
Determination of Intestinal Viral Loads and Distribution of Bovine Viral Diarrhea Virus, Classical Swine Fever Virus, and Peste Des Petits Ruminants Virus: A Pilot Study

 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
Abstract
1. Introduction
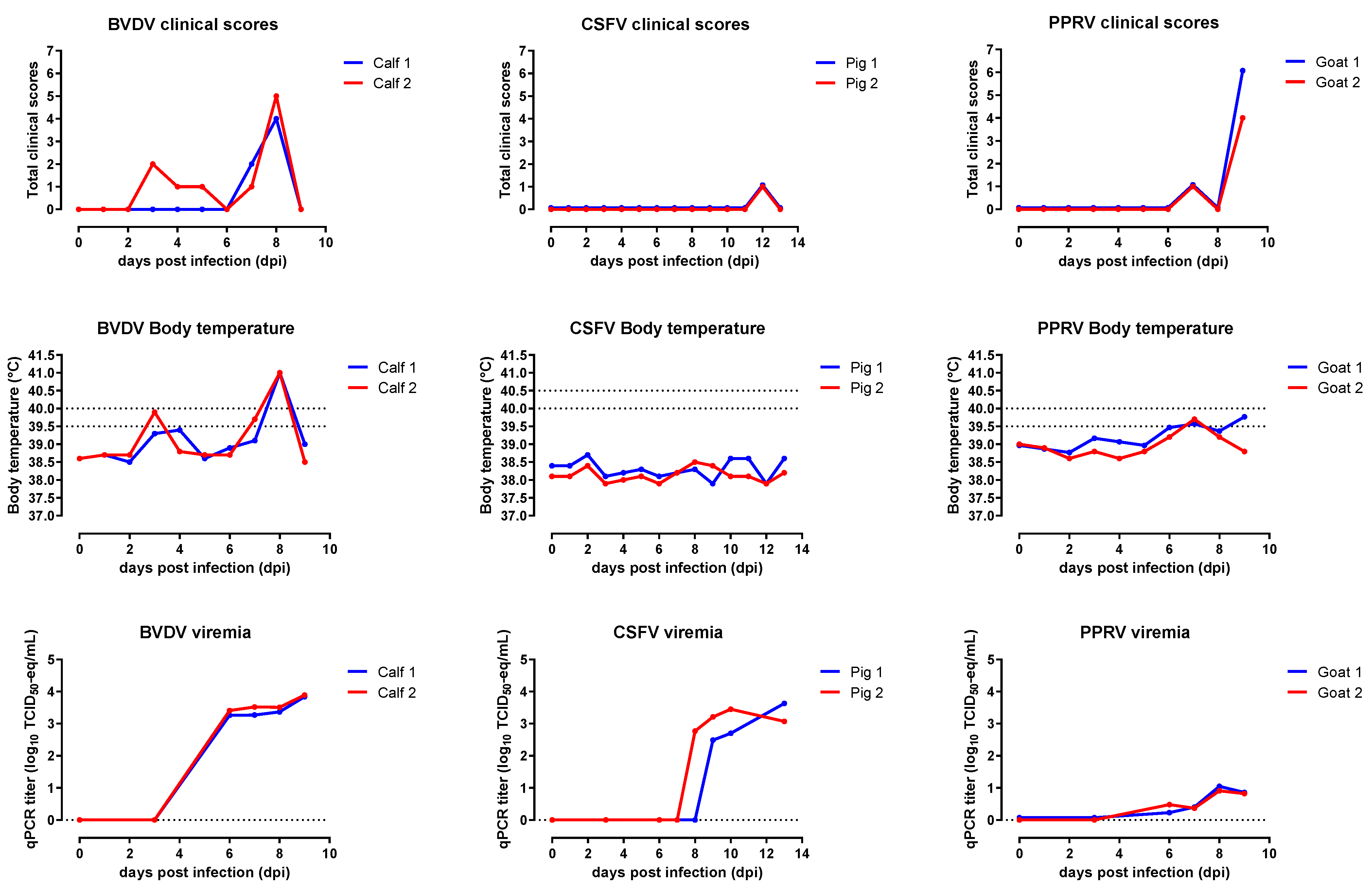
2. Results
2.1. Animal Studies
2.1.1. BVDV
2.1.2. CSFV
2.1.3. PPRV
2.2. Intestinal Viral Loads
2.2.1. BVDV
2.2.2. CSFV
2.2.3. PPRV
3. Discussion
4. Materials and Methods
4.1. Viruses
4.2. Animal Studies
4.3. Intestine Sampling
4.4. Intestinal Viral Loads
4.5. Quantitative RT-PCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wijnker, J.J.; Haas, B.; Berends, B.R. Removal of foot-and-mouth disease virus infectivity in salted natural casings by minor adaptation of standardized industrial procedures. Int. J. Food Microbiol. 2007, 115, 214–219. [Google Scholar] [CrossRef]
- Wijnker, J.J.; Haas, B.; Berends, B.R. Inactivation of foot-and-mouth disease virus in various bovine tissues used for the production of natural sausage casings. Int. J. Food Microbiol. 2012, 153, 237–240. [Google Scholar] [CrossRef]
- Wieringa-Jelsma, T.; Wijnker, J.J.; Zijlstra-Willems, E.M.; Dekker, A.; Stockhofe-Zurwieden, N.; Maas, R.; Wisselink, H.J. Virus inactivation by salt (NaCl) and phosphate supplemented salt in a 3D collagen matrix model for natural sausage casings. Int. J. Food Microbiol. 2011, 148, 128–134. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on animal health risk mitigation treatments as regards imports of animal casings. EFSA J. 2012, 10, 2820. [Google Scholar]
- Jelsma, T.; Wijnker, J.J.; Smid, B.; Verheij, E.; Poel, W.H.M.v.d.; Wisselink, H.J. Salt inactivation of classical swine fever virus and African swine fever virus in porcine intestines confirms the existing in vitro casings model. Vet. Microbiol. 2019, 238, 108424. [Google Scholar] [CrossRef] [PubMed]
- Jelsma, T.; Wijnker, J.J.; van der Poel, W.H.M.; Wisselink, H.J. Intestinal Viral Loads and Inactivation Kinetics of Livestock Viruses Relevant for Natural Casing Production: A Systematic Review and Meta-Analysis. Pathogens 2021, 10, 173. [Google Scholar] [CrossRef] [PubMed]
- Pedrera, M.; Gómez-Villamandos, J.C.; Molina, V.; Risalde, M.A.; Rodríguez-Sánchez, B.; Sánchez-Cordón, P.J. Quantification and determination of spread mechanisms of bovine viral diarrhoea virus in blood and tissues from colostrum-deprived calves during an experimental acute infection induced by a non-cytopathic genotype 1 strain. Transbound. Emerg. Dis. 2012, 59, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Marshall, D.J.; Moxley, R.A.; Kelling, C.L. Distribution of Virus and Viral Antigen in Specific Pathogen-free Calves Following Inoculation with Noncytopathic Bovine Viral Diarrhea Virus. Vet. Pathol. 1996, 33, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Bruschke, C.J.M.; Weerdmeester, K.; Van Oirschot, J.T.; Van Rijn, P.A. Distribution of bovine virus diarrhoea virus in tissues and white blood cells of cattle during acute infection. Vet. Microbiol. 1998, 64, 23–32. [Google Scholar] [CrossRef]
- Wood, L.; Brockman, S.; Harkness, J.; Edwards, S. Classical swine fever: Virulence and tissue distribution of a 1986 English isolate in pigs. Vet. Rec. 1988, 122, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Dahle, J.; Liess, B. A review on classical swine fever infections in pigs: Epizootiology, clinical disease and pathology. Comp. Immunol. Microbiol. Infect. Dis. 1992, 15, 203–211. [Google Scholar] [CrossRef]
- Blome, S.; Staubach, C.; Henke, J.; Carlson, J.; Beer, M. Classical Swine Fever—An Updated Review. Viruses 2017, 9, 86. [Google Scholar] [CrossRef] [PubMed]
- CFSPH_CSFV. CSFV. 2015. Available online: https://www.cfsph.iastate.edu/Factsheets/pdfs/classical_swine_fever.pdf (accessed on 5 September 2021).
- Moennig, V.; Floegel-Niesmann, G.; Greiser-Wilke, I. Clinical Signs and Epidemiology of Classical Swine Fever: A Review of New Knowledge. Vet. J. 2003, 165, 11–20. [Google Scholar] [CrossRef]
- OIE_CSFV. 2019. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.08.03_CSF.pdf (accessed on 5 September 2021).
- Narita, M.; Kawashima, K.; Kimura, K.; Mikami, O.; Shibahara, T.; Yamada, S.; Sakoda, Y. Comparative Immunohistopathology in Pigs Infected with Highly Virulent or Less Virulent Strains of Hog Cholera Virus. Vet. Pathol. 2000, 37, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Izzati, U.Z.; Hoa, N.T.; Lan, N.T.; Diep, N.V.; Fuke, N.; Hirai, T.; Yamaguchi, R. Pathology of the outbreak of subgenotype 2.5 classical swine fever virus in northern Vietnam. Vet. Med. Sci. 2021, 7, 164–174. [Google Scholar] [CrossRef]
- McKercher, P.D.; Morgan, D.O.; McVicar, J.W.; Shuot, N.J. Thermal processing to inactivate viruses in meat products. In Proceedings of the 84th United States Animal Health Association, Louisville, KY, USA, 2–7 November 1980; Volume 84, pp. 320–328. [Google Scholar]
- Helwig, D.M.; Keast, J.C. Viability of virulent swine fever virus in cooked and uncooked ham and sausage casings. Aust. Vet. J. 1966, 42, 131–135. [Google Scholar] [CrossRef]
- Wijnker, J.J.; Depner, K.R.; Berends, B.R. Inactivation of classical swine fever virus in porcine casing preserved in salt. Int. J. Food Microbiol. 2008, 128, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Begum, S.; Nooruzzaman, M.; Islam, M.R.; Chowdhury, E.H. A Sequential Study on the Pathology of Peste Des Petits Ruminants and Tissue Distribution of the Virus Following Experimental Infection of Black Bengal Goats. Front. Vet. Sci. 2021, 8, 635671. [Google Scholar] [CrossRef]
- Kul, O.; Kabakci, N.; Atmaca, H.T.; Ozkul, A. Natural peste des petits ruminants virus infection: Novel pathologic findings resembling other morbillivirus infections. Vet. Pathol. 2007, 44, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Tripathi, B.N.; Sharma, A.K.; Kumar, R.; Sreenivasa, B.P.; Singh, R.P.; Dhar, P.; Bandyopadhyay, S.K. Pathological and immunohistochemical study of experimental peste des petits ruminants virus infection in goats. J. Vet. Med. Ser. B Infect. Dis. Vet. Public Health 2004, 51, 153–159. [Google Scholar] [CrossRef]
- Pope, R.A.; Parida, S.; Bailey, D.; Brownlie, J.; Barrett, T.; Banyard, A.C. Early Events following Experimental Infection with Peste-Des-Petits Ruminants Virus Suggest Immune Cell Targeting. PLoS ONE 2013, 8, e55830. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, M.; Dinesh, M.; Thakor, J.C.; Baloni, S.; Saxena, S.; Shrivastava, S.; Dhama, K.; Singh, K.; Singh, R. Neuropathology mediated through caspase dependent extrinsic pathway in goat kids naturally infected with PPRV. Microb. Pathog. 2020, 140, 103949. [Google Scholar] [CrossRef] [PubMed]
- Toplu, N. Characteristic and non-characteristic pathological findings in peste des petits ruminants (PPR) of sheep in the Ege district of Turkey. J. Comp. Pathol. 2004, 131, 135–141. [Google Scholar] [CrossRef]
- Jagtap, S.P.; Rajak, K.K.; Garg, U.K.; Sen, A.; Bhanuprakash, V.; Sudhakar, S.B.; Balamurugan, V.; Patel, A.; Ahuja, A.; Singh, R.K.; et al. Effect of immunosuppression on pathogenesis of peste des petits ruminants (PPR) virus infection in goats. Microb. Pathog. 2012, 52, 217–226. [Google Scholar] [CrossRef]
- Truong, T.; Boshra, H.; Embury-Hyatt, C.; Nfon, C.; Gerdts, V.; Tikoo, S.; Babiuk, L.A.; Kara, P.; Chetty, T.; Mather, A.; et al. Peste des petits ruminants virus tissue tropism and pathogenesis in sheep and goats following experimental infection. PLoS ONE 2014, 9, e87145. [Google Scholar] [CrossRef]
- Schweizer, M.; Peterhans, E. Pestiviruses. Ann. Rev. Anim. Biosci. 2014, 2, 141–163. [Google Scholar] [CrossRef] [PubMed]
- Baron, M.D.; Diallo, A.; Lancelot, R.; Libeau, G. Chapter 1: Peste des Petits Ruminants Virus. In Advances in Virus Research; Kielian, M., Maramorosch, K., Mettenleiter, T.C., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 95, pp. 1–42. [Google Scholar]
- Greiser-Wilke, I.; Zimmermann, B.; Fritzemeier, J.; Floegel, G.; Moennig, V. Structure and presentation of a World Wide Web database of CSF virus isolates held at the EU Reference Laboratory. Vet. Microbiol. 2000, 73, 131–136. [Google Scholar] [CrossRef]
- Bruschke, C.J.M.; Van Rijn, P.A.; Moormann, R.J.M.; Van Oirschot, J.T. Antigenically different pestivirus strains induce congenital infection in sheep: A model for bovine virus diarrhea virus vaccine efficacy studies. Vet. Microbiol. 1996, 50, 33–43. [Google Scholar] [CrossRef]
- Baron, J.; Bin-Tarif, A.; Herbert, R.; Frost, L.; Taylor, G.; Baron, M.D. Early changes in cytokine expression in peste des petits ruminants disease. Vet. Res. 2014, 45, 1–11. [Google Scholar] [CrossRef] [PubMed]
- El Harrak, M.; Touil, N.; Loutfi, C.; Hammouchi, M.; Parida, S.; Sebbar, G.; Chaffai, N.; Harif, B.; Messoudi, N.; Batten, C.; et al. A reliable and reproducible experimental challenge model for peste des petits ruminants virus. J. Clin. Microbiol. 2012, 50, 3738–3740. [Google Scholar] [CrossRef]
- Weesendorp, E.; Backer, J.; Stegeman, A.; Loeffen, W. Effect of strain and inoculation dose of classical swine fever virus on within-pen transmission. Vet. Res. 2009, 40, 59. [Google Scholar] [CrossRef][Green Version]
- Herbert, R.; Baron, J.; Batten, C.; Baron, M.; Taylor, G. Recombinant adenovirus expressing the haemagglutinin of peste des petits ruminants virus (PPRV) protects goats against challenge with pathogenic virus; a DIVA vaccine for PPR. Vet. Res. 2014, 45, 24. [Google Scholar] [CrossRef] [PubMed]
- Strong, R.; La Rocca, S.A.; Paton, D.; Bensaude, E.; Sandvik, T.; Davis, L.; Turner, J.; Drew, T.; Raue, R.; Vangeel, I.; et al. Viral Dose and Immunosuppression Modulate the Progression of Acute BVDV-1 Infection in Calves: Evidence of Long Term Persistence after Intra-Nasal Infection. PLoS ONE 2015, 10, e0124689. [Google Scholar] [CrossRef]
- Mittelholzer, C.; Moser, C.; Tratschin, J.D.; Hofmann, M.A. Analysis of classical swine fever virus replication kinetics allows differentiation of highly virulent from avirulent strains. Vet. Microbiol. 2000, 74, 293–308. [Google Scholar] [CrossRef]
- Koolmees, P.A.; Tersteeg, M.H.G.; Keizer, G.; Van Den Broek, J.; Bradley, R. Comparative histological studies of mechanically versus manually processed sheep intestines used to make natural sausage casings. J. Food Prot. 2004, 67, 2747–2755. [Google Scholar] [CrossRef] [PubMed]
- Koolmees, P.A.; Houben, J.H. Inventory part: Histology and microbiology of hog and sheep casings. In Colour Print Leaflet EU—CRAFT Project BRE 2. CT 94. 1495: Improved Treatment of Natural Sausage Casings for Quality Improvement in Automated Stuffing Processes; Deutches Institut für Lebensmitteltechnik e.V.: Quackenbrück, Germany, 1997. [Google Scholar]
- Wijnker, J.; Tersteeg, M.; Berends, B.; Vernooij, J.; Koolmees, P. Quantitative Histological Analysis of Bovine Small Intestines before and after Processing into Natural Sausage Casings. J. Food Prot. 2008, 71, 1199–1204. [Google Scholar] [CrossRef] [PubMed]
- ENSCA. Available online: http://www.ensca.eu/index.php?/eng/content/download/2936/22121/file/ENSCA%20cGGP%20revision%20VIII%2018-06-19.pdf (accessed on 15 March 2021).
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef]
- Van Rijn, P.A.; Wellenberg, G.J.; Hakze-Van Der Honing, R.; Jacobs, L.; Moonen, P.L.J.M.; Feitsma, H. Detection of economically important viruses in boar semen by quantitative RealTime PCR™ technology. J. Virol. Methods 2004, 120, 151–160. [Google Scholar] [CrossRef]
- Weesendorp, E.; Backer, J.; Loeffen, W. Quantification of different classical swine fever virus transmission routes within a single compartment. Vet. Microbiol. 2014, 174, 353–361. [Google Scholar] [CrossRef]
- van Rijn, P.A.; Boonstra, J.; van Gennip, H.G.P. Recombinant Newcastle disease viruses with targets for PCR diagnostics for rinderpest and peste des petits ruminants. J. Virol. Methods 2018, 259, 50–53. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jelsma, T.; Wijnker, J.J.; Smid, B.; Verheij, E.; van der Poel, W.H.M.; Wisselink, H.J. Determination of Intestinal Viral Loads and Distribution of Bovine Viral Diarrhea Virus, Classical Swine Fever Virus, and Peste Des Petits Ruminants Virus: A Pilot Study. Pathogens 2021, 10, 1188. https://doi.org/10.3390/pathogens10091188
Jelsma T, Wijnker JJ, Smid B, Verheij E, van der Poel WHM, Wisselink HJ. Determination of Intestinal Viral Loads and Distribution of Bovine Viral Diarrhea Virus, Classical Swine Fever Virus, and Peste Des Petits Ruminants Virus: A Pilot Study. Pathogens. 2021; 10(9):1188. https://doi.org/10.3390/pathogens10091188
Chicago/Turabian StyleJelsma, Tinka, Joris J. Wijnker, Bregtje Smid, Eline Verheij, Wim H. M. van der Poel, and Henk J. Wisselink. 2021. "Determination of Intestinal Viral Loads and Distribution of Bovine Viral Diarrhea Virus, Classical Swine Fever Virus, and Peste Des Petits Ruminants Virus: A Pilot Study" Pathogens 10, no. 9: 1188. https://doi.org/10.3390/pathogens10091188
APA StyleJelsma, T., Wijnker, J. J., Smid, B., Verheij, E., van der Poel, W. H. M., & Wisselink, H. J. (2021). Determination of Intestinal Viral Loads and Distribution of Bovine Viral Diarrhea Virus, Classical Swine Fever Virus, and Peste Des Petits Ruminants Virus: A Pilot Study. Pathogens, 10(9), 1188. https://doi.org/10.3390/pathogens10091188