
G6P[8] Rotavirus a Possessing a Wa-like VP3 Gene from a Child with Acute Gastroenteritis Living in the Northwest Amazon Region


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. A G6P[8] RV Was Detected in a Fecal Sample Obtained from a Child Living in the Northwest Amazon Region
2.2. Sample G6P[8] ID-LVCA: 28,398 Was Initially Genotyped as G9 with Multiplex PCR
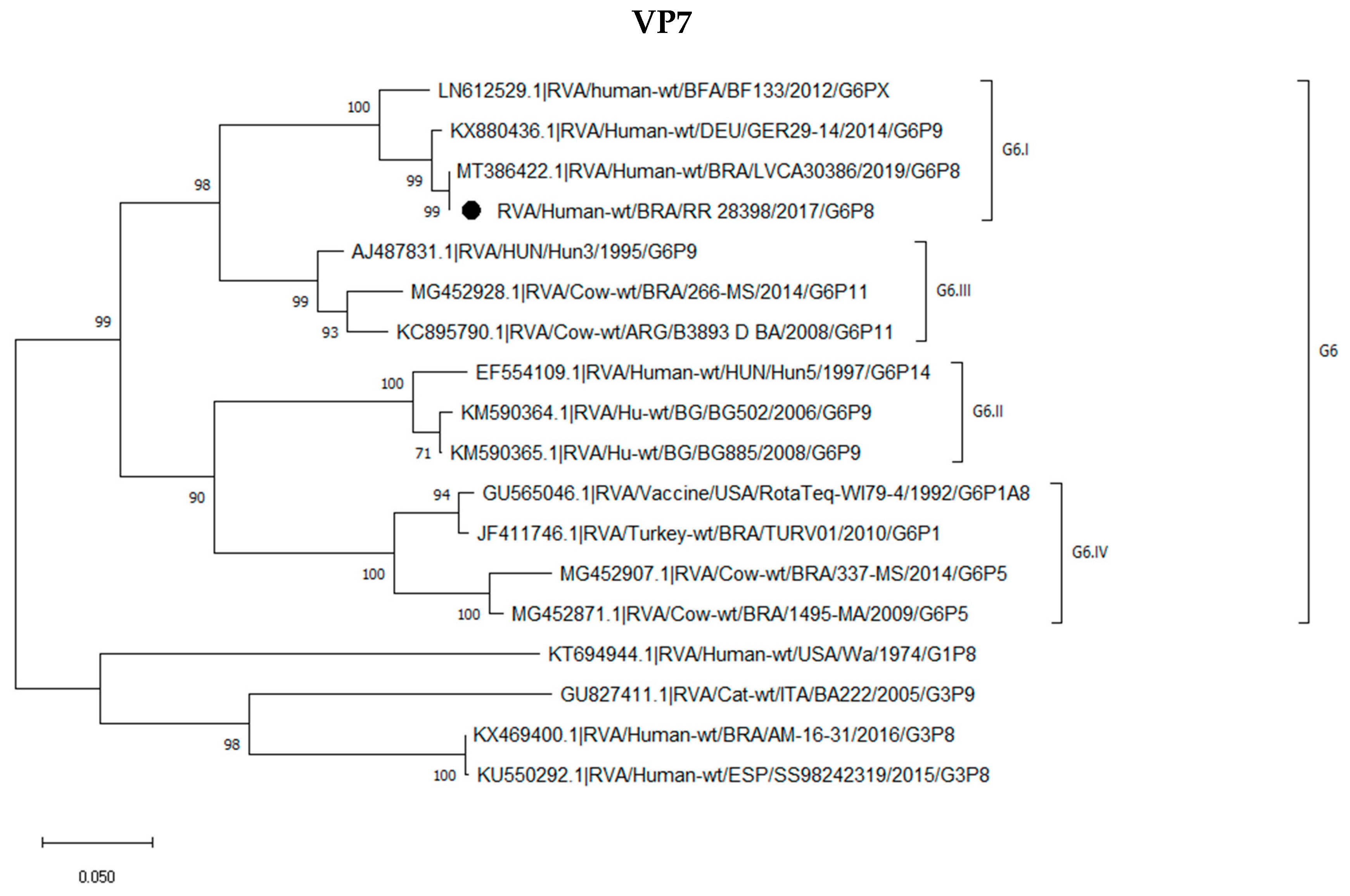
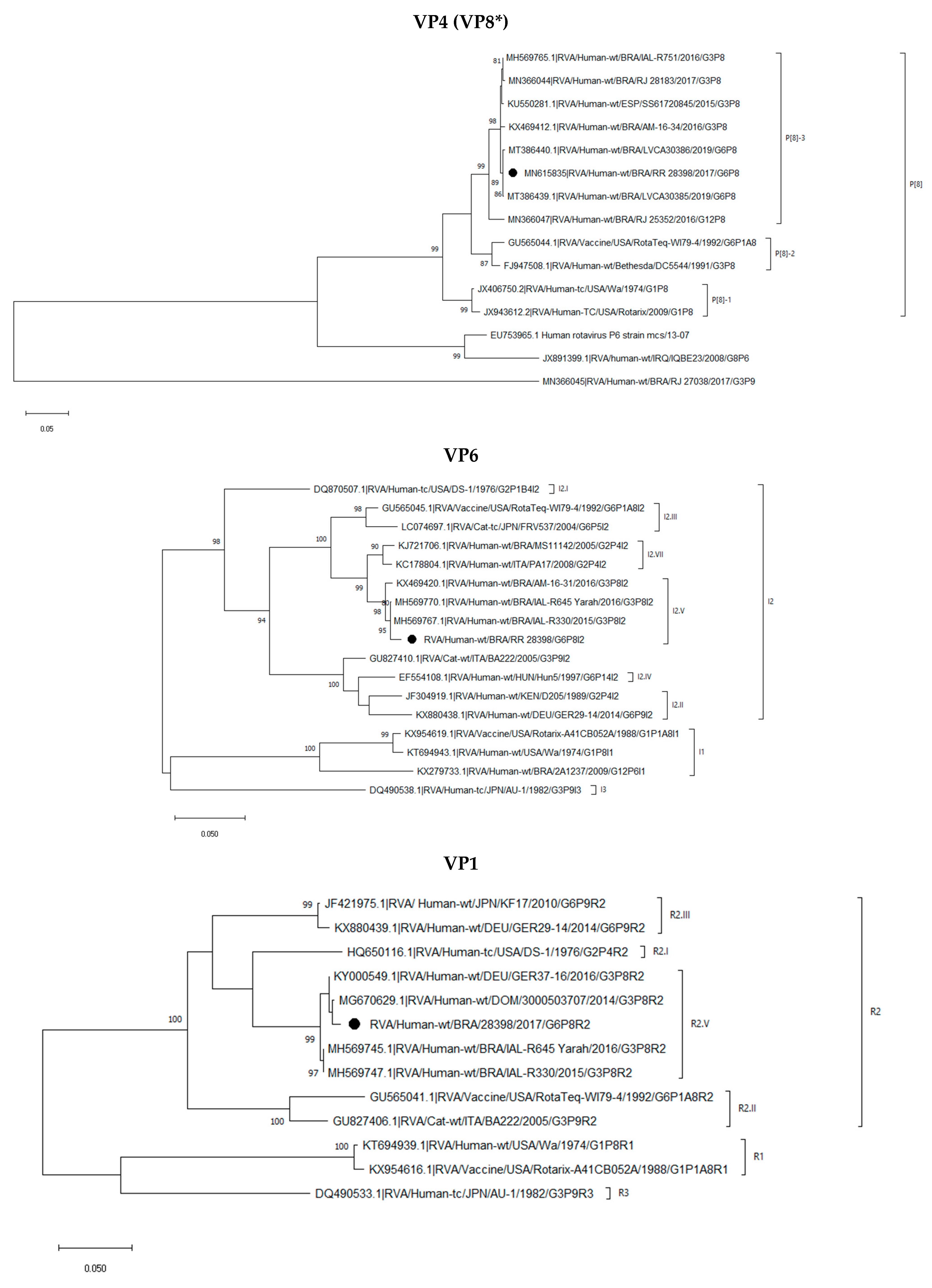
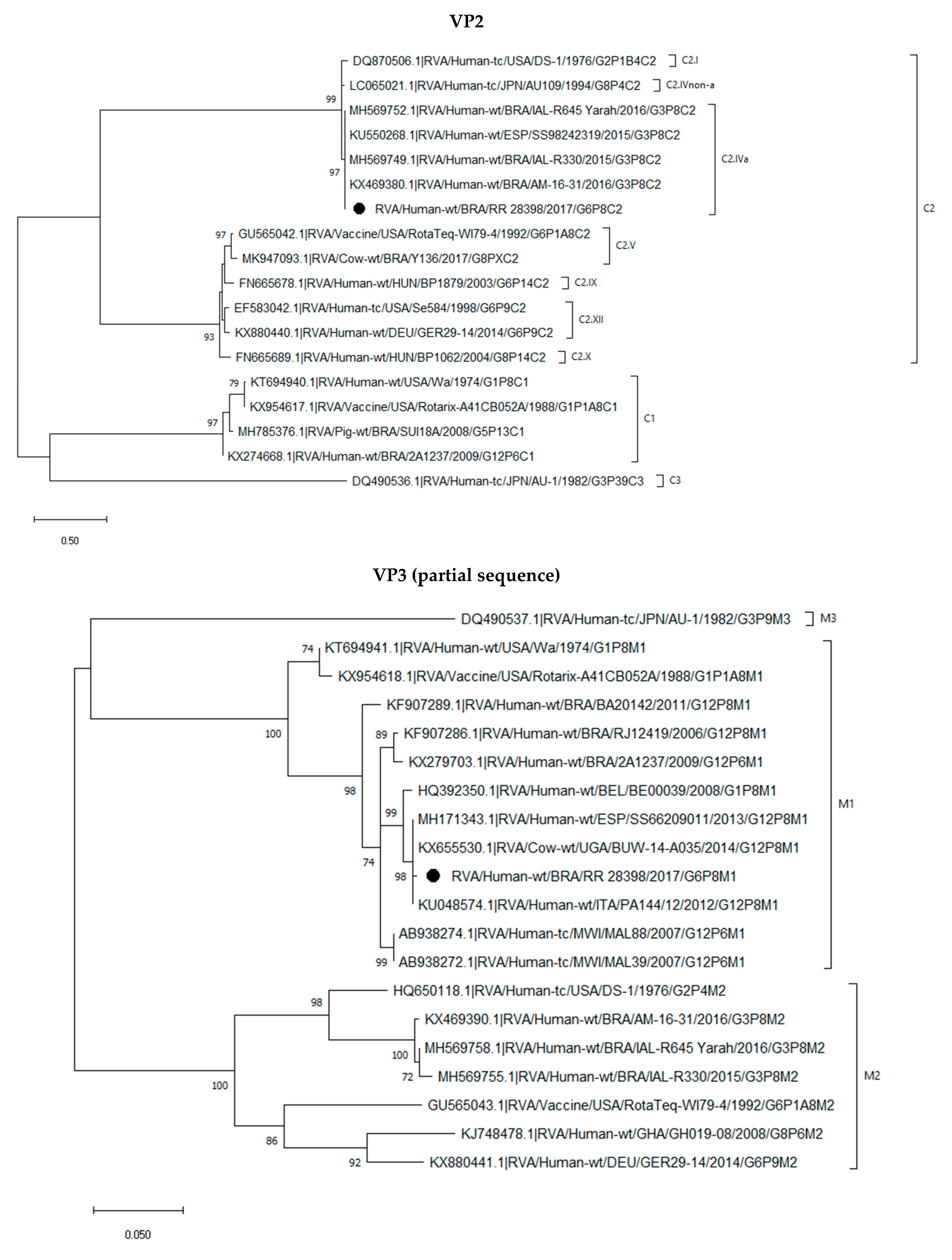
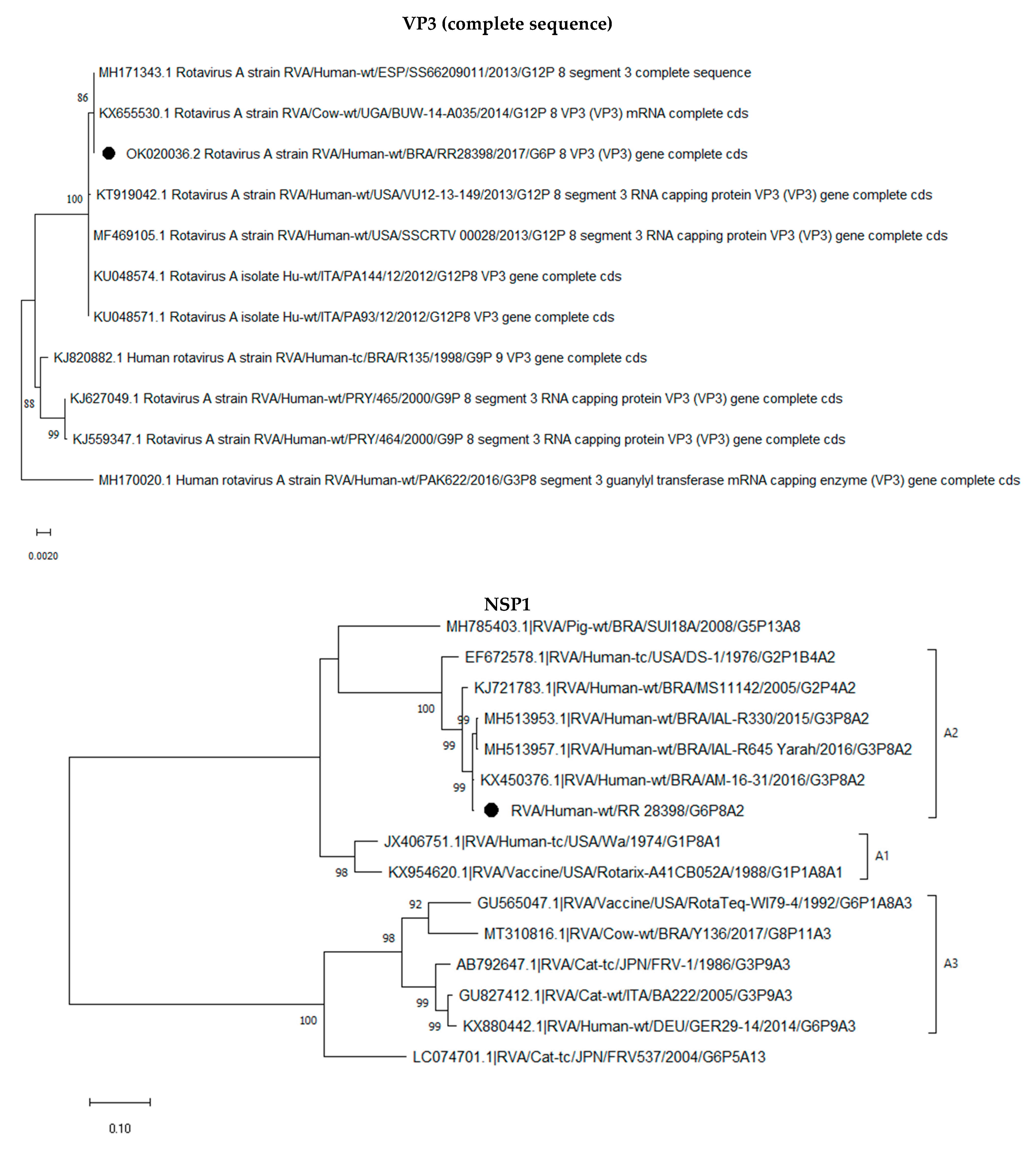
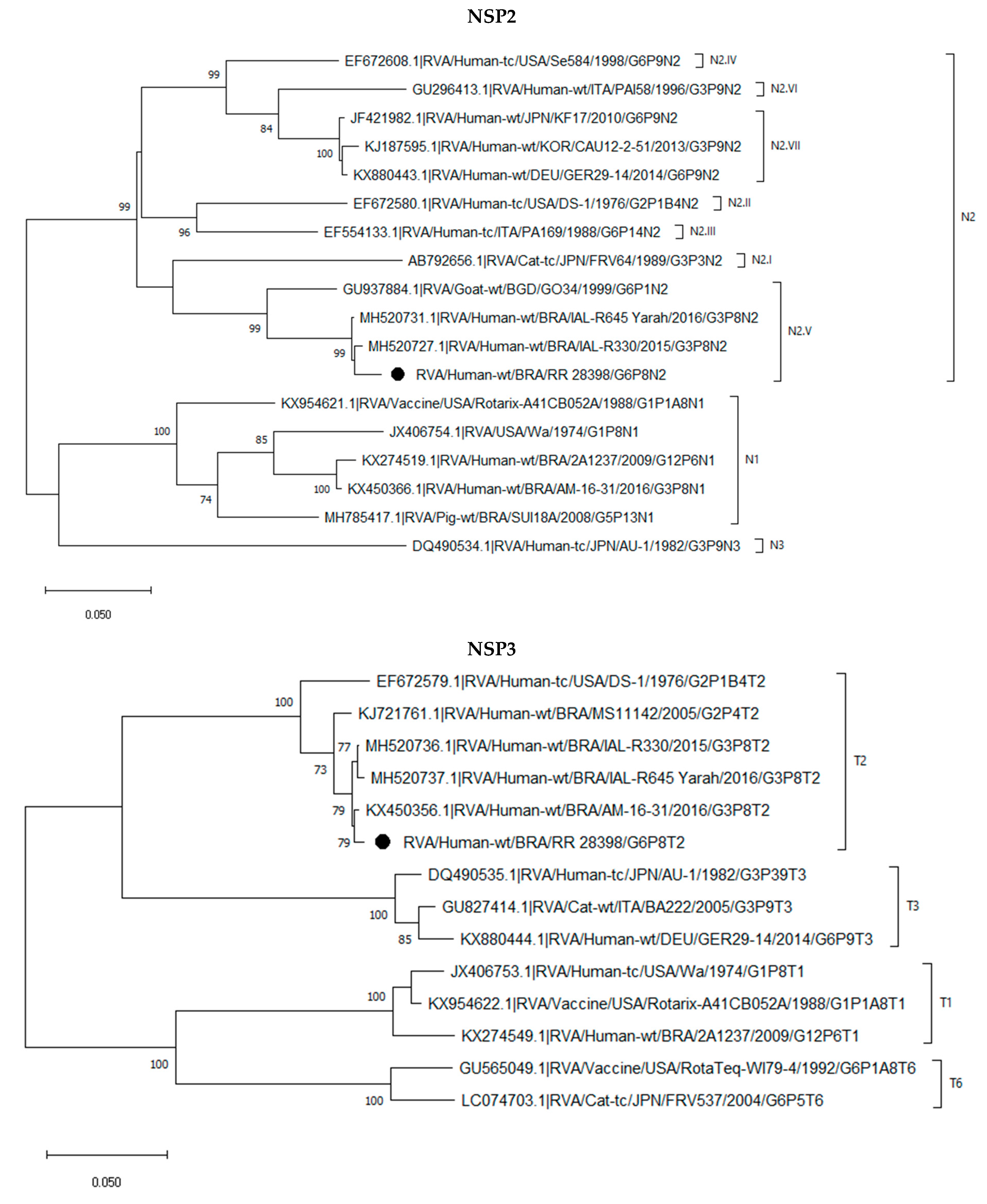
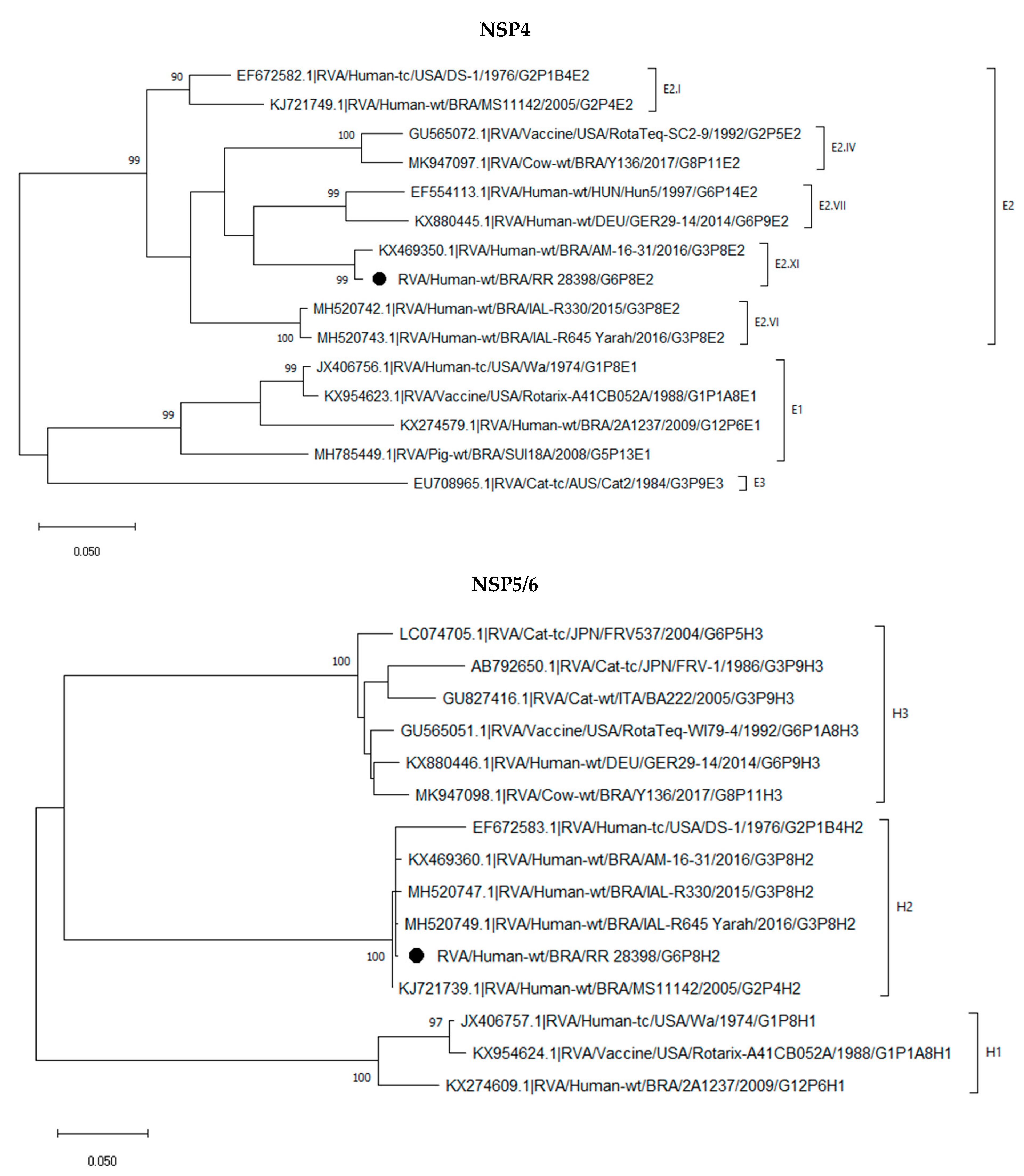
2.3. Phylogenetic Analysis of the G6P[8] Sample Shows a Constellation Possessing a Wa-like VP3
3. Discussion
4. Materials and Methods
4.1. Sample of This Study and Rotavirus A Detection
4.2. G/P Rotavirus A Genotyping
4.3. Full Genotyping of Rotavirus A ID-LVCA: 28,398 Sample
4.4. Phylogenetic Trees and RVA Genome Designation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Burden of Disease Collaborative Network. Global Burden of Disease Study 2017 (GBD 2017) Results. Available online: https://www.healthdata.org/sites/default/files/files/policy_report/2019/GBD_2017_Booklet.pdf (accessed on 30 April 2023).
- Desselberger, U. Rotaviruses. Virus Res. 2014, 190, 75–96. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Parashar, U.D. World health organization–coordinated global rotavirus surveillance network. Global, regional, and national estimates of rotavirus mortality in children <5 Years of Age, 2000–2013. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2016, 62 (Suppl. S2), S96–S105. [Google Scholar]
- Centers for Disease Control and Prevention. Rotavirus. Centers for Disease Control and Prevention. Available online: https://www.cdc.gov/vaccines/pubs/pinkbook/rota.html (accessed on 30 April 2023).
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Ciarlet, M.; Rahman, M.; Attoui, H.; Banyai, K.; Estes, M.; Gentsch, J.R.; Iturriza-Gómara, M.; Kirkwood, C.D.; Martella, V.; et al. Recommendations for the classification of group A rotaviruses using all 11 genomic RNA segments. Arch. Virol. 2008, 153, 1621–1629. [Google Scholar] [CrossRef]
- Matthijnssens, J.; Ciarlet, M.; McDonald, S.M.; Attoui, H.; Bányai, K.; Brister, J.R.; Buesa, J.; Esona, M.D.; Estes, M.K.; Gentsch, J.R.; et al. Uniformity of rotavirus strain nomenclature proposed by the Rotavirus Classification Working Group (RCWG). Arch. Virol. 2011, 156, 1397–1413. [Google Scholar] [CrossRef] [PubMed]
- PATH. Impact of Rotavirus Vaccination in the Americas. Available online: https://www.paho.org/hq/dmdocuments/2016/PAHO-GIN-2015-08.pdf (accessed on 30 April 2023).
- Gurgel, R.Q.; Cunliffe, N.A.; Nakagomi, O.; Cuevas, L.E. Rotavirus genotypes circulating in Brazil before national rotavirus vaccination: A review. J. Clin. Virol. 2008, 43, 1–8. [Google Scholar] [CrossRef]
- Gutierrez, M.B.; Fialho, A.M.; Maranhão, A.G.; Malta, F.C.; Andrade, J.S.R.A.; de Assis, R.M.S.; da Silva, E.; Mouta, S.; Miagostovich, M.P.; Leite, J.P.G.; et al. Rotavirus A in Brazil: Molecular Epidemiology and Surveillance during 2018–2019. Pathogens 2020, 9, 515. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.B.; de Assis, R.M.S.; Arantes, I.; Fumian, T.M. Full genotype constellations analysis of unusual DS-1-like G12P[6] and G6P[8] rotavirus strains detected in Brazil, 2019. Virology 2022, 577, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Kabir, H.; Miyaoka, Y.; Yamaguchi, M.; Takehara, K. G and P genotype profiles of rotavirus a field strains circulating in a vaccinated bovine farm as parameters for assessing biosecurity level. J. Vet. Med. Sci. 2022, 84, 929–937. [Google Scholar] [CrossRef]
- Medeiros, T.N.S.; Lorenzetti, E.; Alfieri, A.F.; Alfieri, A.A. G and P genotype profiles of rotavirus A field strains circulating in beef and dairy cattle herds in Brazil, 2006–2015. Comp. Immunol. Microbiol. Infect. Dis. 2019, 64, 90–98. [Google Scholar] [CrossRef]
- Rahman, M.; De Leener, K.; Goegebuer, T.; Wollants, E.; der Donck, I.V.; Hoovels, L.V.; Ranst, M.V. Genetic Characterization of a Novel, Naturally Occurring Recombinant Human G6P[6] Rotavirus. Virology 2003, 41, 2088–2095. [Google Scholar] [CrossRef] [PubMed]
- Olivares, A.I.O.; Leitão, G.A.L.; Pimenta, Y.C.; Cantelli, C.P.; Fumian, T.M.; Fialho, A.M.; e Mouta Junior, S.S.; Delgado, I.F.; Nordgren, J.; Svensson, L.; et al. Epidemiology of enteric virus infections in children living in the Amazon region. Int. J. Infect Dis. 2021, 108, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Moraes, M.T.B.; Leitão, G.A.A.L.; Olivares, A.I.O.O.; Xavier, M.P.T.P.; Bispo, R.S.; Sharma, S.; Leite, J.P.G.; Svensson, L.; Nordgren, J. Molecular Epidemiology of Sapovirus in Children Living in the Northwest Amazon Region. Pathogens 2021, 10, 965. [Google Scholar] [CrossRef] [PubMed]
- Leitão, G.A.A.; Olivares, A.I.O.; Pimenta, Y.C.; Delgado, I.F.; Miagostovich, M.P.; Leite, J.P.G.; Moraes, M.T.B. Human Bocavirus genotypes 1 and 2 detected in younger Amazonian children with acute gastroenteritis or respiratory infections, respectively. Int. J. Infect. Dis. 2020, 95, 32–37. [Google Scholar] [CrossRef]
- Bonkoungou, I.J.; Ouédraogo, N.; Tamini, L.; Teguera, R.K.; Yaméogo, P.; Drabo, M.K.; Dedah, I.; Barro, N.; Sharma, S.; Svensson, L.; et al. Rotavirus and norovirus in children with severe diarrhea in Burkina Faso before rotavirus vaccine introduction. J. Med. Virol. 2018, 90, 1453–1460. [Google Scholar] [CrossRef]
- Bwogi, J.; Jere, K.C.; Karamagi, C.; Byarugaba, D.K.; Namuwulya, P.; Baliraine, F.N.; Desselberger, U.; Iturriza-Gomara, M. Whole genome analysis of selected human and animal rotaviruses identified in Uganda from 2012 to 2014 reveals complex genome reassortment events between human, bovine, caprine and porcine strains. PLoS ONE 2017, 12, e0178855. [Google Scholar] [CrossRef]
- Alarcón, R.G.D.; Liotta, D.J.L.; Miño, S. Zoonotic RVA: State of the Art and Distribution in the Animal World. Viruses 2022, 14, 2554. [Google Scholar] [CrossRef]
- Arana, A.; Montes, M.; Jere, K.C.; Alkorta, M.; Iturriza-Gómara, M.; Cilla, G. Emergence and spread of G3P[8] rotaviruses possessing an equine-like VP7 and a DS-1-like genetic backbone in the Basque Country (North of Spain), 2015. Infect. Genet. Evol. 2016, 44, 137–144. [Google Scholar] [CrossRef]
- Guerra, S.F.S.; Lobo, P.S.; Junior Penha, E.T.P.; Junior Sousa, E.C.; Bezerra, D.A.M.; Vaz, L.R.; Linhares, A.C.; Mascarenhas, J.D.P. Detection of a novel equine-like G3 rotavirus associated with acute gastroenteritis in Brazil. J. Gen. Virol. 2016, 97, 3131–3138. [Google Scholar] [CrossRef]
- Luchs, A.; da Costa, A.C.; Ciili, A.; Komninakis, S.C.V.; Carmona, R.C.C.; Boen, L.; Morillo, S.G.; Sabino, E.C.; Timenetsky, M.C.S.T. Spread of the emerging equine-like G3P[8] DS-1-like genetic backbone rotavirus strain in Brazil and identification of potential genetic variants. J. Gen. Virol. 2019, 100, 7–25. [Google Scholar] [CrossRef]
- Silva-Sales, M.; Leal, E.; Milagres, F.A.P.; Brustulin, R.; Morais, V.S.; Marcatti, R.; Araújo, E.L.L.; Witkin, S.S.; Deng, X.; Sabino, E.C.; et al. Genomic constellation of human Rotavirus A strains identified in Northern Brazil: A 6-year follow-up (2010–2016). Rev. Inst. Med. Trop. Sao Paulo 2020, 62, e98. [Google Scholar] [CrossRef] [PubMed]
- Guerra, S.F.S.; Fecury, P.C.M.S.; Bezerra, D.A.M.; Lobo, P.S.; Penha Júnior, E.T.; Sousa Júnior, E.C.; Mascarenhas, J.D.P.; Soares, L.S.; Justino, M.C.A.; Linhares, A.C. Emergence of G12P[6] rotavirus strains among hospitalised children with acute gastroenteritis in Belém, Northern Brazil, following introduction of a rotavirus vaccine. Arch. Virol. 2019, 164, 2107–2117. [Google Scholar] [CrossRef] [PubMed]
- da Silva, L.S.; Fátima, G.S.S.; Socorro, A.O.L.; da Silva, F.S.; Fátima, E.M.C.M.; Mascarenhas, J.D.; Linhares, A.C. Diversity of rotavirus strains circulating in Northern Brazil after introduction of a rotavirus vaccine: High prevalence of G3P[6] genotype. J. Med. Virol. 2014, 86, 1065–1072. [Google Scholar] [CrossRef]
- Pietsch, C.; Liebert, U.G. Evidence for presumable feline origin of sporadic G6P[9] rotaviruses in humans. Infect. Genet. Evol. 2018, 63, 180–194. [Google Scholar] [CrossRef]
- Gómez, M.M.; Resque, H.R.; Volotão, E.M.; Rose, T.L.; da Silva, M.F.M.; Heylen, E.; Zeller, M.; Matthijnssens, J.; Leite, J.P.G. Distinct evolutionary origins of G12P[8] and G12P[9] group A rotavirus strains circulating in Brazil. Infect. Genet. Evol. 2014, 28, 385–388. [Google Scholar] [CrossRef] [PubMed]
- McKell, A.O.; Nichols, J.C.; McDonald, S.M. PCR-based approach to distinguish group A human rotavirus genotype 1 vs genotype 2 genes. J. Virol. Methods. 2013, 194, 197–205. [Google Scholar] [CrossRef]
- Rahman, M.; Matthijnssens, J.; Yang, X.; Delbeke, T.; Arijs, I.; Taniguchi, K.; Iturriza-Gómara, M.; Iftekharuddin, N.; Azim, T.; Ranst, M.V. Evolutionary history and global spread of the emerging g12 human rotaviruses. J. Virol. 2007, 81, 2382–2390. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moraes, M.T.B.d.; Silva, M.F.d.; Pimenta, Y.C.; Cantelli, C.P.; Assis, R.M.S.d.; Fialho, A.M.; Bueno, M.G.; Olivares, A.I.O.; Svensson, L.; Leite, J.P.G.; et al. G6P[8] Rotavirus a Possessing a Wa-like VP3 Gene from a Child with Acute Gastroenteritis Living in the Northwest Amazon Region. Pathogens 2023, 12, 956. https://doi.org/10.3390/pathogens12070956
Moraes MTBd, Silva MFd, Pimenta YC, Cantelli CP, Assis RMSd, Fialho AM, Bueno MG, Olivares AIO, Svensson L, Leite JPG, et al. G6P[8] Rotavirus a Possessing a Wa-like VP3 Gene from a Child with Acute Gastroenteritis Living in the Northwest Amazon Region. Pathogens. 2023; 12(7):956. https://doi.org/10.3390/pathogens12070956
Chicago/Turabian StyleMoraes, Marcia Terezinha Baroni de, Mauro França da Silva, Yan Cardoso Pimenta, Carina Pacheco Cantelli, Rosane Maria Santos de Assis, Alexandre Madi Fialho, Marina Galvão Bueno, Alberto Ignácio Olivares Olivares, Lennart Svensson, José Paulo Gagliardi Leite, and et al. 2023. "G6P[8] Rotavirus a Possessing a Wa-like VP3 Gene from a Child with Acute Gastroenteritis Living in the Northwest Amazon Region" Pathogens 12, no. 7: 956. https://doi.org/10.3390/pathogens12070956
APA StyleMoraes, M. T. B. d., Silva, M. F. d., Pimenta, Y. C., Cantelli, C. P., Assis, R. M. S. d., Fialho, A. M., Bueno, M. G., Olivares, A. I. O., Svensson, L., Leite, J. P. G., & Nordgren, J. (2023). G6P[8] Rotavirus a Possessing a Wa-like VP3 Gene from a Child with Acute Gastroenteritis Living in the Northwest Amazon Region. Pathogens, 12(7), 956. https://doi.org/10.3390/pathogens12070956