
Medroxyprogesterone Acetate (MPA) Enhances HIV-1 Accumulation and Release in Primary Cervical Epithelial Cells by Inhibiting Lysosomal Activity


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
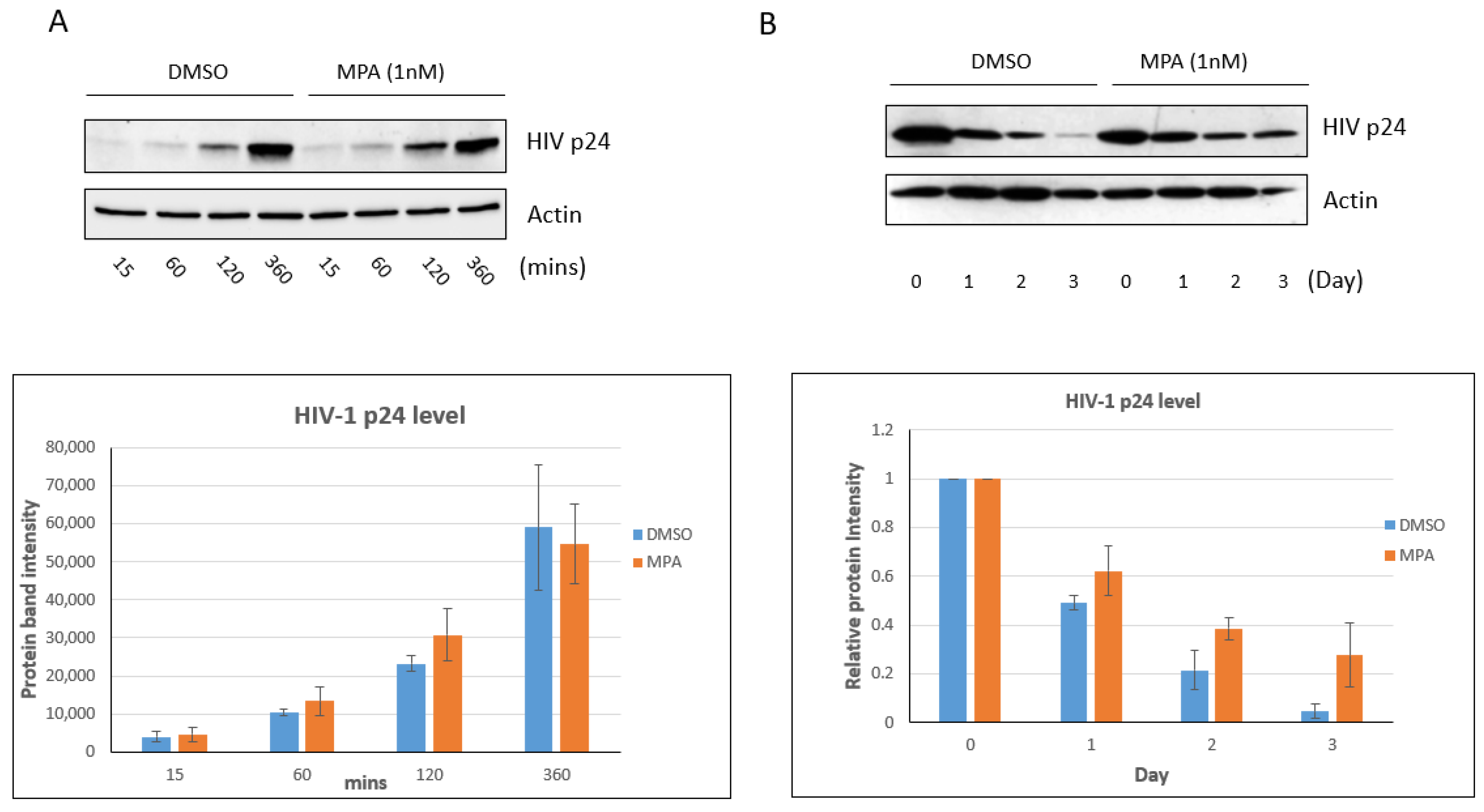
2.1. MPA Treatment Enhances Intracellular HIV-1 Accumulation
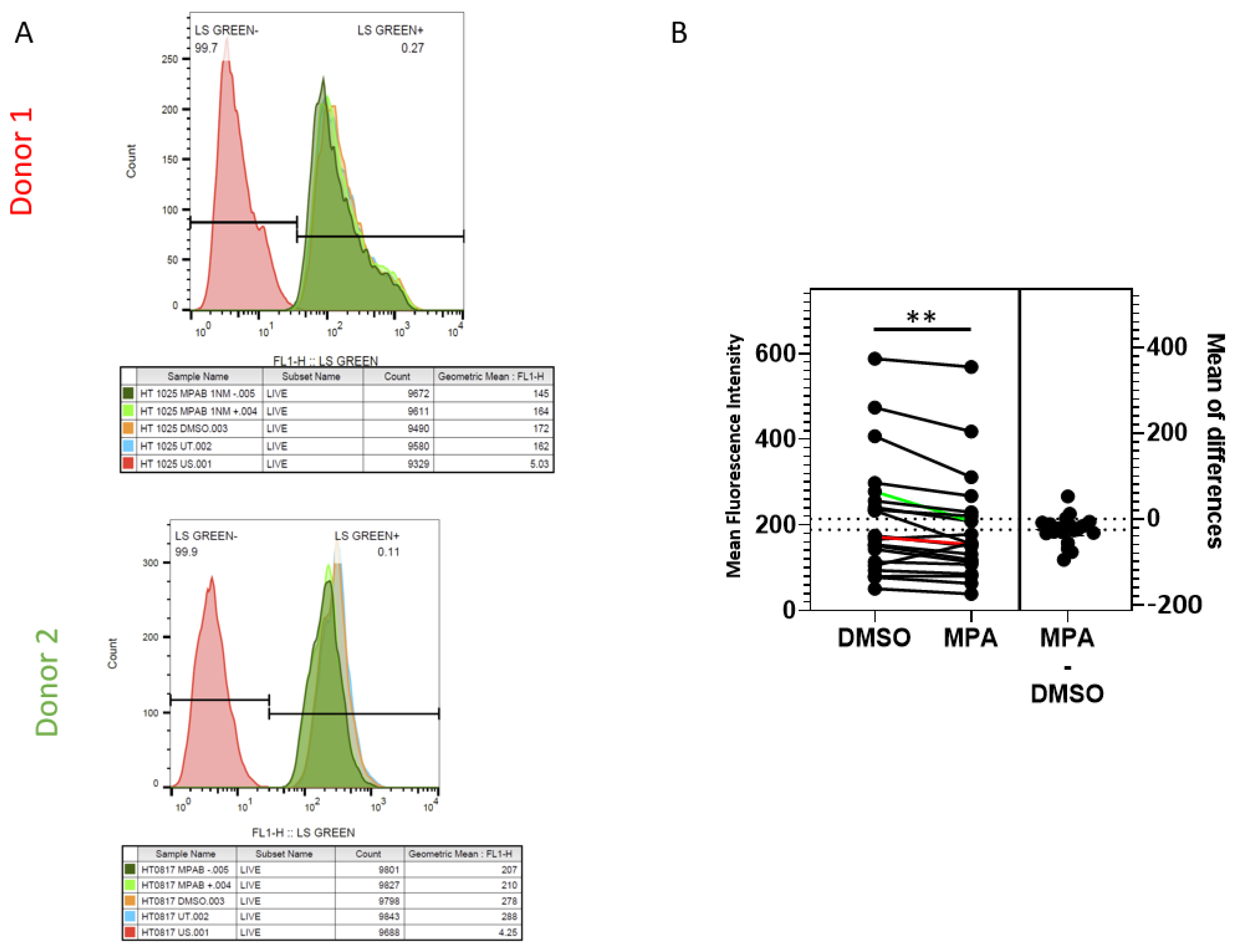
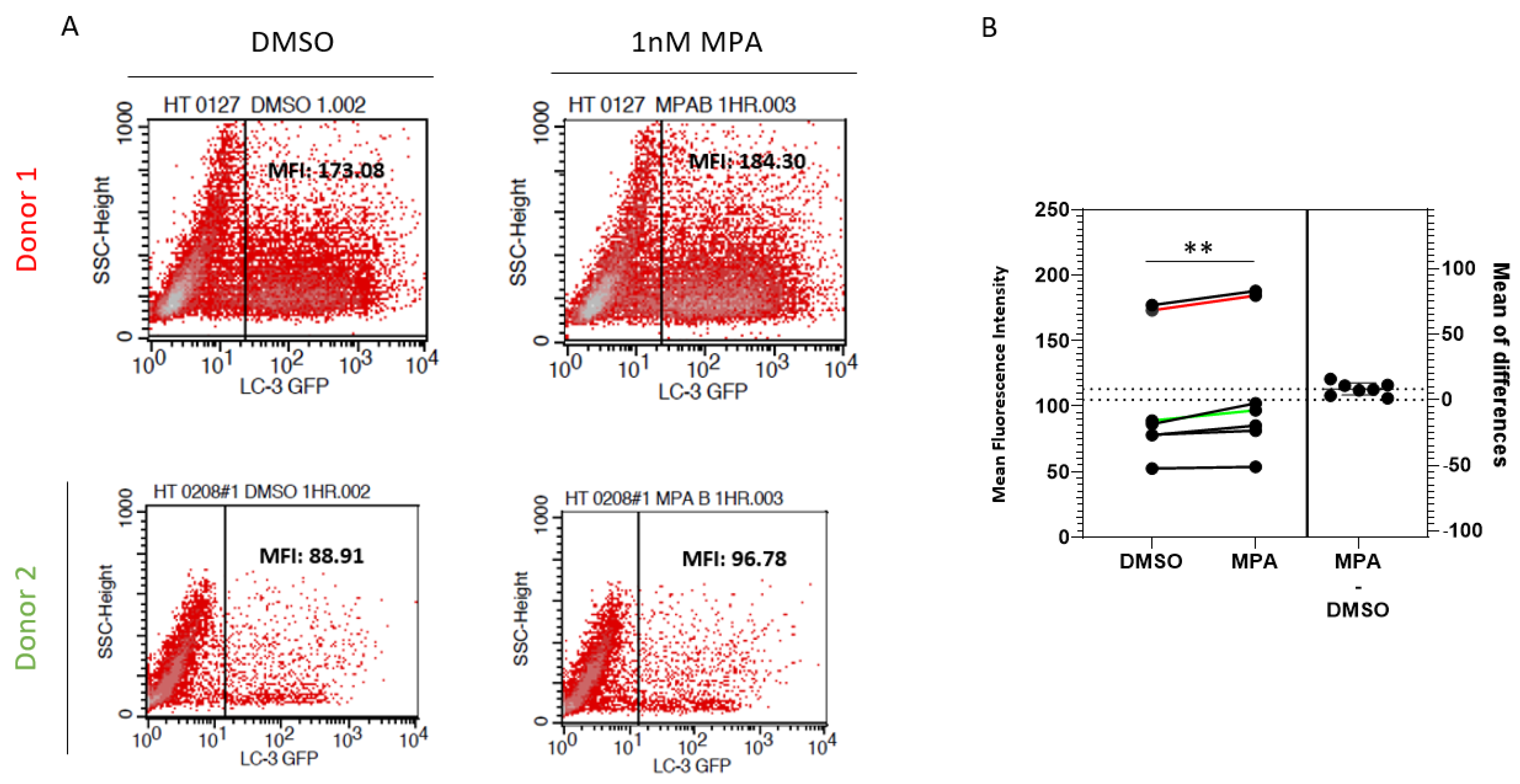
2.2. MPA Reduces Lysosomal Activity
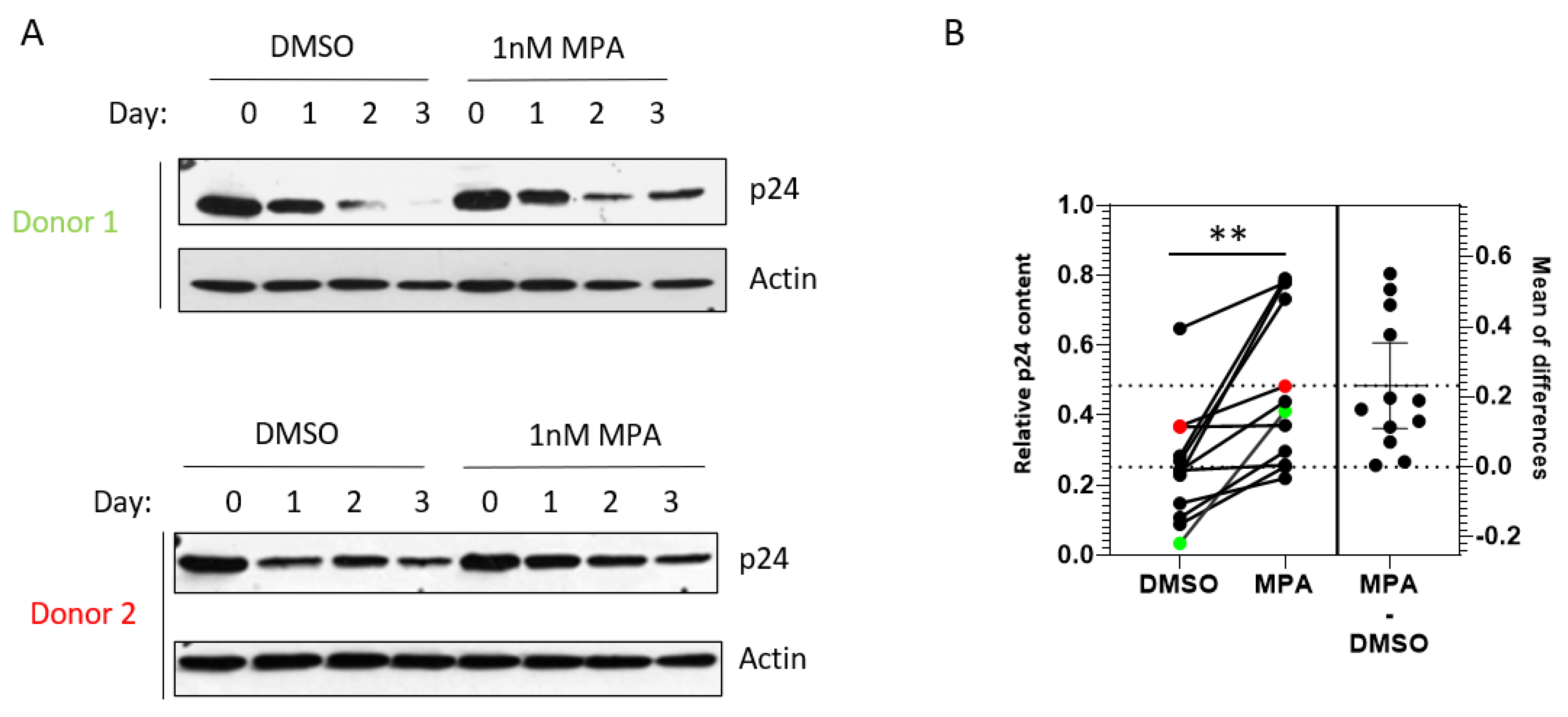
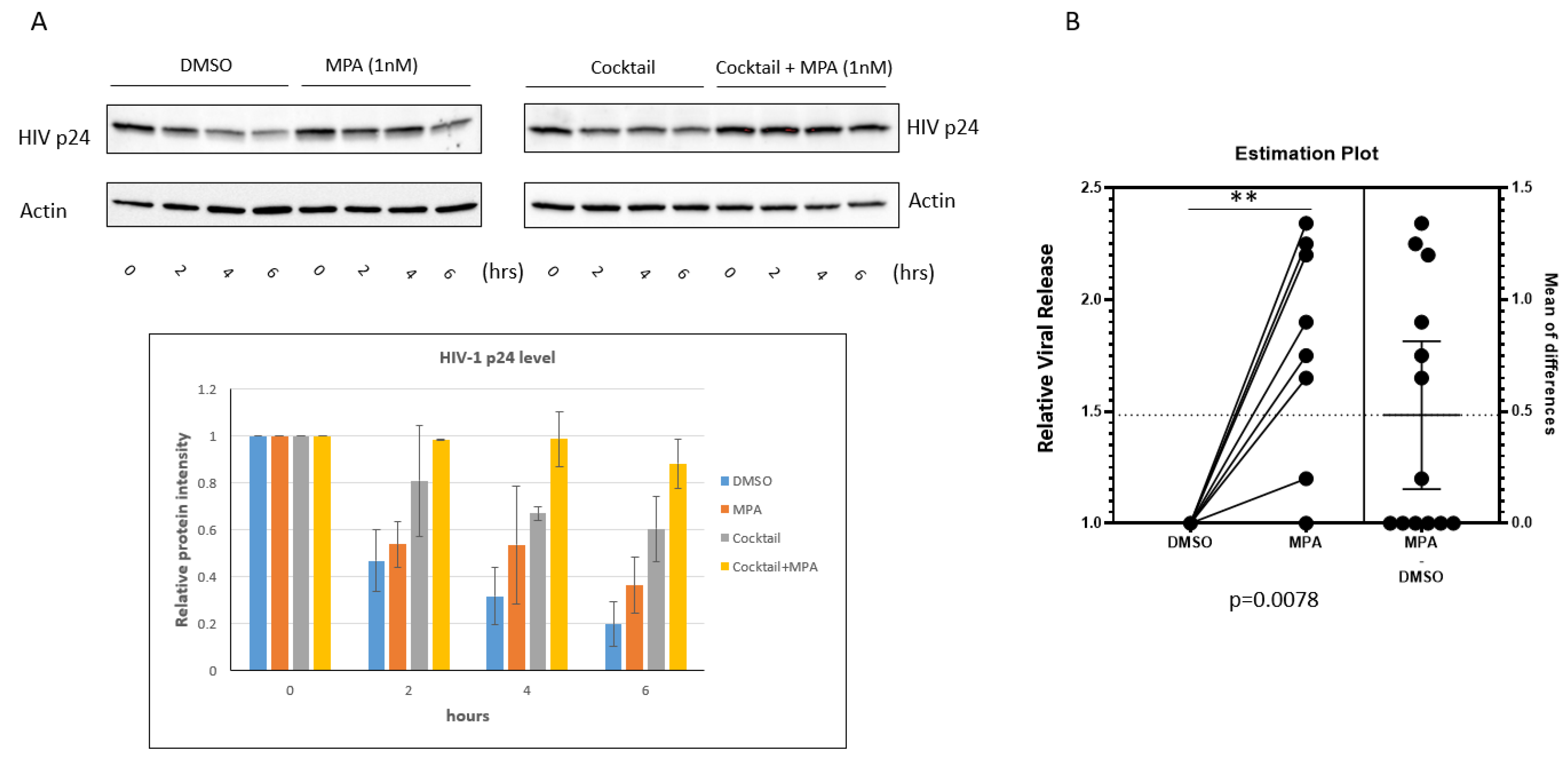
2.3. MPA Enhances HIV-1 Release from Human Primary Cervical Epithelial Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines, HIV-1 Virus, Plasmids, Reagents, and Antibodies
4.2. Source of Tissues as Well as Cervical Epithelial Cell Isolation and Culture
4.3. MPA, Lysosomal Inhibitors Cocktail Treatment and Viral Inoculation
4.4. SDS-PAGE and Western Blot
4.5. LysoSensorTM Staining
4.6. Transfection and Flow Cytometry Analysis
4.7. qRT-PCR
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tebit, D.M.; Ndembi, N.; Weinberg, A.; Quiñones-Mateu, M.E. Mucosal transmission of human immunodeficiency virus. Curr. HIV Res. 2012, 10, 3–8. [Google Scholar] [CrossRef]
- Hladik, F.; McElrath, M.J. Setting the stage: Host invasion by HIV. Nat. Rev. Immunol. 2008, 8, 447–457. [Google Scholar] [CrossRef]
- Nicolosi, A.; Correa Leite, M.L.; Musicco, M.; Arici, C.; Gavazzeni, G.; Lazzarin, A. The efficiency of male-to-female and female-to-male sexual transmission of the human immunodeficiency virus: A study of 730 stable couples. Italian Study Group on HIV Heterosexual Transmission. Epidemiology 1994, 5, 570–575. [Google Scholar] [CrossRef]
- Hel, Z.; Stringer, E.; Mestecky, J. Sex steroid hormones, hormonal contraception, and the immunobiology of human immunodeficiency virus-1 infection. Endocr. Rev. 2010, 31, 79–97. [Google Scholar] [CrossRef]
- Wand, H.; Ramjee, G. The effects of injectable hormonal contraceptives on HIV seroconversion and on sexually transmitted infections. Aids 2012, 26, 375–380. [Google Scholar] [CrossRef]
- Martin, H.L., Jr.; Nyange, P.M.; Richardson, B.A.; Lavreys, L.; Mandaliya, K.; Jackson, D.J.; Ndinya-Achola, J.O.; Kreiss, J. Hormonal contraception, sexually transmitted diseases, and risk of heterosexual transmission of human immunodeficiency virus type 1. J. Infect. Dis. 1998, 178, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Kiddugavu, M.; Makumbi, F.; Wawer, M.J.; Serwadda, D.; Sewankambo, N.K.; Wabwire-Mangen, F.; Lutalo, T.; Meehan, M.; Xianbin; Gray, R.H. Hormonal contraceptive use and HIV-1 infection in a population-based cohort in Rakai, Uganda. Aids 2003, 17, 233–240. [Google Scholar] [CrossRef]
- Polis, C.B.; Curtis, K.M.; Hannaford, P.C.; Phillips, S.J.; Chipato, T.; Kiarie, J.N.; Westreich, D.J.; Steyn, P.S. An updated systematic review of epidemiological evidence on hormonal contraceptive methods and HIV acquisition in women. Aids 2016, 30, 2665–2683. [Google Scholar] [CrossRef] [PubMed]
- Morrison, C.S.; Chen, P.L.; Kwok, C.; Baeten, J.M.; Brown, J.; Crook, A.M.; van Damme, L.; Delany-Moretlwe, S.; Francis, S.C.; Friedland, B.A.; et al. Hormonal contraception and the risk of HIV acquisition: An individual participant data meta-analysis. PLoS Med. 2015, 12, e1001778. [Google Scholar] [CrossRef] [PubMed]
- Ralph, L.J.; McCoy, S.I.; Shiu, K.; Padian, N.S. Hormonal contraceptive use and women’s risk of HIV acquisition: A meta-analysis of observational studies. Lancet Infect. Dis. 2015, 15, 181–189. [Google Scholar] [CrossRef]
- Evidence for Contraceptive O, Consortium HIVOT: HIV incidence among women using intramuscular depot medroxyprogesterone acetate, a copper intrauterine device, or a levonorgestrel implant for contraception: A randomised, multicentre, open-label trial. Lancet 2019, 394, 303–313. [CrossRef]
- Hapgood, J.P. Is the Injectable Contraceptive Depo-Medroxyprogesterone Acetate (DMPA-IM) Associated with an Increased Risk for HIV Acquisition? The Jury Is Still Out. AIDS Res. Hum. Retrovir. 2020, 36, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Sathyamala, C. Depot contraception and HIV: An exercise in obfuscation. BMJ 2019, 367, l5768. [Google Scholar] [CrossRef] [PubMed]
- Marx, P.A.; Spira, A.I.; Gettie, A.; Dailey, P.J.; Veazey, R.S.; Lackner, A.A.; Mahoney, C.J.; Miller, C.J.; Claypool, L.E.; Ho, D.D.; et al. Progesterone implants enhance SIV vaginal transmission and early virus load. Nat. Med. 1996, 2, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Vishwanathan, S.A.; Guenthner, P.C.; Lin, C.Y.; Dobard, C.; Sharma, S.; Adams, D.R.; Otten, R.A.; Heneine, W.; Hendry, R.M.; McNicholl, J.M.; et al. High susceptibility to repeated, low-dose, vaginal SHIV exposure late in the luteal phase of the menstrual cycle of pigtail macaques. J. Acquir. Immune Defic. Syndr. 2011, 57, 261–264. [Google Scholar] [CrossRef]
- Zalenskaya, I.A.; Chandra, N.; Yousefieh, N.; Fang, X.; Adedipe, O.E.; Jackson, S.S.; Anderson, S.M.; Mauck, C.K.; Schwartz, J.L.; Thurman, A.R.; et al. Use of contraceptive depot medroxyprogesterone acetate is associated with impaired cervicovaginal mucosal integrity. J. Clin. Investig. 2018, 128, 4622–4638. [Google Scholar] [CrossRef]
- Chandra, N.; Thurman, A.R.; Anderson, S.; Cunningham, T.D.; Yousefieh, N.; Mauck, C.; Doncel, G.F. Depot medroxyprogesterone acetate increases immune cell numbers and activation markers in human vaginal mucosal tissues. AIDS Res. Hum. Retrovir. 2013, 29, 592–601. [Google Scholar] [CrossRef]
- Poonia, B.; Walter, L.; Dufour, J.; Harrison, R.; Marx, P.A.; Veazey, R.S. Cyclic changes in the vaginal epithelium of normal rhesus macaques. J. Endocrinol. 2006, 190, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Hild-Petito, S.; Veazey, R.S.; Larner, J.M.; Reel, J.R.; Blye, R.P. Effects of two progestin-only contraceptives, Depo-Provera and Norplant-II, on the vaginal epithelium of rhesus monkeys. AIDS Res. Hum. Retrovir. 1998, 14 (Suppl. 1), S125–S130. [Google Scholar]
- Edfeldt, G.; Lajoie, J.; Röhl, M.; Oyugi, J.; Åhlberg, A.; Khalilzadeh-Binicy, B.; Bradley, F.; Mack, M.; Kimani, J.; Omollo, K.; et al. Regular use of depot medroxyprogesterone acetate causes thinning of the superficial lining and apical distribution of HIV target cells in the human ectocervix. J. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Byrne, E.H.; Anahtar, M.N.; Cohen, K.E.; Moodley, A.; Padavattan, N.; Ismail, N.; Bowman, B.A.; Olson, G.S.; Mabhula, A.; Leslie, A.; et al. Association between injectable progestin-only contraceptives and HIV acquisition and HIV target cell frequency in the female genital tract in South African women: A prospective cohort study. Lancet Infect. Dis. 2016, 16, 441–448. [Google Scholar] [CrossRef]
- Guthrie, B.L.; Introini, A.; Roxby, A.C.; Choi, R.Y.; Bosire, R.; Lohman-Payne, B.; Hirbod, T.; Farquhar, C.; Broliden, K. Depot Medroxyprogesterone Acetate Use Is Associated With Elevated Innate Immune Effector Molecules in Cervicovaginal Secretions of HIV-1-Uninfected Women. J. Acquir. Immune Defic. Syndr. 2015, 69, 1–10. [Google Scholar] [CrossRef]
- Deese, J.; Masson, L.; Miller, W.; Cohen, M.; Morrison, C.; Wang, M.; Ahmed, K.; Agot, K.; Crucitti, T.; Abdellati, S.; et al. Injectable Progestin-Only Contraception is Associated With Increased Levels of Pro-Inflammatory Cytokines in the Female Genital Tract. Am. J. Reprod. Immunol. 2015, 74, 357–367. [Google Scholar] [CrossRef]
- Morrison, C.; Fichorova, R.N.; Mauck, C.; Chen, P.L.; Kwok, C.; Chipato, T.; Salata, R.; Doncel, G.F. Cervical inflammation and immunity associated with hormonal contraception, pregnancy, and HIV-1 seroconversion. J. Acquir. Immune Defic. Syndr. 2014, 66, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, K.G.; Shah, N.; Klein, R.S.; Mayer, K.H.; Sobel, J.D.; Warren, D.L.; Jamieson, D.J.; Duerr, A.C.; Rompalo, A.M. Group HIVERS: Influence of sex hormones, HIV status, and concomitant sexually transmitted infection on cervicovaginal inflammation. J. Infect. Dis. 2005, 191, 358–366. [Google Scholar] [CrossRef]
- Wieser, F.; Hosmann, J.; Tschugguel, W.; Czerwenka, K.; Sedivy, R.; Huber, J.C. Progesterone increases the number of Langerhans cells in human vaginal epithelium. Fertil. Steril. 2001, 75, 1234–1235. [Google Scholar] [CrossRef]
- Huijbregts, R.P.; Helton, E.S.; Michel, K.G.; Sabbaj, S.; Richter, H.E.; Goepfert, P.A.; Hel, Z. Hormonal contraception and HIV-1 infection: Medroxyprogesterone acetate suppresses innate and adaptive immune mechanisms. Endocrinology 2013, 154, 1282–1295. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Trunova, N.; Tsai, L.; Tung, S.; Schneider, E.; Harouse, J.; Gettie, A.; Simon, V.; Blanchard, J.; Cheng-Mayer, C. Progestin-based contraceptive suppresses cellular immune responses in SHIV-infected rhesus macaques. Virology 2006, 352, 169–177. [Google Scholar] [CrossRef]
- Mitchell, C.M.; McLemore, L.; Westerberg, K.; Astronomo, R.; Smythe, K.; Gardella, C.; Mack, M.; Magaret, A.; Patton, D.; Agnew, K.; et al. Long-term effect of depot medroxyprogesterone acetate on vaginal microbiota, epithelial thickness and HIV target cells. J. Infect. Dis. 2014, 210, 651–655. [Google Scholar] [CrossRef]
- Ferreira, V.H.; Dizzell, S.; Nazli, A.; Kafka, J.K.; Mueller, K.; Nguyen, P.V.; Tremblay, M.J.; Cochrane, A.; Kaushic, C. Medroxyprogesterone Acetate Regulates HIV-1 Uptake and Transcytosis but Not Replication in Primary Genital Epithelial Cells, Resulting in Enhanced T-Cell Infection. J. Infect. Dis. 2015, 211, 1745–1756. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Gach, J.S.; Becerra, J.C.; Phan, T.B.; Pudney, J.; Moldoveanu, Z.; Joseph, S.B.; Landucci, G.; Supnet, M.J.; Ping, L.H.; et al. The Neonatal Fc Receptor (FcRn) Enhances Human Immunodeficiency Virus Type 1 (HIV-1) Transcytosis across Epithelial Cells. PLoS Pathog. 2013, 9, e1003776. [Google Scholar] [CrossRef]
- Bomsel, M. Transcytosis of infectious human immunodeficiency virus across a tight human epithelial cell line barrier. Nat. Med. 1997, 3, 42–47. [Google Scholar] [CrossRef]
- Bobardt, M.D.; Chatterji, U.; Selvarajah, S.; van der Schueren, B.; David, G.; Kahn, B.; Gallay, P.A. Cell-free human immunodeficiency virus type 1 transcytosis through primary genital epithelial cells. J. Virol. 2007, 81, 395–405. [Google Scholar] [CrossRef]
- Devito, C.; Broliden, K.; Kaul, R.; Svensson, L.; Johansen, K.; Kiama, P.; Kimani, J.; Lopalco, L.; Piconi, S.; Bwayo, J.J.; et al. Mucosal and plasma IgA from HIV-1-exposed uninfected individuals inhibit HIV-1 transcytosis across human epithelial cells. J. Immunol. 2000, 165, 5170–5176. [Google Scholar] [CrossRef] [PubMed]
- Saidi, H.; Magri, G.; Nasreddine, N.; Requena, M.; Belec, L. R5- and X4-HIV-1 use differentially the endometrial epithelial cells HEC-1A to ensure their own spread: Implication for mechanisms of sexual transmission. Virology 2007, 358, 55–68. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Maher, D.; Wu, X.; Schacker, T.; Horbul, J.; Southern, P. HIV binding, penetration, and primary infection in human cervicovaginal tissue. Proc. Natl. Acad. Sci. USA 2005, 102, 11504–11509. [Google Scholar] [CrossRef] [PubMed]
- Carias, A.M.; McCoombe, S.; McRaven, M.; Anderson, M.; Galloway, N.; Vandergrift, N.; Fought, A.J.; Lurain, J.; Duplantis, M.; Veazey, R.S.; et al. Defining the interaction of HIV-1 with the mucosal barriers of the female reproductive tract. J. Virol. 2013, 87, 11388–11400. [Google Scholar] [CrossRef]
- Kinlock, B.L.; Wang, Y.; Turner, T.M.; Wang, C.; Liu, B. Transcytosis of HIV-1 through vaginal epithelial cells is dependent on trafficking to the endocytic recycling pathway. PLoS ONE 2014, 9, e96760. [Google Scholar] [CrossRef]
- Halpern, V.; Combes, S.L.; Dorflinger, L.J.; Weiner, D.H.; Archer, D.F. Pharmacokinetics of subcutaneous depot medroxyprogesterone acetate injected in the upper arm. Contraception 2014, 89, 31–35. [Google Scholar] [CrossRef]
- Ortiz, A.; Hirol, M.; Stanczyk, F.Z.; Goebelsmann, U.; Mishell, D.R. Serum medroxyprogesterone acetate (MPA) concentrations and ovarian function following intramuscular injection of depo-MPA. J. Clin. Endocrinol. Metab. 1977, 44, 32–38. [Google Scholar] [CrossRef]
- Manuals & Protocols: Lysotracker and Lysosensor Probes. Available online: https://assets.thermofisher.com/TFS-Assets/LSG/manuals/mp07525.pdf (accessed on 16 August 2021).
- Tanida, I.; Minematsu-Ikeguchi, N.; Ueno, T.; Kominami, E. Lysosomal turnover, but not a cellular level, of endogenous LC3 is a marker for autophagy. Autophagy 2005, 1, 84–91. [Google Scholar] [CrossRef]
- Tanida, I.; Yamaji, T.; Ueno, T.; Ishiura, S.; Kominami, E.; Hanada, K. Consideration about negative controls for LC3 and expression vectors for four colored fluorescent protein-LC3 negative controls. Autophagy 2008, 4, 131–134. [Google Scholar] [CrossRef]
- Joseph, S.B.; Swanstrom, R.; Kashuba, A.D.; Cohen, M.S. Bottlenecks in HIV-1 transmission: Insights from the study of founder viruses. Nat. Rev. Microbiol. 2015, 13, 414–425. [Google Scholar] [CrossRef]
- Helen, R.; Consortium, E. DMPA and HIV: Why we need a trial. Contraception 2014, 90, 354–356. [Google Scholar]
- Gollub, E.L.; Jones, H.E.; Ralph, L.J.; van de Wijgert, J.; Padian, N.; Stein, Z. The Need for Policy Change Regarding Progestin-Only Injectable Contraceptives. J. Women’s Health 2019, 28, 1180–1184. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.E. Time to focus on improving the contraceptive method mix in high HIV prevalence settings and let go of unanswerable questions. Contraception 2014, 90, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, E.; Soros, V.B.; Greene, W.C. Compensatory link between fusion and endocytosis of human immunodeficiency virus type 1 in human CD4 T lymphocytes. J. Virol. 2004, 78, 1375–1383. [Google Scholar] [CrossRef]
- Chikhlikar, P.; de Barros Arruda, L.; Maciel, M.; Silvera, P.; Lewis, M.G.; August, J.T.; Marques, E.T. DNA encoding an HIV-1 Gag/human lysosome-associated membrane protein-1 chimera elicits a broad cellular and humoral immune response in Rhesus macaques. PLoS ONE 2006, 1, e135. [Google Scholar] [CrossRef]
- Starodubova, E.S.; Isaguliants, M.G.; Kuzmenko, Y.V.; Latanova, A.A.; Krotova, O.A.; Karpov, V.L. Fusion to the Lysosome Targeting Signal of the Invariant Chain Alters the Processing and Enhances the Immunogenicity of HIV-1 Reverse Transcriptase. Acta Nat. 2014, 6, 61–68. [Google Scholar] [CrossRef]
- Kubo, Y.; Izumida, M.; Yashima, Y.; Yoshii-Kamiyama, H.; Tanaka, Y.; Yasui, K.; Hayashi, H.; Matsuyama, T. Gamma-interferon-inducible, lysosome/endosome-localized thiolreductase, GILT, has anti-retroviral activity and its expression is counteracted by HIV-1. Oncotarget 2016, 7, 71255–71273. [Google Scholar] [CrossRef]
- Shi, J.; Xiong, R.; Zhou, T.; Su, P.; Zhang, X.; Qiu, X.; Li, H.; Li, S.; Yu, C.; Wang, B.; et al. HIV-1 Nef Antagonizes SERINC5 Restriction by Downregulation of SERINC5 via the Endosome/Lysosome System. J. Virol. 2018, 92, 11. [Google Scholar] [CrossRef]
- Marques, E.T., Jr.; Chikhlikar, P.; de Arruda, L.B.; Leao, I.C.; Lu, Y.; Wong, J.; Chen, J.S.; Byrne, B.; August, J.T. HIV-1 p55Gag encoded in the lysosome-associated membrane protein-1 as a DNA plasmid vaccine chimera is highly expressed, traffics to the major histocompatibility class II compartment, and elicits enhanced immune responses. J. Biol. Chem. 2003, 278, 37926–37936. [Google Scholar] [CrossRef]
- Fields, J.; Dumaop, W.; Eleuteri, S.; Campos, S.; Serger, E.; Trejo, M.; Kosberg, K.; Adame, A.; Spencer, B.; Rockenstein, E.; et al. HIV-1 Tat alters neuronal autophagy by modulating autophagosome fusion to the lysosome: Implications for HIV-associated neurocognitive disorders. J. Neurosci. 2015, 35, 1921–1938. [Google Scholar] [CrossRef]
- Santerre, M.; Arjona, S.P.; Allen, C.N.; Callen, S.; Buch, S.; Sawaya, B.E. HIV-1 Vpr protein impairs lysosome clearance causing SNCA/alpha-synuclein accumulation in neurons. Autophagy 2021, 17, 1768–1782. [Google Scholar] [CrossRef]
- Moorjani, H.; Craddock, B.P.; Morrison, S.A.; Steigbigel, R.T. Impairment of phagosome-lysosome fusion in HIV-1-infected macrophages. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1996, 13, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Fredericksen, B.L.; Wei, B.L.; Yao, J.; Luo, T.; Garcia, J.V. Inhibition of endosomal/lysosomal degradation increases the infectivity of human immunodeficiency virus. J. Virol. 2002, 76, 11440–11446. [Google Scholar] [CrossRef]
- Wei, B.L.; Denton, P.W.; O′Neill, E.; Luo, T.; Foster, J.L.; Garcia, J.V. Inhibition of lysosome and proteasome function enhances human immunodeficiency virus type 1 infection. J. Virol. 2005, 79, 5705–5712. [Google Scholar] [CrossRef]
- Rowell, J.F.; Ruff, A.L.; Guarnieri, F.G.; Staveley-O’ Carroll, K.; Lin, X.; Tang, J.; August, J.T.; Siliciano, R.F. Lysosome-associated membrane protein-1-mediated targeting of the HIV-1 envelope protein to an endosomal/lysosomal compartment enhances its presentation to MHC class II-restricted T cells. J. Immunol. 1995, 155, 1818–1828. [Google Scholar] [PubMed]
- Africander, D.; Louw, R.; Verhoog, N.; Noeth, D.; Hapgood, J.P. Differential regulation of endogenous pro-inflammatory cytokine genes by medroxyprogesterone acetate and norethisterone acetate in cell lines of the female genital tract. Contraception 2011, 84, 423–435. [Google Scholar] [CrossRef]
- Ratner, L.; Haseltine, W.; Patarca, R.; Livak, K.J.; Starcich, B.; Josephs, S.F.; Doran, E.R.; Rafalski, J.A.; Whitehorn, E.A.; Baumeister, K.; et al. Complete nucleotide sequence of the AIDS virus, HTLV-III. Nature 1985, 313, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Sarngadharan, M.G.; Read, E.; Gallo, R.C. Detection, isolation, and continuous production of cytopathic retroviruses (HTLV-III) from patients with AIDS and pre-AIDS. Science 1984, 224, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Read-Connole, E.; Gallo, R.C. T4 positive human neoplastic cell lines susceptible to and permissive for HTLV-III. Lancet 1984, 2, 1472–1473. [Google Scholar] [CrossRef]
- Keele, B.F.; Giorgi, E.E.; Salazar-Gonzalez, J.F.; Decker, J.M.; Pham, K.T.; Salazar, M.G.; Sun, C.; Grayson, T.; Wang, S.; Li, H.; et al. Identification and characterization of transmitted and early founder virus envelopes in primary HIV-1 infection. Proc. Natl. Acad. Sci. USA 2008, 105, 7552–7557. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Choe, J.S.; Seo, H.R.; Park, J.S. Effect of Poly (ethylene glycol) Grafting on Polyethylenimine as a Gene Transfer Vector in vitro. Bull. Korean Chem. Soc. 2001, 22, 46–52. [Google Scholar]
- Fichorova, R.N.; Rheinwald, J.G.; Anderson, D.J. Generation of papillomavirus-immortalized cell lines from normal human ectocervical, endocervical, and vaginal epithelium that maintain expression of tissue-specific differentiation proteins. Biol. Reprod. 1997, 57, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Wehrly, K.; Chesebro, B. P24 antigen capture assay for quantification of human immunodeficiency virus using readily available inexpensive reagents. Methods 1997, 12, 288–293. [Google Scholar] [CrossRef]
- Wang, Y.; Shao, Q.; Yu, X.; Kong, W.; Hildreth, J.E.; Liu, B. N-terminal hemagglutinin tag renders lysine-deficient APOBEC3G resistant to HIV-1 Vif-induced degradation by reduced polyubiquitination. J. Virol. 2011, 85, 4510–4519. [Google Scholar] [CrossRef][Green Version]
- Drosten, C.; Muller-Kunert, E.; Dietrich, M.; Gerdes, J.; Schmitz, H. Topographic and quantitative display of integrated human immunodeficiency virus-1 provirus DNA in human lymph nodes by real-time polymerase chain reaction. J. Mol. Diagn. 2005, 7, 219–225. [Google Scholar] [CrossRef][Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, X.; Shao, Q.; Chaudhry, A.R.; Kinlock, B.L.; Izban, M.G.; Zhang, H.-Y.; Villalta, F.; Hildreth, J.E.K.; Liu, B. Medroxyprogesterone Acetate (MPA) Enhances HIV-1 Accumulation and Release in Primary Cervical Epithelial Cells by Inhibiting Lysosomal Activity. Pathogens 2021, 10, 1192. https://doi.org/10.3390/pathogens10091192
Jia X, Shao Q, Chaudhry AR, Kinlock BL, Izban MG, Zhang H-Y, Villalta F, Hildreth JEK, Liu B. Medroxyprogesterone Acetate (MPA) Enhances HIV-1 Accumulation and Release in Primary Cervical Epithelial Cells by Inhibiting Lysosomal Activity. Pathogens. 2021; 10(9):1192. https://doi.org/10.3390/pathogens10091192
Chicago/Turabian StyleJia, Xiangxu, Qiujia Shao, Ahsen R. Chaudhry, Ballington L. Kinlock, Michael G. Izban, Hong-Ying Zhang, Fernando Villalta, James E. K. Hildreth, and Bindong Liu. 2021. "Medroxyprogesterone Acetate (MPA) Enhances HIV-1 Accumulation and Release in Primary Cervical Epithelial Cells by Inhibiting Lysosomal Activity" Pathogens 10, no. 9: 1192. https://doi.org/10.3390/pathogens10091192
APA StyleJia, X., Shao, Q., Chaudhry, A. R., Kinlock, B. L., Izban, M. G., Zhang, H.-Y., Villalta, F., Hildreth, J. E. K., & Liu, B. (2021). Medroxyprogesterone Acetate (MPA) Enhances HIV-1 Accumulation and Release in Primary Cervical Epithelial Cells by Inhibiting Lysosomal Activity. Pathogens, 10(9), 1192. https://doi.org/10.3390/pathogens10091192