
First Record of Alternaria pogostemonis: A Novel Species Causing Leaf Spots in Pogostemon cablin



Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Pathogen Isolation
2.2. DNA Extraction and PCR Amplification
2.3. Phylogenetic Analysis
2.4. Morphological Description
2.5. Pathogenicity Tests
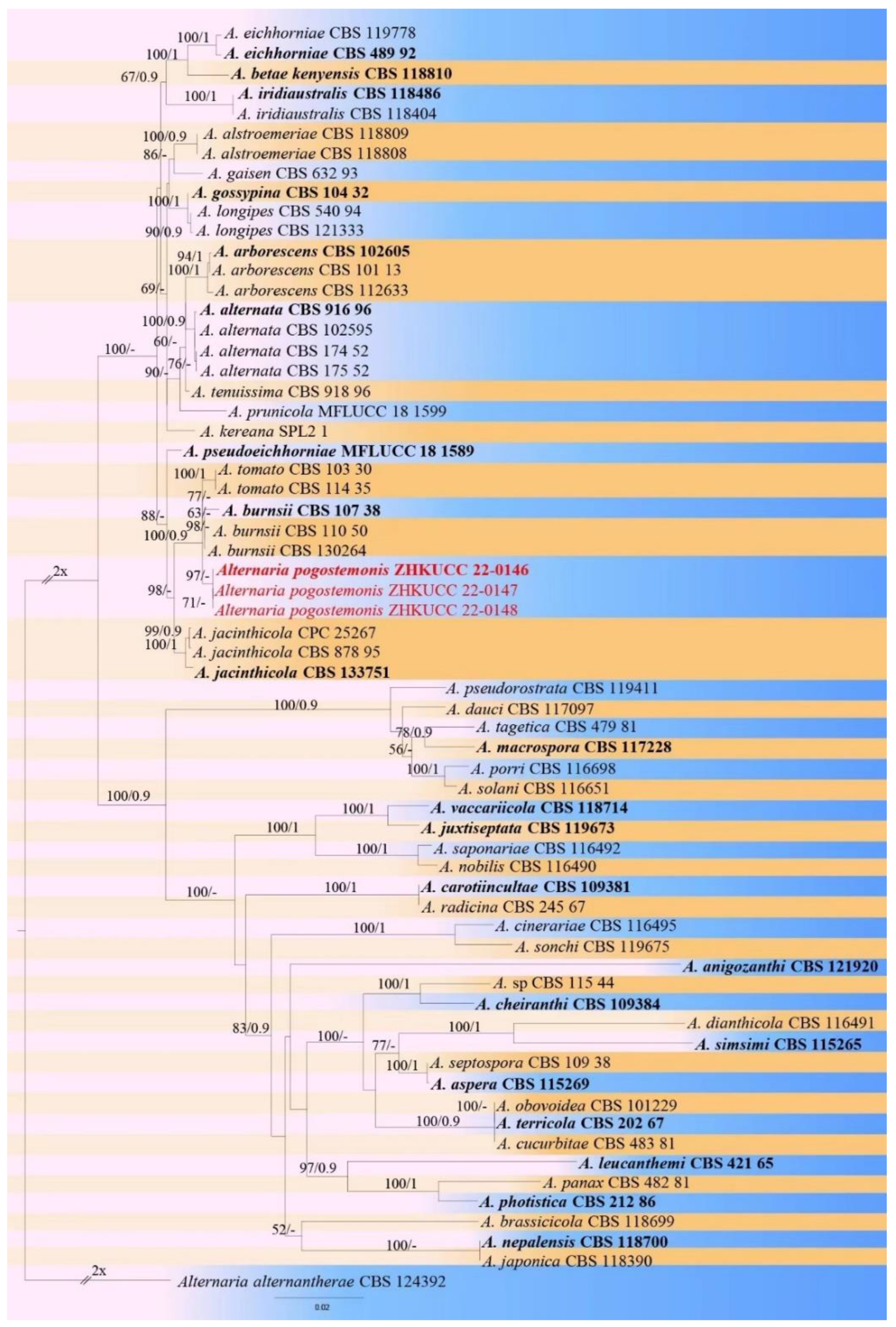
3. Results
3.1. Field Symptoms
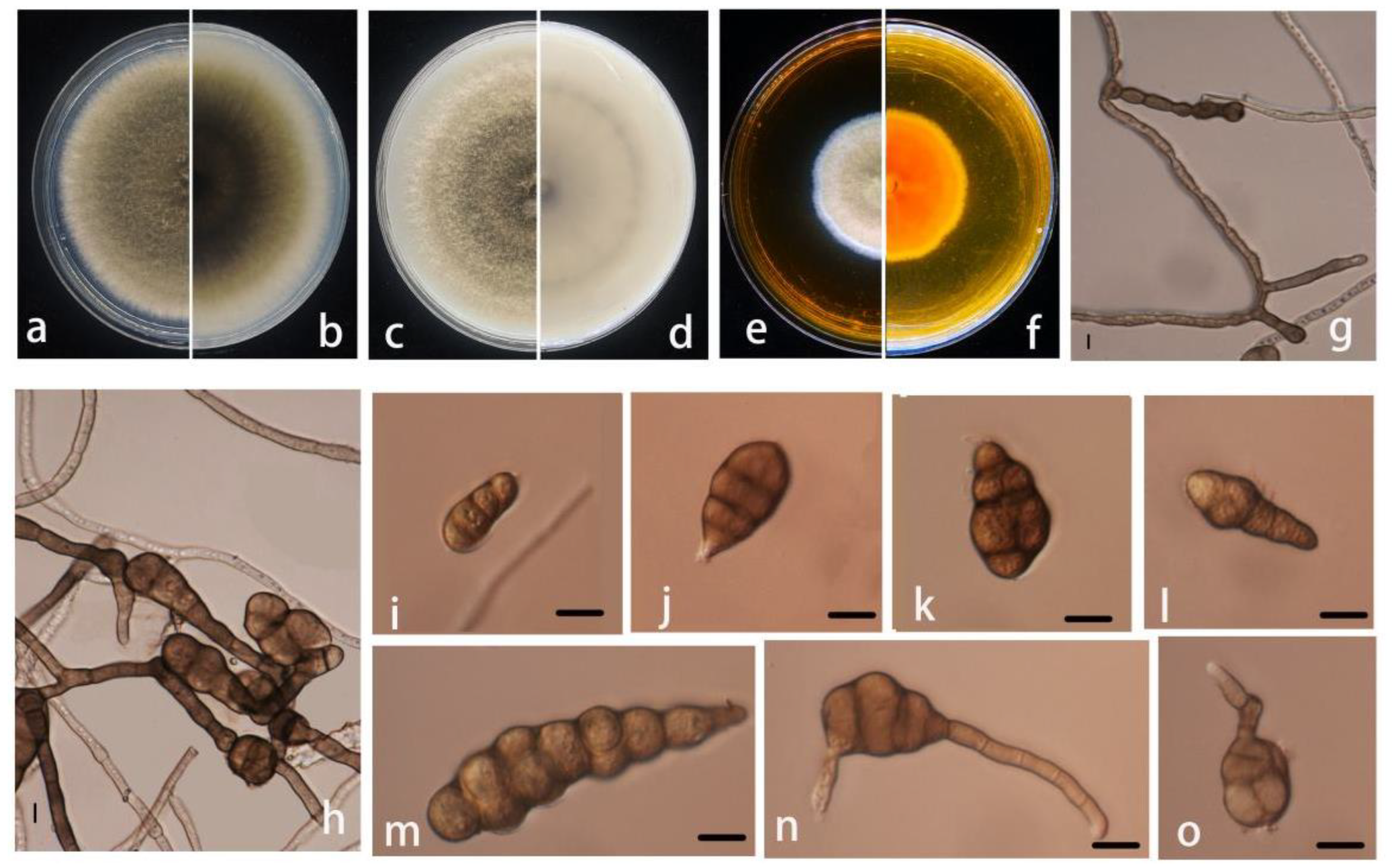
3.2. Morphological and Molecular Characterization
- Alternaria pogostemonis M. Luo, M.P. Zhao, and Z.Y. Dong, sp. nov.;
- Index Fungorum number: IF554928 (Figure 3).
- Etymology: In reference to the host genus name Pogostemon;
- Holotype: ZHKUCC 22-0146;
3.3. Disease Symptoms and Pathogenicity Tests
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dantas, T.; Cabral, T.; Neto, A.; Moura, M. Enrichmnent of patchoulol extracted from patchouli (Pogostemon cablin) oil by molecular distillation using response surface and artificial neural network models. J. Ind. Eng. Chem. 2020, 81, 219–227. [Google Scholar] [CrossRef]
- Miyazawa, M.; Okuno, Y.; Nakamura, S.I.; Kosaka, H. Antimutagenic activity of flavonoids from Pogostemon cablin. J. Agric. Food Chem. 2000, 48, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.G.; Guo, Q.S.; Zheng, H.Q. Studies on residuals of organochlorine pesticides and heavy metals in soil of planting base and Pogostemon cablin. J. Chin. Mater. Med. 2008, 33, 1528–1532. [Google Scholar]
- Arrigoni-Blank, M.; Santos, P.S.; Blank, A.F.; Brito, F.; Alves, P.B. Influence of storage temperatures and storage time of dry leaves on patchouli [Pogostemon cablin (blanco) benth.] essential oil. Biosci. J. 2020, 36, 1175–1184. [Google Scholar] [CrossRef]
- Zaim, M.; Ali, A.; Joseph, J.; Khan, F. Serological and Molecular Studies of a Novel Virus Isolate Causing Yellow Mosaic of Patchouli [Pogostemon cablin (Blanco) Benth]. PLoS ONE 2013, 8, e83790. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.J.; Wang, X.F.; Xu, X.H.; Xu, H.H.; Li, W.; Xu, L.; Deng, F.T.; Zhao, T. The classification of cultivars of Pogostemon cablin cultivated in Guangdong province of China. J. South Chin. Norm. Univ. Nat. Sci. 2003, 1, 82–86. [Google Scholar]
- Chien, J.H.; Lee, S.C.; Chang, K.F.; Huang, X.F.; Tsai, N.M. Extract of Pogostemon cablin possesses potent anticancer activity against colorectal cancer cells in vitro and in vivo. Evid.-Based Complementary Altern. Med. 2020, 3, 9758156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, G.; Li, Q.; He, L.; Zhang, Y.; Wang, Y.; He, H. Identification and characterization of virulence-attenuated mutants in Ralstonia solanacearum as potential biocontrol agents against bacterial wilt of Pogostemon cablin. Microb. Pathog. 2020, 147, 104418. [Google Scholar] [CrossRef]
- Borah, B.; Ahmed, R.; Hussain, M.; Phukon, P.; Wann, S.B.; Sarmah, D.K.; Singh, B.B. Suppression of root-knot disease in Pogostemon cablin caused by Meloidogyne incognita in a rhizobacteria mediated activation of phenylpropanoid pathway. Biol. Control 2018, 119, 43–50. [Google Scholar] [CrossRef]
- Thuy, T.T.T.; Yen, N.T.; Tuyet, N.T.A.; Te, L.L.; Waele, D.D. Population dynamics of Meloidogyne incognita on black pepper plants in two agro-ecological regions in Vietnam. Arch. Phytopathol. Plant Prot. 2012, 45, 1527–1537. [Google Scholar] [CrossRef]
- Chen, X.Y.; Sui, C.; Gan, B.C.; Wei, J.H.; Zhou, Y.K. First Report of Corynespora Leaf Spot on Patchouli Caused by Corynespora cassiicola in China. Plant Dis. 2010, 94, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.Q.; Huang, H.Q.; Lei, Z.X.; Huang, Y.C.; Huang, Y.; Fan, H.Y. First report of Phomopsis leaf spot on patchouli [Pogostemon cablin(blanco) benth.] caused by Diaporthe arecae in China. Agric. Biotechnol. 2020, 9, 18–19. [Google Scholar]
- Dong, Z.Y.; Huang, Y.H.; Manawasinghe, I.S.; Wanasinghe, D.N.; Liu, J.W.; Shu, Y.X.; Zhao, M.P.; Xiang, M.M.; Luo, M. Stagonosporopsis pogostemonis: A Novel Ascomycete Fungus Causing Leaf Spot and Stem Blight on Pogostemon cablin (Lamiaceae) in South China. Pathogens 2021, 10, 1093. [Google Scholar] [CrossRef] [PubMed]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Paul, N.C.; Deng, J.X.; Lee, H.B.; Yu, S.H. Characterization and pathogenicity of Alternaria burnsii from seeds of Cucurbita maxima (Cucurbitaceae) in Bangladesh. Mycobiology 2015, 43, 384–391. [Google Scholar] [CrossRef]
- Al-Lami, H.F.D.; You, M.P.; Barbetti, M.J. Incidence, pathogenicity and diversity of Alternaria spp. associated with Alternaria leaf spot of canola (Brassica napus) in Australia. Plant Pathol. 2019, 68, 492–503. [Google Scholar] [CrossRef]
- Nira, S.T.; Hossain, M.F.; Mahmud, N.U.; Hassan, O.; Islam, T.; Akanda, A.M. Alternaria leaf spot of broccoli caused by Alternaria alternata in Bangladesh. Plant Protect. Sci. 2022, 58, 49–56. [Google Scholar] [CrossRef]
- Chen, R.; Wang, J.Z.; Xue, X.M.; Wang, G.P. Comparison of Several Modified DNA Extraction Methods and Optimization of PCR System for Colletotrichum gloeosporioides in Apple. Acta Agric. Jiangxi 2017, 29, 69–71. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Liu, Y.L.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Hong, S.G.; Cramer, R.A.; Lawrence, C.B.; Pryor, B.M. Alt-a 1 allergen homologs from Alternaria and related taxa: Analysis of phylogenetic content and secondary structure. Fungal Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Diver. Evolut. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Hall, T.B.; Raleigh, N.C. Department of Microbiology, North Carolina State University. 2006. Available online: http://www.mbio.ncsu.edu/BioEdit/page2.html (accessed on 1 July 2022).
- Ronquist, F.R.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinf. Oxf. 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 309, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.T.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE Computer Society: Washington, DC, USA, 2010. [Google Scholar]
- Nylander, J.A.A. MrModeltest Version 2; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rao, V.G. The fungus genus Alternaria Nees. In BombayMaharashtra-1 [Dissertation]; Poona University: Pune, India, 1964. [Google Scholar]
- Simmons, E.G. Alternaria an Identification Manual; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2007; pp. 10–12. [Google Scholar]
- Cheng, H.; Zhao, L.; Wei, X.; Liu, X.; Gao, G.Y.; Deng, J.X.; Li, M.J. Alternaria species causing leaf spot on hemp (Cannabis sativa) in Northern China. Eur. J. Plant Pathol. 2022, 162, 957–970. [Google Scholar] [CrossRef]
- Meena, M.; Gupta, S.K.; Swapnil, P.; Zehra, A.; Dubey, M.K.; Upadhyay, R.S. Alternaria toxins: Potential virulence factors and genes related to pathogenesis. Front. Microbiol. 2017, 8, 1451. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, N.; Hanson, L.E.; Mambetova, S.; Jiang, Q.W.; Guza, C.; Stewart, J.; Somohana, P. Fungicide Sensitivity Monitoring of Alternaria spp. Causing Leaf Spot of Sugarbeet (Beta vulgaris) in the Upper Great Lakes. Plant Dis. 2019, 103, 2263–2270. [Google Scholar] [CrossRef]
- Saito, S.; Wang, F.; Obenland, D.; Xiao, C.L. Effects of Peroxyacetic Acid on Postharvest Diseases and Quality of Blueberries. Plant Dis. 2021, 105, 3231–3237. [Google Scholar] [CrossRef]
- Al-Nadabi, H.; Maharachchikumbura, S.; Agrama, H.; Al-Azri, M.; Nasehi, A.; Al-Sadi, A.M. Molecular characterization and pathogenicity of Alternaria species on wheat and date palms in Oman. Eur. J. Plant Pathol. 2018, 152, 577–588. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Seidl, M.F.; Groenewald, J.Z.; de Vries, M.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.H.; Kirschner, R. Diversity of Endophytic Fungi of the Coastal Plant Vitex rotundifolia in Taiwan. Microbes Environ. 2019, 34, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Manawasinghe, I.S.; Li, X.; Zhang, W.; Zhou, Y.; Hyde, K.D. Morphological and phylogenetic characterisation of endophytic fungi associated with the grapevine flowers in China. Phytotaxa 2020, 455, 95–118. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Phillips, A.J.L.; Xu, J.; Balasuriya, A.; Hyde, K.D.; Tępień, L.; Harischandra, D.L.; Karunarathna, A.; Yan, J.; Weerasinghe, J.; et al. Defining a species in plant pathology: Beyond the species level. Fungal Divers. 2021, 109, 267–282. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Primer DNA Sequence (5′–3′) | Reference |
|---|---|---|---|
| ITS | ITS 4 | TCCTCCGCTTATTGATATGC | [19] |
| ITS5 | GGAAGTAAAAGTCGTAACAAGG | ||
| gapdh | gpd1 | GCCAAGCAGTGTTGTGC | [20] |
| gpd2 | TCCTCCGCTTATTGATATGC | ||
| rpb2 | fRPB2-5F | GAYGAYMGWGATCAYTTYGG | [21] |
| fRPB2-7cR | CCCATRGCTTGTYYRCCCAT | ||
| tef1-α | TEF1-728F | CATCGAGAAGTTCGAGAAGG | [22] |
| TEF1-986R | TACTTG AAGGAACCCTTACC | ||
| Alt-a1 | Alt-F | ATGCAGTTCACCACCATCGC | [23] |
| Alt-R | ACGAGGGTGAYGTAGGCGTC |
| Species | Conidia | Pigment in PDA | References | |||
|---|---|---|---|---|---|---|
| Size (um) | Shape | Septa | ||||
| Transverse | Longitudinal | |||||
| A. burnsii | 16~60 (90) × 6.5~14 (~16) | Long ellipsoid, obclavate or ovoid | 2~6 (11) | 0~2 (~4) | None | [15] |
| A. burnsii | 25.5~105 × 8.4~20 | Obovate | 4~9 | 0~4 | No report | [30] |
| A. burnsii | 30~50 × 9~13 | Ovoid to ellipsoid | 5~8 | 1~5 | None | [31] |
| A. tomato | 30~50 × 10~13 | Narrow-ovoid | 6~9 | 1 (~2) | None | [31] |
| A. pogostemonis | 17–77 × 9–22 μm ( = 3 3 × 14 μm, n = 50) | Long ellipsoid, obclavate or ovoid | 2~7 | 0~5 | grey to grey brown | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, M.; Zhao, M.; Huang, Y.; Liu, J.; Huang, Q.; Shu, Y.; Dong, Z. First Record of Alternaria pogostemonis: A Novel Species Causing Leaf Spots in Pogostemon cablin. Pathogens 2022, 11, 1105. https://doi.org/10.3390/pathogens11101105
Luo M, Zhao M, Huang Y, Liu J, Huang Q, Shu Y, Dong Z. First Record of Alternaria pogostemonis: A Novel Species Causing Leaf Spots in Pogostemon cablin. Pathogens. 2022; 11(10):1105. https://doi.org/10.3390/pathogens11101105
Chicago/Turabian StyleLuo, Mei, Minping Zhao, Yinghua Huang, Jiawei Liu, Qiurong Huang, Yongxin Shu, and Zhangyong Dong. 2022. "First Record of Alternaria pogostemonis: A Novel Species Causing Leaf Spots in Pogostemon cablin" Pathogens 11, no. 10: 1105. https://doi.org/10.3390/pathogens11101105
APA StyleLuo, M., Zhao, M., Huang, Y., Liu, J., Huang, Q., Shu, Y., & Dong, Z. (2022). First Record of Alternaria pogostemonis: A Novel Species Causing Leaf Spots in Pogostemon cablin. Pathogens, 11(10), 1105. https://doi.org/10.3390/pathogens11101105