J.K.; Monroe, S.S. Evaluation and Validation of Real-Time Reverse Transcription-Pcr Assay Using the LightCycler System for Detection and Quantitation of Norovirus. J. Clin. Microbiol. 2004, 42, 4679–4685. [Google Scholar] [CrossRef]
- Gentsch, J.R.; Glass, R.I.; Woods, P.; Gouvea, V.; Gorziglia, M.; Flores, J.; Das, B.K.; Bhan, M.K. Identification of Group A Rotavirus Gene 4 Types by Polymerase Chain Reaction. J. Clin. Microbiol. 1992, 30, 1365–1373. [Google Scholar] [CrossRef]
- Gouvea, V.; Glass, R.I.; Woods, P.; Taniguchi, K.; Clark, H.F.; Forrester, B.; Fang, Z.Y. Polymerase Chain Reaction Amplification and Typing of Rotavirus Nucleic Acid from Stool Specimens. J. Clin. Microbiol. 1990, 28, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Iturriza-Gomara, M.; Kang, G.; Gray, J. Rotavirus Genotyping: Keeping up with an Evolving Population of Human Rotaviruses. J. Clin. Virol. 2004, 31, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year of Surveillance | No. of Samples Tested | No. of Rotavirus-Positive Samples | Rotavirus Prevalence (%) | |
|---|---|---|---|---|
| Pre-vaccine period | 2002 | 196 | 76 | 38.78 |
| 2003 | 214 | 55 | 25.70 | |
| 2004 | 196 | 82 | 41.84 | |
| 2005 | 273 | 148 | 54.21 | |
| 2006 | 179 | 47 | 26.26 | |
| 2007 | 217 | 93 | 42.86 | |
| 2008 | 154 | 68 | 44.16 | |
| 2009 | 342 | 67 | 19.59 | |
| 2010 | 386 | 60 | 15.54 | |
| 2011 | 377 | 126 | 33.42 | |
| 2012 | 679 | 237 | 34.90 | |
| Overall Pre-vaccine | 3.213 | 1.059 | 36.18 | |
| Post-vaccine period | 2013 | 759 | 119 | 15.68 |
| 2014 | 627 | 47 | 7.50 | |
| 2015 | 504 | 134 | 26.59 | |
| 2016 | 492 | 64 | 13.01 | |
| 2017 | 707 | 87 | 12.31 | |
| 2018 | 468 | 29 | 6.20 | |
| 2019 | 683 | 93 | 10.94 | |
| 2020 | 393 | 43 | 25.44 | |
| Overall Post-vaccine | 4.633 | 616 | 13.36 |
| Months | Pre-Vaccine Prevalence (%) | Post-Vaccine Prevalence (%) |
|---|---|---|
| January | 26.07 | 12.59 |
| February | 50.55 | 7.75 |
| March | 46.99 | 14.48 |
| April | 44.24 | 21.21 |
| May | 56.96 | 21.83 |
| June | 44.51 | 16.98 |
| July | 28.57 | 18.25 |
| August | 21.71 | 18.08 |
| September | 22.41 | 11.47 |
| October | 12.60 | 5.43 |
| November | 19.18 | 8.51 |
| December | 25.22 | 5.09 |
| (a) G1P[8] | Epitopes | |||||
| 7-1a | 7-1b | 7-2 | ||||
| Strain/sub-lineage (No. of sequences analyzed) | Positions | |||||
| 94 | 212 | 97 | 123 | 147 | 217 | |
| Rotarix® (A41CB052A /1988/G1P1A[8]) | N | V | E | S | N | M |
| RotaTeq® (WI79-9/1992/G1P7[5]) | N | V | D | S | S | M |
| G1P[8] sub-lineage Ic (54) | S | T | E | N | N | T |
| G1P[8] sub-lineage IIa (6) | N | V | E | S/N | N | M |
| (b) G2P[4] | Epitopes | |||||
| 7-1a | 7-1b | |||||
| Strain/sub-lineage (No. of sequences analyzed) | Positions | |||||
| 87 | 96 | 213 | ||||
| RotaTeq® (SC2-9/1992/G2P7[5]) | A | D | S | |||
| G2P[4] sub-lineage IVa-1 (18) | T | N/S | D | |||
| G2P[4] sub-lineage IVa-3 (18) | T | N | D | |||
| (c) G3P[8] | Epitopes | |||||
| 7-1a | 7-1b | |||||
| Strain/sub-lineage (No. of sequences analyzed) | Positions | |||||
| 87 | 212 | 213 | 238 | 242 | ||
| RotaTeq® (WI78-8/1992/G3P75) | T | A | N | K | D | |
| G3P[8] human (24) | T/I | T | N | N | N | |
| G3P[8] equine-like (39) | S | T/A | T | D | A | |
| (d) G4P[8] | Epitopes | |||||
| 7-1a | 7-1b | 7-2 | ||||
| Strain/sub-lineage (No. of sequences analyzed) | Positions | |||||
| 130 | 211 | 143 | 145 | |||
| RotaTeq® (BrB-9/1996/G4Ia) | D | D | R | A | ||
| G4P[8] sub-lineage Ic (14) | E | N | K | T | ||
| Epitopes | ||||||||
|---|---|---|---|---|---|---|---|---|
| 8-1 | 8-3 | |||||||
| Strain/Sub-Lineage (No. of Sequences Analyzed) | Positions | |||||||
| 150 | 194 | 195 | 113 | 125 | 131 | 135 | ||
| Vaccine | Rotarix® (A1CB052A/1988/G1P[8]) | F | N | N | N | S | S | N |
| RotaTeq® (WI79-4/1992/G6P[8]) | F | N | D | N | N | R | D | |
| P[8]-I | G1P[8] (4) | F | N | N | N/K/D | S | S | N |
| P[8]-III | G1P[8] (50) | D | N/D | G/D | N/D | N | R | D |
| G3P[8] (19) | D | N/D | G | N/D | N | R | D | |
| G3P[8] Eq-like (30) | D | D | G | D | N/K | R | D | |
| G9P[8] (43) | D | D | G | N | N | R | D | |
| G12P[8] (21) | D | N/D | G | N/D | N | R | D | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonura, F.; Mangiaracina, L.; Filizzolo, C.; Bonura, C.; Martella, V.; Ciarlet, M.; Giammanco, G.M.; De Grazia, S. Impact of Vaccination on Rotavirus Genotype Diversity: A Nearly Two-Decade-Long Epidemiological Study before and after Rotavirus Vaccine Introduction in Sicily, Italy. Pathogens 2022, 11, 424. https://doi.org/10.3390/pathogens11040424
Bonura F, Mangiaracina L, Filizzolo C, Bonura C, Martella V, Ciarlet M, Giammanco GM, De Grazia S. Impact of Vaccination on Rotavirus Genotype Diversity: A Nearly Two-Decade-Long Epidemiological Study before and after Rotavirus Vaccine Introduction in Sicily, Italy. Pathogens. 2022; 11(4):424. https://doi.org/10.3390/pathogens11040424
Chicago/Turabian StyleBonura, Floriana, Leonardo Mangiaracina, Chiara Filizzolo, Celestino Bonura, Vito Martella, Max Ciarlet, Giovanni M. Giammanco, and Simona De Grazia. 2022. "Impact of Vaccination on Rotavirus Genotype Diversity: A Nearly Two-Decade-Long Epidemiological Study before and after Rotavirus Vaccine Introduction in Sicily, Italy" Pathogens 11, no. 4: 424. https://doi.org/10.3390/pathogens11040424
APA StyleBonura, F., Mangiaracina, L., Filizzolo, C., Bonura, C., Martella, V., Ciarlet, M., Giammanco, G. M., & De Grazia, S. (2022). Impact of Vaccination on Rotavirus Genotype Diversity: A Nearly Two-Decade-Long Epidemiological Study before and after Rotavirus Vaccine Introduction in Sicily, Italy. Pathogens, 11(4), 424. https://doi.org/10.3390/pathogens11040424