
Oxidative Stress Response in Pseudomonas aeruginosa

 ,
,  ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Pseudomonas aeruginosa Pathogenicity and Virulence
2. Oxidative Stress
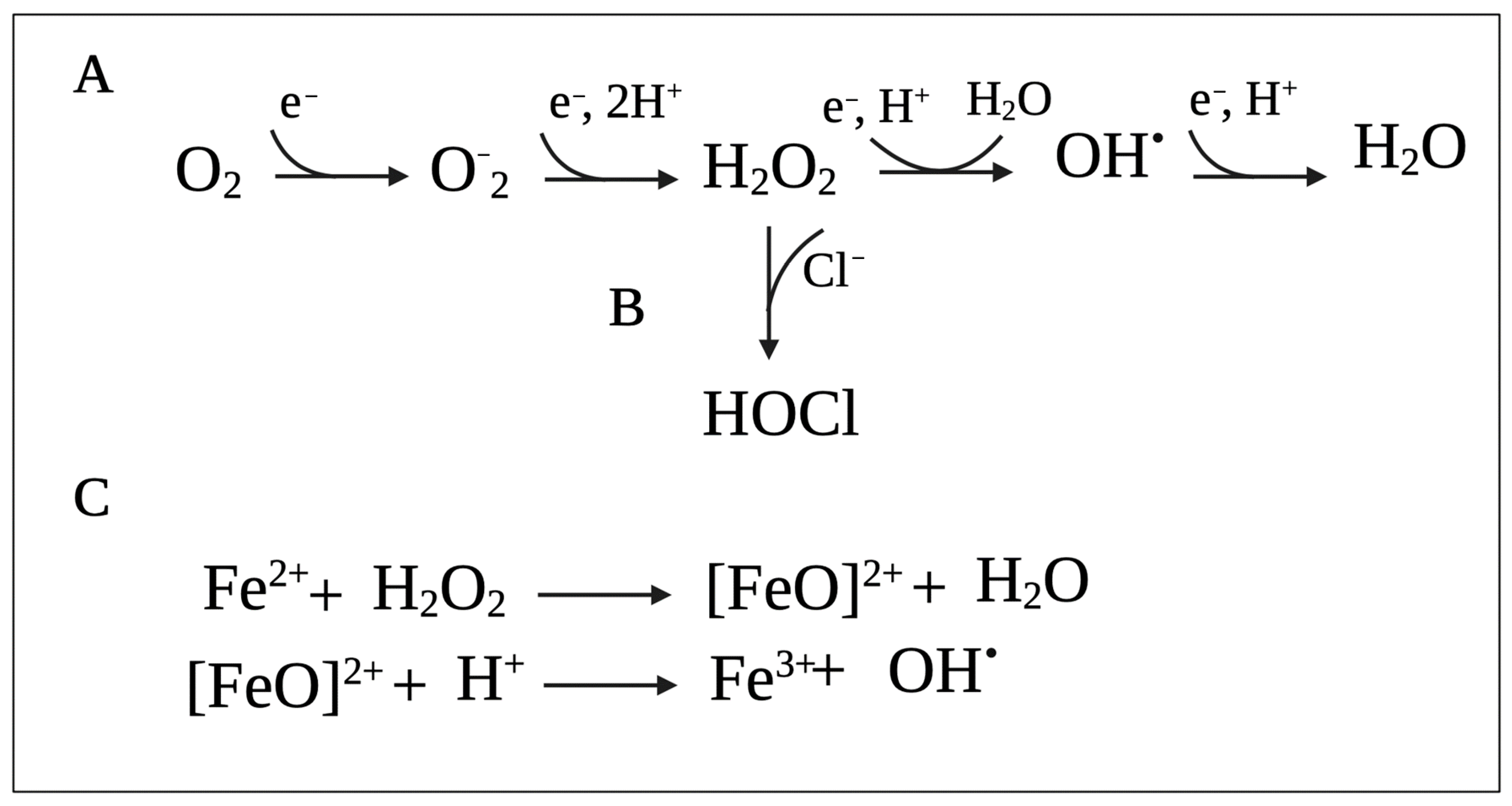
2.1. Reactive Oxygen Species
2.2. Reactive Chlorine Species
2.3. Oxidative Stress Generated by the Host Immune System
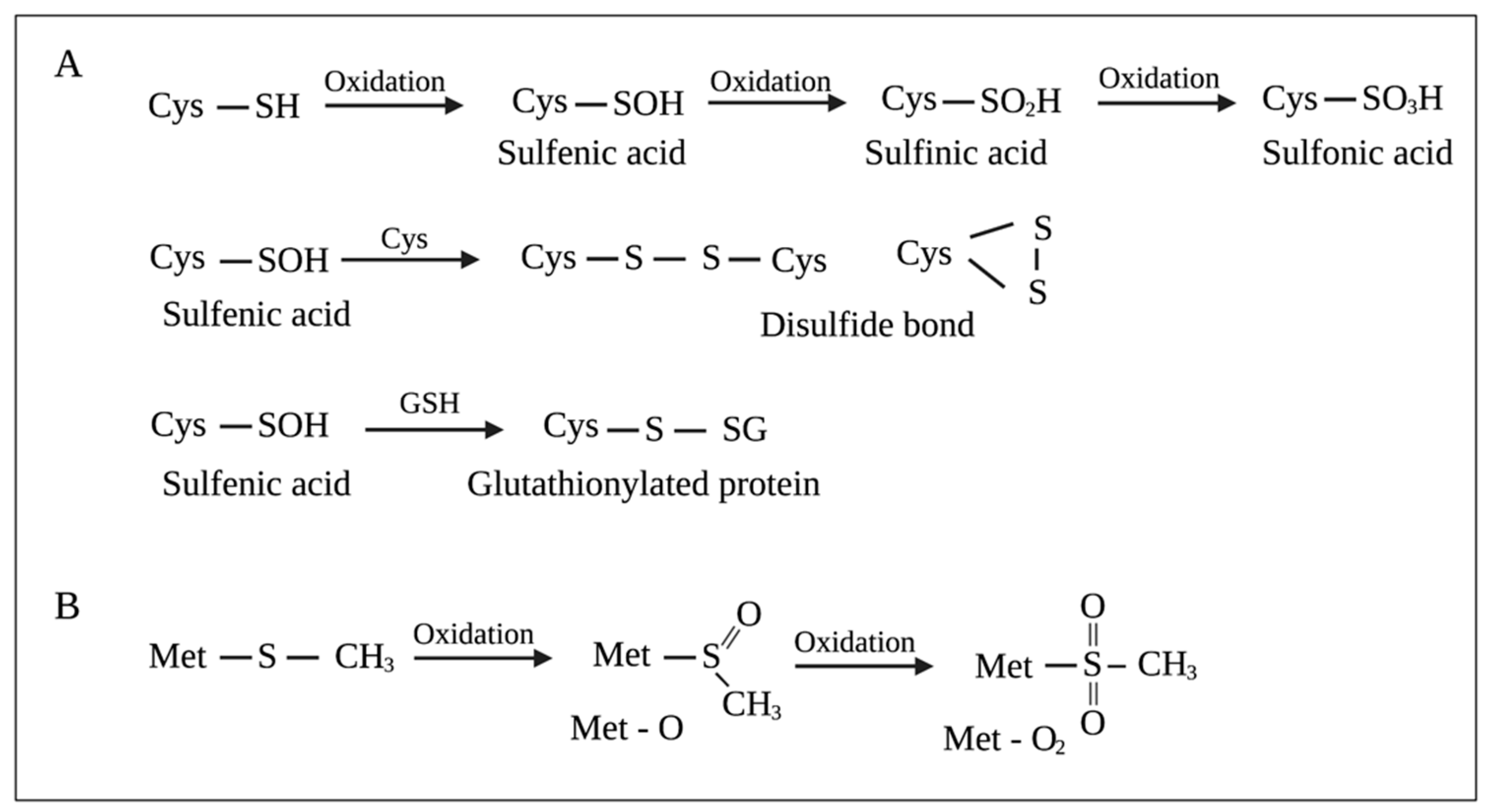
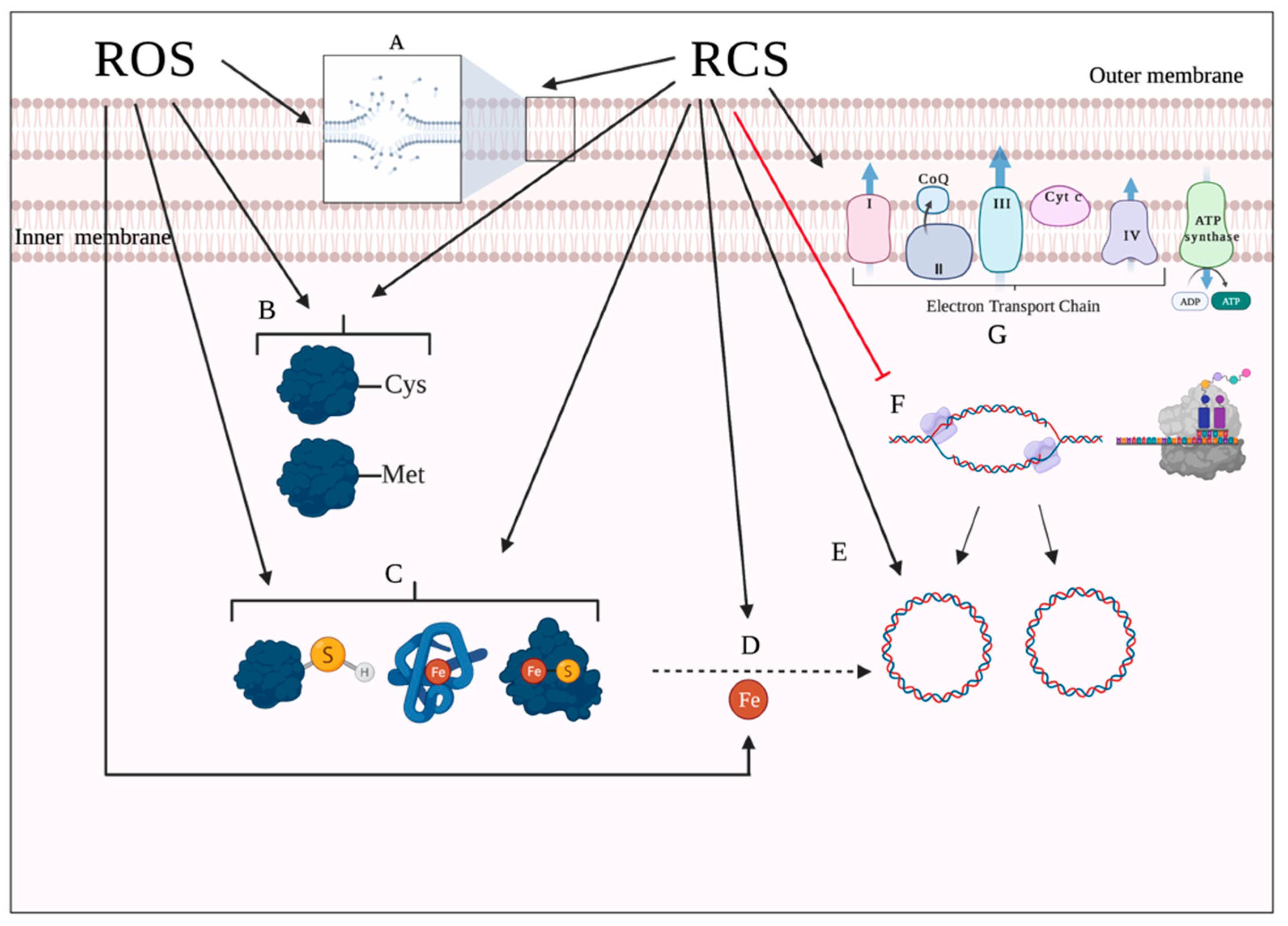
2.4. Mechanisms of Oxidative Cell Damage
3. Oxidative Stress Responses in P. aeruginosa
3.1. Evaluation of Oxidative Stress Responses of P. aeruginosa by Transcriptomic Studies
3.2. Cellular Responses to Oxidative Stress
3.2.1. Antioxidant Enzymes
3.2.2. Protein Repair Systems
3.2.3. Quorum Sensing (QS)
3.2.4. Induction of a Mucoid Phenotype
3.2.5. Pigment Production
3.2.6. Carbon Metabolism
3.2.7. Post-Transcriptional Modification
3.2.8. Other Stress Response Mechanisms
3.3. Transcriptional Regulators Involved in P. aeruginosa Oxidative Stress Responses
3.3.1. OxyR: The Master Regulator of Oxidative Stress
3.3.2. OhrR and OspR
3.3.3. IscR
3.3.4. Fur
3.3.5. SoxR
3.3.6. Other Transcriptional Regulators
3.4. Oxidative Stress Response Contributes to Antibiotic Resistance of P. aeruginosa
4. Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heimesaat, M.M.; Escher, U.; Grunau, A.; Kühl, A.A.; Bereswill, S. Multidrug-Resistant Pseudomonas aeruginosa Accelerate Intestinal, Extra-Intestinal, and Systemic Inflammatory Responses in Human Microbiota-Associated Mice With Subacute Ileitis. Front. Immunol. 2019, 10, 49. [Google Scholar] [CrossRef]
- Mohamed, F.A.; Shaker, G.H.; Askoura, M.M. Oxidative Stress Influences Pseudomonas aeruginosa Susceptibility to Antibiotics and Reduces Its Pathogenesis in Host. Curr. Microbiol. 2020, 77, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Sadikot, R.T.; Blackwell, T.S.; Christman, J.W.; Prince, A.S. Pathogen–Host Interactions in Pseudomonas aeruginosa Pneumonia. Am. J. Respir. Crit. Care Med. 2005, 171, 1209–1223. [Google Scholar] [CrossRef] [PubMed]
- Nixon, G.M.; Armstrong, D.S.; Carzino, R.; Carlin, J.B.; Olinsky, A.; Robertson, C.F.; Grimwood, K. Clinical Outcome after Early Pseudomonas aeruginosa Infection in Cystic Fibrosis. J. Pediatrics 2001, 138, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Harrison, F. Microbial Ecology of the Cystic Fibrosis Lung. Microbiology 2007, 153, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Parkins, M.D.; Somayaji, R.; Waters, V.J. Epidemiology, Biology, and Impact of Clonal Pseudomonas aeruginosa Infections in Cystic Fibrosis. Clin. Microbiol. Rev. 2018, 31, e00019-18. [Google Scholar] [CrossRef] [PubMed]
- Bassetti, M.; Vena, A.; Croxatto, A.; Righi, E.; Guery, B. How to Manage Pseudomonas aeruginosa Infections. Drugs Context 2018, 7, 1–18. [Google Scholar] [CrossRef]
- Horcajada, J.P.; Montero, M.; Oliver, A.; Sorlí, L.; Luque, S.; Gómez-Zorrilla, S.; Benito, N.; Grau, S. Epidemiology and Treatment of Multidrug-Resistant and Extensively Drug-Resistant Pseudomonas aeruginosa Infections. Clin. Microbiol. Rev. 2019, 32, e00031-19. [Google Scholar] [CrossRef]
- Diggle, S.P.; Whiteley, M. Microbe Profile: Pseudomonas aeruginosa: Opportunistic Pathogen and Lab Rat. Microbiology 2020, 166, 30–33. [Google Scholar] [CrossRef]
- Mao, G.; Song, Y.; Bartlam, M.; Wang, Y. Long-Term Effects of Residual Chlorine on Pseudomonas aeruginosa in Simulated Drinking Water Fed With Low AOC Medium. Front. Microbiol. 2018, 9, 879. [Google Scholar] [CrossRef]
- Breidenstein, E.B.M.; de la Fuente-Núñez, C.; Hancock, R.E.W. Pseudomonas aeruginosa: All Roads Lead to Resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef]
- Poole, K. Pseudomonas aeruginosa: Resistance to the Max. Front. Microbiol. 2011, 2, 65. [Google Scholar] [CrossRef] [PubMed]
- Breidenstein, E.B.M.; Janot, L.; Strehmel, J.; Fernandez, L.; Taylor, P.K.; Kukavica-Ibrulj, I.; Gellatly, S.L.; Levesque, R.C.; Overhage, J.; Hancock, R.E.W. The Lon Protease Is Essential for Full Virulence in Pseudomonas aeruginosa. PLoS ONE 2012, 7, e49123. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Jiang, T.; Zhang, S.; Yu, X. Exposure to Mutagenic Disinfection Byproducts Leads to Increase of Antibiotic Resistance in Pseudomonas aeruginosa. Environ. Sci. Technol. 2014, 48, 8188–8195. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic Resistance in Pseudomonas aeruginosa: Mechanisms and Alternative Therapeutic Strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Kolter, R. Flagellar and Twitching Motility Are Necessary for Pseudomonas aeruginosa Biofilm Development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef]
- Barken, K.B.; Pamp, S.J.; Yang, L.; Gjermansen, M.; Bertrand, J.J.; Klausen, M.; Givskov, M.; Whitchurch, C.B.; Engel, J.N.; Tolker-Nielsen, T. Roles of Type IV Pili, Flagellum-Mediated Motility and Extracellular DNA in the Formation of Mature Multicellular Structures in Pseudomonas aeruginosa Biofilms. Environ. Microbiol. 2008, 10, 2331–2343. [Google Scholar] [CrossRef]
- Persat, A.; Inclan, Y.F.; Engel, J.N.; Stone, H.A.; Gitai, Z. Type IV Pili Mechanochemically Regulate Virulence Factors in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2015, 112, 7563–7568. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.R. The Type III Secretion System of Pseudomonas aeruginosa: Infection by Injection. Nat. Rev. Microbiol. 2009, 7, 654–665. [Google Scholar] [CrossRef]
- Williams, B.J.; Dehnbostel, J.; Blackwell, T.S. Pseudomonas aeruginosa: Host Defence in Lung Diseases. Respirology 2010, 15, 1037–1056. [Google Scholar] [CrossRef]
- Blier, A.-S.; Vieillard, J.; Gerault, E.; Dagorn, A.; Varacavoudin, T.; Le Derf, F.; Orange, N.; Feuilloley, M.; Lesouhaitier, O. Quantification of Pseudomonas aeruginosa Hydrogen Cyanide Production by a Polarographic Approach. J. Microbiol. Methods 2012, 90, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Gellatly, S.L.; Hancock, R.E.W. Pseudomonas aeruginosa: New Insights into Pathogenesis and Host Defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Pritt, B.; O’Brien, L.; Winn, W. Mucoid Pseudomonas in Cystic Fibrosis. Am. J. Clin. Pathol. 2007, 128, 32–34. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Anraku, M.; Nakagawa, S.; Ojima, T. Discovery of a Novel Alginate Lyase from Nitratiruptor Sp. SB155-2 Thriving at Deep-Sea Hydrothermal Vents and Identification of the Residues Responsible for Its Heat Stability. J. Biol. Chem. 2016, 291, 15551–15563. [Google Scholar] [CrossRef] [PubMed]
- King, J.D.; Kocíncová, D.; Westman, E.L.; Lam, J.S. Review: Lipopolysaccharide Biosynthesis in Pseudomonas aeruginosa. Innate Immun. 2009, 15, 261–312. [Google Scholar] [CrossRef]
- Hall, S.; McDermott, C.; Anoopkumar-Dukie, S.; McFarland, A.J.; Forbes, A.; Perkins, A.V.; Davey, A.K.; Chess-Williams, R.; Kiefel, M.J.; Arora, D.; et al. Cellular Effects of Pyocyanin, a Secreted Virulence Factor of Pseudomonas aeruginosa. Toxins 2016, 8, 236. [Google Scholar] [CrossRef]
- Kim, D.; Sitepu, I.R.; Hashidoko, Y. Induction of Biofilm Formation in the Betaproteobacterium Burkholderia Unamae CK43B Exposed to Exogenous Indole and Gallic Acid. Appl. Environ. Microbiol. 2013, 79, 4845–4852. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, K.K. What Drives Bacteria to Produce a Biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef]
- Hoffman, L.R.; D’Argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside Antibiotics Induce Bacterial Biofilm Formation. Nature 2005, 436, 1171–1175. [Google Scholar] [CrossRef]
- Linares, J.F.; Gustafsson, I.; Baquero, F.; Martinez, J.L. Antibiotics as Intermicrobial Signaling Agents Instead of Weapons. Proc. Natl. Acad. Sci. USA 2006, 103, 19484–19489. [Google Scholar] [CrossRef]
- Bagge, N.; Schuster, M.; Hentzer, M.; Ciofu, O.; Givskov, M.; Greenberg, E.P.; Høiby, N. Pseudomonas aeruginosa Biofilms Exposed to Imipenem Exhibit Changes in Global Gene Expression and Beta-Lactamase and Alginate Production. Antimicrob. Agents Chemother. 2004, 48, 1175–1187. [Google Scholar] [CrossRef]
- Da Cruz Nizer, W.S.; Inkovskiy, V.; Overhage, J. Surviving Reactive Chlorine Stress: Responses of Gram-Negative Bacteria to Hypochlorous Acid. Microorganisms 2020, 8, 1220. [Google Scholar] [CrossRef]
- Klebensberger, J.; Birkenmaier, A.; Geffers, R.; Kjelleberg, S.; Philipp, B. SiaA and SiaD Are Essential for Inducing Autoaggregation as a Specific Response to Detergent Stress in Pseudomonas aeruginosa. Environ. Microbiol. 2009, 11, 3073–3086. [Google Scholar] [CrossRef]
- Kaplan, J.B. Antibiotic-Induced Biofilm Formation. Int. J. Artif. Organs 2011, 34, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Odenholt, I. Pharmacodynamic Effects of Subinhibitory Antibiotic Concentrations. Int. J. Antimicrob. Agents 2001, 17, 1–8. [Google Scholar] [CrossRef]
- Langendonk, R.F.; Neill, D.R.; Fothergill, J.L. The Building Blocks of Antimicrobial Resistance in Pseudomonas aeruginosa: Implications for Current Resistance-Breaking Therapies. Front. Cell. Infect. Microbiol. 2021, 11, 665759. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, M.; Linley, E.; Denyer, S.P.; McDonnell, G.; Simons, C.; Maillard, J.-Y. Mode of Action of Hydrogen Peroxide and Other Oxidizing Agents: Differences between Liquid and Gas Forms. J. Antimicrob. Chemother. 2010, 65, 2108–2115. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. Where in the World Do Bacteria Experience Oxidative Stress? Environ. Microbiol. 2019, 21, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, X. Contribution of Oxidative Damage to Antimicrobial Lethality. Antimicrob. Agents Chemother. 2009, 53, 1395–1402. [Google Scholar] [CrossRef]
- Imlay, J.A. The Molecular Mechanisms and Physiological Consequences of Oxidative Stress: Lessons from a Model Bacterium. Nat. Rev. Microbiol. 2013, 11, 443–454. [Google Scholar] [CrossRef]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.-F. Oxidative Stress, Protein Damage and Repair in Bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef]
- Imlay, J.A. A Metabolic Enzyme That Rapidly Produces Superoxide, Fumarate Reductase of Escherichia coli(∗). J. Biol. Chem. 1995, 270, 19767–19777. [Google Scholar] [CrossRef]
- Korshunov, S.; Imlay, J.A. Two Sources of Endogenous Hydrogen Peroxide in Escherichia coli. Mol. Microbiol. 2010, 75, 1389–1401. [Google Scholar] [CrossRef]
- Seaver, L.C.; Imlay, J.A. Are Respiratory Enzymes the Primary Sources of Intracellular Hydrogen Peroxide?*. J. Biol. Chem. 2004, 279, 48742–48750. [Google Scholar] [CrossRef]
- Storz, G.; Imlay, J.A. Oxidative Stress. Curr. Opin. Microbiol. 1999, 2, 188–194. [Google Scholar] [CrossRef]
- Dwyer, D.J.; Collins, J.J.; Walker, G.C. Unraveling the Physiological Complexities of Antibiotic Lethality. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 313–332. [Google Scholar] [CrossRef] [PubMed]
- Yeom, J.; Imlay, J.A.; Park, W. Iron Homeostasis Affects Antibiotic-Mediated Cell Death in Pseudomonas Species*. J. Biol. Chem. 2010, 285, 22689–22695. [Google Scholar] [CrossRef] [PubMed]
- Kohanski, M.A.; DePristo, M.A.; Collins, J.J. Sublethal Antibiotic Treatment Leads to Multidrug Resistance via Radical-Induced Mutagenesis. Mol. Cell 2010, 37, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Foti, J.J.; Devadoss, B.; Winkler, J.A.; Collins, J.J.; Walker, G.C. Oxidation of the Guanine Nucleotide Pool Underlies Cell Death by Bactericidal Antibiotics. Science 2012, 336, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Root, R.K.; Metcalf, J.; Oshino, N.; Chance, B. H2O2 Release from Human Granulocytes during Phagocytosis. I. Documentation, Quantitation, and Some Regulating Factors. J. Clin. Investig. 1975, 55, 945–955. [Google Scholar] [CrossRef]
- Babior, B.M.; Kipnes, R.S.; Curnutte, J.T. Biological Defense Mechanisms. The production by leukocytes of superoxide, a potential bactericidal agent. J. Clin. Investig. 1973, 52, 741–744. [Google Scholar] [CrossRef]
- Murdoch, L.E.; Bailey, L.; Banham, E.; Watson, F.; Adams, N.M.T.; Chewins, J. Evaluating Different Concentrations of Hydrogen Peroxide in an Automated Room Disinfection System. Lett. Appl. Microbiol. 2016, 63, 178–182. [Google Scholar] [CrossRef]
- Gray, M.J.; Wholey, W.-Y.; Jakob, U. Bacterial Responses to Reactive Chlorine Species. Annu. Rev. Microbiol. 2013, 67, 141–160. [Google Scholar] [CrossRef]
- Fukuzaki, S. Mechanisms of Actions of Sodium Hypochlorite in Cleaning and Disinfection Processes. Biocontrol Sci. 2006, 11, 147–157. [Google Scholar] [CrossRef]
- Rutala, W.A.; Weber, D.J. Uses of Inorganic Hypochlorite (Bleach) in Health-Care Facilities. Clin. Microbiol. Rev. 1997, 10, 597–610. [Google Scholar] [CrossRef]
- Sasai, M.; Yamamoto, M. Pathogen Recognition Receptors: Ligands and Signaling Pathways by Toll-Like Receptors. Int. Rev. Immunol. 2013, 32, 116–133. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kobayashi, K. Macrophages in Inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Amulic, B.; Cazalet, C.; Hayes, G.L.; Metzler, K.D.; Zychlinsky, A. Neutrophil Function: From Mechanisms to Disease. Annu. Rev. Immunol. 2012, 30, 459–489. [Google Scholar] [CrossRef] [PubMed]
- Underhill, D.M.; Ozinsky, A. Phagocytosis of Microbes: Complexity in Action. Annu. Rev. Immunol. 2002, 20, 825–852. [Google Scholar] [CrossRef] [PubMed]
- Flannagan, R.S.; Cosío, G.; Grinstein, S. Antimicrobial Mechanisms of Phagocytes and Bacterial Evasion Strategies. Nat. Rev. Microbiol. 2009, 7, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Rada, B. Interactions between Neutrophils and Pseudomonas aeruginosa in Cystic Fibrosis. Pathogens 2017, 6, 10. [Google Scholar] [CrossRef]
- Moser, C.; Jensen, P.Ø.; Thomsen, K.; Kolpen, M.; Rybtke, M.; Lauland, A.S.; Trøstrup, H.; Tolker-Nielsen, T. Immune Responses to Pseudomonas aeruginosa Biofilm Infections. Front. Immunol. 2021, 12, 237. [Google Scholar] [CrossRef]
- Versey, Z.; da Cruz Nizer, W.S.; Russell, E.; Zigic, S.; DeZeeuw, K.G.; Marek, J.E.; Overhage, J.; Cassol, E. Biofilm-Innate Immune Interface: Contribution to Chronic Wound Formation. Front. Immunol. 2021, 12, 648554. [Google Scholar] [CrossRef]
- Panday, A.; Sahoo, M.K.; Osorio, D.; Batra, S. NADPH Oxidases: An Overview from Structure to Innate Immunity-Associated Pathologies. Cell. Mol. Immunol. 2015, 12, 5–23. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Kettle, A.J.; Hampton, M.B. Reactive Oxygen Species and Neutrophil Function. Annu. Rev. Biochem. 2016, 85, 765–792. [Google Scholar] [CrossRef]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH Oxidase Activation and Bacterial Resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef] [PubMed]
- West, A.P.; Brodsky, I.E.; Rahner, C.; Woo, D.K.; Erdjument-Bromage, H.; Tempst, P.; Walsh, M.C.; Choi, Y.; Shadel, G.S.; Ghosh, S. TLR Signalling Augments Macrophage Bactericidal Activity through Mitochondrial ROS. Nature 2011, 472, 476–480. [Google Scholar] [CrossRef] [PubMed]
- West, A.P.; Shadel, G.S.; Ghosh, S. Mitochondria in Innate Immune Responses. Nat. Rev. Immunol. 2011, 11, 389–402. [Google Scholar] [CrossRef]
- Abuaita, B.H.; Schultz, T.L.; O’Riordan, M.X. Mitochondria-Derived Vesicles Deliver Antimicrobial Reactive Oxygen Species to Control Phagosome-Localized Staphylococcus aureus. Cell Host Microbe 2018, 24, 625–636.e5. [Google Scholar] [CrossRef] [PubMed]
- Painter, R.G.; Wang, G. Direct Measurement of Free Chloride Concentrations in the Phagolysosomes of Human Neutrophils. Anal. Chem. 2006, 78, 3133–3137. [Google Scholar] [CrossRef]
- Klebanoff, S.J.; Kettle, A.J.; Rosen, H.; Winterbourn, C.C.; Nauseef, W.M. Myeloperoxidase: A Front-Line Defender against Phagocytosed Microorganisms. J. Leukoc. Biol. 2013, 93, 185–198. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Hampton, M.B.; Livesey, J.H.; Kettle, A.J. Modeling the Reactions of Superoxide and Myeloperoxidase in the Neutrophil Phagosome: Implications for Microbial Killing. J. Biol. Chem. 2006, 281, 39860–39869. [Google Scholar] [CrossRef]
- Jiang, Q.; Griffin, D.A.; Barofsky, D.F.; Hurst, J.K. Intraphagosomal Chlorination Dynamics and Yields Determined Using Unique Fluorescent Bacterial Mimics. Chem. Res. Toxicol. 1997, 10, 1080–1089. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Kettle, A.J. Redox Reactions and Microbial Killing in the Neutrophil Phagosome. Antioxid. Redox Signal. 2013, 18, 642–660. [Google Scholar] [CrossRef]
- Weiss, S.J. Tissue Destruction by Neutrophils. N. Engl. J. Med. 1989, 320, 365–376. [Google Scholar] [CrossRef]
- Allen, R.C.; Stephens, J.T. Myeloperoxidase Selectively Binds and Selectively Kills Microbes. Infect. Immun. 2011, 79, 474–485. [Google Scholar] [CrossRef]
- Dukan, S.; Belkin, S.; Touati, D. Reactive Oxygen Species Are Partially Involved in the Bacteriocidal Action of Hypochlorous Acid. Arch. Biochem. Biophys. 1999, 367, 311–316. [Google Scholar] [CrossRef]
- Nyström, T. Role of Oxidative Carbonylation in Protein Quality Control and Senescence. EMBO J. 2005, 24, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Hampton, M.B. Thiol Chemistry and Specificity in Redox Signaling. Free Radic. Biol. Med. 2008, 45, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P. Kinetics and Mechanisms of Thiol-Disulfide Exchange Covering Direct Substitution and Thiol Oxidation-Mediated Pathways. Antioxid. Redox Signal. 2012, 18, 1623–1641. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. The Oxidative Environment and Protein Damage. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2005, 1703, 93–109. [Google Scholar] [CrossRef]
- Winter, J.; Ilbert, M.; Graf, P.C.F.; Ozcelik, D.; Jakob, U. Bleach Activates a Redox-Regulated Chaperone by Oxidative Protein Unfolding. Cell 2008, 135, 691–701. [Google Scholar] [CrossRef]
- Barrette, W.C.; Albrich, J.M.; Hurst, J.K. Hypochlorous Acid-Promoted Loss of Metabolic Energy in Escherichia coli. Infect. Immun. 1987, 55, 2518–2525. [Google Scholar] [CrossRef]
- Dukan, S.; Touati, D. Hypochlorous Acid Stress in Escherichia coli: Resistance, DNA Damage, and Comparison with Hydrogen Peroxide Stress. J. Bacteriol. 1996, 178, 6145–6150. [Google Scholar] [CrossRef]
- Albrich, J.M.; Gilbaugh, J.H.; Callahan, K.B.; Hurst, J.K. Effects of the Putative Neutrophil-Generated Toxin, Hypochlorous Acid, on Membrane Permeability and Transport Systems of Escherichia coli. J. Clin. Investig. 1986, 78, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Candeias, L.P.; Steenken, S. Electron Transfer in Di(Deoxy)Nucleoside Phosphates in Aqueous Solution: Rapid Migration of Oxidative Damage (via Adenine) to Guanine. J. Am. Chem. Soc. 1993, 115, 2437–2440. [Google Scholar] [CrossRef]
- Giese, B. Electron Transfer in DNA. Curr. Opin. Chem. Biol. 2002, 6, 612–618. [Google Scholar] [CrossRef]
- Hogg, M.; Wallace, S.S.; Doublié, S. Bumps in the Road: How Replicative DNA Polymerases See DNA Damage. Curr. Opin. Struct. Biol. 2005, 15, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Touati, D.; Jacques, M.; Tardat, B.; Bouchard, L.; Despied, S. Lethal Oxidative Damage and Mutagenesis Are Generated by Iron in Delta Fur Mutants of Escherichia coli: Protective Role of Superoxide Dismutase. J. Bacteriol. 1995, 177, 2305–2314. [Google Scholar] [CrossRef]
- Cornelis, P.; Wei, Q.; Andrews, S.C.; Vinckx, T. Iron Homeostasis and Management of Oxidative Stress Response in Bacteria. Metallomics 2011, 3, 540–549. [Google Scholar] [CrossRef]
- Hassett, D.J.; Sokol, P.A.; Howell, M.L.; Ma, J.-F.; Schweizer, H.T.; Ochsner, U.; Vasil, M.L. Ferric Uptake Regulator (Fur) Mutants of Pseudomonas aeruginosa Demonstrate Defective Siderophore-Mediated Iron Uptake, Altered Aerobic Growth, and Decreased Superoxide Dismutase and Catalase Activities. J. Bacteriol. 1996, 178, 8. [Google Scholar] [CrossRef]
- Keyer, K.; Imlay, J.A. Superoxide Accelerates DNA Damage by Elevating Free-Iron Levels. Proc. Natl. Acad. Sci. USA 1996, 93, 13635–13640. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Davies, M.J. Hypochlorite-Induced Damage to DNA, RNA, and Polynucleotides: Formation of Chloramines and Nitrogen-Centered Radicals. Chem. Res. Toxicol. 2002, 15, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Kohen, R.; Nyska, A. Invited Review: Oxidation of Biological Systems: Oxidative Stress Phenomena, Antioxidants, Redox Reactions, and Methods for Their Quantification. Toxicol. Pathol. 2002, 30, 620–650. [Google Scholar] [CrossRef] [PubMed]
- Girotti, A.W. Lipid Hydroperoxide Generation, Turnover, and Effector Action in Biological Systems. J. Lipid Res. 1998, 39, 1529–1542. [Google Scholar] [CrossRef]
- Yin, H.; Xu, L.; Porter, N.A. Free Radical Lipid Peroxidation: Mechanisms and Analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Geiger, O. Bacterial Membrane Lipids: Diversity in Structures and Pathways. FEMS Microbiol. Rev. 2016, 40, 133–159. [Google Scholar] [CrossRef] [PubMed]
- Marles-Wright, J.; Lewis, R.J. Stress Responses of Bacteria. Curr. Opin. Struct. Biol. 2007, 17, 755–760. [Google Scholar] [CrossRef]
- Balasubramanian, D.; Mathee, K. Comparative Transcriptome Analyses of Pseudomonas aeruginosa. Hum. Genom. 2009, 3, 349. [Google Scholar] [CrossRef]
- Nde, C.W.; Jang, H.-J.; Toghrol, F.; Bentley, W.E. Toxicogenomic Response of Pseudomonas aeruginosa to Ortho-Phenylphenol. BMC Genom. 2008, 9, 473. [Google Scholar] [CrossRef]
- Small, D.A.; Chang, W.; Toghrol, F.; Bentley, W.E. Toxicogenomic Analysis of Sodium Hypochlorite Antimicrobial Mechanisms in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2007, 74, 176–185. [Google Scholar] [CrossRef]
- Palma, M.; DeLuca, D.; Worgall, S.; Quadri, L.E.N. Transcriptome Analysis of the Response of Pseudomonas aeruginosa to Hydrogen Peroxide. J. Bacteriol. 2004, 186, 248–252. [Google Scholar] [CrossRef]
- Salunkhe, P.; Töpfer, T.; Buer, J.; Tümmler, B. Genome-Wide Transcriptional Profiling of the Steady-State Response of Pseudomonas aeruginosa to Hydrogen Peroxide. J. Bacteriol. 2005, 187, 2565–2572. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Small, D.A.; Toghrol, F.; Bentley, W.E. Microarray Analysis of Pseudomonas aeruginosa Reveals Induction of Pyocin Genes in Response to Hydrogen Peroxide. BMC Genom. 2005, 6, 115. [Google Scholar] [CrossRef] [PubMed]
- Groitl, B.; Dahl, J.-U.; Schroeder, J.W.; Jakob, U. Pseudomonas aeruginosa Defense Systems against Microbicidal Oxidants. Mol. Microbiol. 2017, 106, 335–350. [Google Scholar] [CrossRef]
- Chang, W.; Small, D.A.; Toghrol, F.; Bentley, W.E. Microarray Analysis of Toxicogenomic Effects of Peracetic Acid on Pseudomonas aeruginosa. Environ. Sci. Technol. 2005, 39, 5893–5899. [Google Scholar] [CrossRef]
- Small, D.A.; Chang, W.; Toghrol, F.; Bentley, W.E. Comparative Global Transcription Analysis of Sodium Hypochlorite, Peracetic Acid, and Hydrogen Peroxide on Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2007, 76, 1093–1105. [Google Scholar] [CrossRef]
- Kashmiri, Z.N.; Mankar, S.A. Free Radicals and Oxidative Stress in Bacteria. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 34–40. [Google Scholar]
- Farrant, K.V.; Spiga, L.; Davies, J.C.; Williams, H.D. Response of Pseudomonas aeruginosa to the Innate Immune System-Derived Oxidants Hypochlorous Acid and Hypothiocyanous Acid. J. Bacteriol. 2021, 203, 23. [Google Scholar]
- Romsang, A.; Duang-Nkern, J.; Leesukon, P.; Saninjuk, K.; Vattanaviboon, P.; Mongkolsuk, S. The Iron-Sulphur Cluster Biosynthesis Regulator IscR Contributes to Iron Homeostasis and Resistance to Oxidants in Pseudomonas aeruginosa. PLoS ONE 2014, 9, e86763. [Google Scholar] [CrossRef] [PubMed]
- Cavinato, L.; Genise, E.; Luly, F.R.; Di Domenico, E.G.; Del Porto, P.; Ascenzioni, F. Escaping the Phagocytic Oxidative Burst: The Role of SODB in the Survival of Pseudomonas aeruginosa Within Macrophages. Front. Microbiol. 2020, 11, 326. [Google Scholar] [CrossRef]
- Ghorbal, S.K.B.; Maalej, L.; Chourabi, K.; Khefacha, S.; Ouzari, H.-I.; Chatti, A. Antioxidant Defense Mechanisms in Pseudomonas aeruginosa: Role of Iron-Cofactored Superoxide Dismutase in Response to UV-C Radiations. Curr. Microbiol. 2016, 73, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Hassett, D.J.; Woodruff, W.A.; Wozniak, D.J.; Vasil, M.L.; Cohen, M.S.; Ohman, D.E. Cloning and Characterization of the Pseudomonas aeruginosa SodA and SodB Genes Encoding Manganese- and Iron-Cofactored Superoxide Dismutase: Demonstration of Increased Manganese Superoxide Dismutase Activity in Alginate-Producing Bacteria. J. Bacteriol. 1993, 175, 7658–7665. [Google Scholar] [CrossRef] [PubMed]
- Hassett, D.J.; Schweizer, H.P.; Ohman, D.E. Pseudomonas aeruginosa SodA and SodB Mutants Defective in Manganese- and Iron-Cofactored Superoxide Dismutase Activity Demonstrate the Importance of the Iron-Cofactored Form in Aerobic Metabolism. J. Bacteriol. 1995, 177, 6330–6337. [Google Scholar] [CrossRef] [PubMed]
- Iiyama, K.; Chieda, Y.; Lee, J.M.; Kusakabe, T.; Yasunaga-Aoki, C.; Shimizu, S. Effect of Superoxide Dismutase Gene Inactivation on Virulence of Pseudomonas aeruginosa PAO1 toward the Silkworm, Bombyx mori. Appl. Environ. Microbiol. 2007, 73, 1569–1575. [Google Scholar] [CrossRef]
- Su, S.; Panmanee, W.; Wilson, J.J.; Mahtani, H.K.; Li, Q.; VanderWielen, B.D.; Makris, T.M.; Rogers, M.; McDaniel, C.; Lipscomb, J.D.; et al. Catalase (KatA) Plays a Role in Protection against Anaerobic Nitric Oxide in Pseudomonas aeruginosa. PLoS ONE 2014, 9, e91813. [Google Scholar] [CrossRef]
- Heo, Y.-J.; Chung, I.-Y.; Cho, W.-J.; Lee, B.-Y.; Kim, J.-H.; Choi, K.-H.; Lee, J.-W.; Hassett, D.J.; Cho, Y.-H. The Major Catalase Gene (KatA) of Pseudomonas aeruginosa PA14 Is under Both Positive and Negative Control of the Global Transactivator OxyR in Response to Hydrogen Peroxide. J. Bacteriol. 2010, 192, 381–390. [Google Scholar] [CrossRef]
- Lee, J.-S.; Heo, Y.-J.; Lee, J.K.; Cho, Y.-H. KatA, the Major Catalase, Is Critical for Osmoprotection and Virulence in Pseudomonas aeruginosa PA14. Infect. Immun. 2005, 73, 4399–4403. [Google Scholar] [CrossRef] [PubMed]
- Elkins, J.G.; Hassett, D.J.; Stewart, P.S.; Schweizer, H.P.; McDermott, T.R. Protective Role of Catalase in Pseudomonas aeruginosa Biofilm Resistance to Hydrogen Peroxide. Appl. Environ. Microbiol. 1999, 65, 4594–4600. [Google Scholar] [CrossRef]
- Mossialos, D.; Tavankar, G.R.; Zlosnik, J.E.A.; Williams, H.D. Defects in a Quinol Oxidase Lead to Loss of KatC Catalase Activity in Pseudomonas aeruginosa: KatC Activity Is Temperature Dependent and It Requires an Intact Disulphide Bond Formation System. Biochem. Biophys. Res. Commun. 2006, 341, 697–702. [Google Scholar] [CrossRef]
- Orlandi, V.T.; Martegani, E.; Bolognese, F. Catalase A Is Involved in the Response to Photooxidative Stress in Pseudomonas aeruginosa. Photodiagn. Photodyn. Ther. 2018, 22, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S.; Roe, F.; Rayner, J.; Elkins, J.G.; Lewandowski, Z.; Ochsner, U.A.; Hassett, D.J. Effect of Catalase on Hydrogen Peroxide Penetration into Pseudomonas aeruginosa Biofilms. Appl. Environ. Microbiol. 2000, 66, 836–838. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.-H.; Choi, Y.-S.; Cho, Y.-H. Unusual Properties of Catalase A (KatA) of Pseudomonas aeruginosa PA14 Are Associated with Its Biofilm Peroxide Resistance. J. Bacteriol. 2008, 190, 2663–2670. [Google Scholar] [CrossRef]
- Hare, N.J.; Scott, N.E.; Shin, E.H.H.; Connolly, A.M.; Larsen, M.R.; Palmisano, G.; Cordwell, S.J. Proteomics of the Oxidative Stress Response Induced by Hydrogen Peroxide and Paraquat Reveals a Novel AhpC-like Protein in Pseudomonas aeruginosa. Proteomics 2011, 11, 3056–3069. [Google Scholar] [CrossRef]
- Ochsner, U.A.; Vasil, M.L.; Alsabbagh, E.; Parvatiyar, K.; Hassett, D.J. Role of the Pseudomonas aeruginosa OxyR-RecG Operon in Oxidative Stress Defense and DNA Repair: OxyR-Dependent Regulation of KatB-AnkB, AhpB, and AhpC-AhpF. J. Bacteriol. 2000, 182, 4533–4544. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T.; Lee, S.S.; Mondal, S.; Tripathi, B.N.; Kim, S.; Lee, K.W.; Hong, S.H.; Bai, H.-W.; Cho, J.-Y.; Chung, B.Y. Enhancement of the Chaperone Activity of Alkyl Hydroperoxide Reductase C from Pseudomonas aeruginosa PAO1 Resulting from a Point-Specific Mutation Confers Heat Tolerance in Escherichia coli. Mol. Cells 2016, 39, 594–602. [Google Scholar] [CrossRef]
- Hong, S.H.; Singh, S.; Tripathi, B.N.; Mondal, S.; Lee, S.; Jung, H.S.; Cho, C.; Kaur, S.; Kim, J.-H.; Lee, S.; et al. Functional Properties and the Oligomeric State of Alkyl Hydroperoxide Reductase Subunit F (AhpF) in Pseudomonas aeruginosa. Protoplasma 2020, 257, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. Cellular Defenses against Superoxide and Hydrogen Peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef] [PubMed]
- Romsang, A.; Atichartpongkul, S.; Trinachartvanit, W.; Vattanaviboon, P.; Mongkolsuk, S. Gene Expression and Physiological Role of Pseudomonas aeruginosa Methionine Sulfoxide Reductases during Oxidative Stress. J. Bacteriol. 2013, 195, 3299–3308. [Google Scholar] [CrossRef]
- Ezraty, B.; Aussel, L.; Barras, F. Methionine Sulfoxide Reductases in Prokaryotes. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2005, 1703, 221–229. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, K.; Baum, K. The Role of Methionine Sulfoxide Reductases in Oxidative Stress Tolerance and Virulence of Staphylococcus aureus and Other Bacteria. Antioxidants 2018, 7, 128. [Google Scholar] [CrossRef]
- Sultana, S.; Foti, A.; Dahl, J.-U. Bacterial Defense Systems against the Neutrophilic Oxidant Hypochlorous Acid. Infect. Immun. 2020, 88, e00964-19. [Google Scholar] [CrossRef]
- Pena, R.T.; Blasco, L.; Ambroa, A.; González-Pedrajo, B.; Fernández-García, L.; López, M.; Bleriot, I.; Bou, G.; García-Contreras, R.; Wood, T.K.; et al. Relationship Between Quorum Sensing and Secretion Systems. Front. Microbiol. 2019, 10, 1100. [Google Scholar] [CrossRef] [PubMed]
- Maura, D.; Hazan, R.; Kitao, T.; Ballok, A.E.; Rahme, L.G. Evidence for Direct Control of Virulence and Defense Gene Circuits by the Pseudomonas aeruginosa Quorum Sensing Regulator, MvfR. Sci. Rep. 2016, 6, 34083. [Google Scholar] [CrossRef]
- Balasubramanian, D.; Schneper, L.; Kumari, H.; Mathee, K. A Dynamic and Intricate Regulatory Network Determines Pseudomonas aeruginosa Virulence. Nucleic Acids Res. 2013, 41, 1–20. [Google Scholar] [CrossRef]
- Sabra, W.; Kim, E.-J.; Zeng, A.-P. Physiological Responses of Pseudomonas aeruginosa PAO1 to Oxidative Stress in Controlled Microaerobic and Aerobic Cultures. Microbiology 2002, 148, 3195–3202. [Google Scholar] [CrossRef]
- Pacheco, G.J.; Reis, R.S.; Fernandes, A.C.L.B.; da Rocha, S.L.G.; Pereira, M.D.; Perales, J.; Freire, D.M.G. Rhamnolipid Production: Effect of Oxidative Stress on Virulence Factors and Proteome of Pseudomonas aeruginosa PA1. Appl. Microbiol. Biotechnol. 2012, 95, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Malešević, M.; Di Lorenzo, F.; Filipić, B.; Stanisavljević, N.; Novović, K.; Senerovic, L.; Polović, N.; Molinaro, A.; Kojić, M.; Jovčić, B. Pseudomonas aeruginosa Quorum Sensing Inhibition by Clinical Isolate Delftia Tsuruhatensis 11304: Involvement of N -Octadecanoylhomoserine Lactones. Sci. Rep. 2019, 9, 16465. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Kong, J.; Dong, B.; Huang, H.; Wang, K.; Wu, L.; Hou, C.; Liang, Y.; Li, B.; Chen, Y. Baicalein Attenuates the Quorum Sensing-Controlled Virulence Factors of Pseudomonas aeruginosa and Relieves the Inflammatory Response in P. Aeruginosa-Infected Macrophages by Downregulating the MAPK and NFκB Signal-Transduction Pathways. Drug Des. Dev. Ther. 2016, 10, 183–203. [Google Scholar] [CrossRef]
- Kievit, T.R.D. Quorum Sensing in Pseudomonas aeruginosa Biofilms. Environ. Microbiol. 2009, 11, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Sakuragi, Y.; Kolter, R. Quorum-Sensing Regulation of the Biofilm Matrix Genes (Pel) of Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 5383–5386. [Google Scholar] [CrossRef] [PubMed]
- Hassett, D.J.; Ma, J.-F.; Elkins, J.G.; McDermott, T.R.; Ochsner, U.A.; West, S.E.H.; Huang, C.-T.; Fredericks, J.; Burnett, S.; Stewart, P.S.; et al. Quorum Sensing in Pseudomonas aeruginosa Controls Expression of Catalase and Superoxide Dismutase Genes and Mediates Biofilm Susceptibility to Hydrogen Peroxide. Mol. Microbiol. 1999, 34, 1082–1093. [Google Scholar] [CrossRef]
- Bollinger, N.; Hassett, D.J.; Iglewski, B.H.; Costerton, J.W.; McDermott, T.R. Gene Expression in Pseudomonas aeruginosa: Evidence of Iron Override Effects on Quorum Sensing and Biofilm-Specific Gene Regulation. J. Bacteriol. 2001, 183, 1990–1996. [Google Scholar] [CrossRef]
- García-Contreras, R.; Nuñez-López, L.; Jasso-Chávez, R.; Kwan, B.W.; Belmont, J.A.; Rangel-Vega, A.; Maeda, T.; Wood, T.K. Quorum Sensing Enhancement of the Stress Response Promotes Resistance to Quorum Quenching and Prevents Social Cheating. ISME J. 2015, 9, 115–125. [Google Scholar] [CrossRef]
- Kafle, P.; Amoh, A.N.; Reaves, J.M.; Suneby, E.G.; Tutunjian, K.A.; Tyson, R.L.; Schneider, T.L. Molecular Insights into the Impact of Oxidative Stress on the Quorum-Sensing Regulator Protein LasR. J. Biol. Chem. 2016, 291, 11776–11786. [Google Scholar] [CrossRef]
- Häussler, S.; Becker, T. The Pseudomonas Quinolone Signal (PQS) Balances Life and Death in Pseudomonas aeruginosa Populations. PLoS Pathog. 2008, 4, e1000166. [Google Scholar] [CrossRef] [PubMed]
- Rao, J.; Damron, F.H.; Basler, M.; Digiandomenico, A.; Sherman, N.E.; Fox, J.W.; Mekalanos, J.J.; Goldberg, J.B. Comparisons of Two Proteomic Analyses of Non-Mucoid and Mucoid Pseudomonas aeruginosa Clinical Isolates from a Cystic Fibrosis Patient. Front. Microbiol. 2011, 2, 162. [Google Scholar] [CrossRef] [PubMed]
- Pulcrano, G.; Iula, D.V.; Raia, V.; Rossano, F.; Catania, M.R. Different Mutations in MucA Gene of Pseudomonas aeruginosa Mucoid Strains in Cystic Fibrosis Patients and Their Effect on AlgU Gene Expression. New Microbiol. 2012, 35, 295–305. [Google Scholar]
- Qiu, D.; Eisinger, V.M.; Rowen, D.W.; Yu, H.D. Regulated Proteolysis Controls Mucoid Conversion in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2007, 104, 8107–8112. [Google Scholar] [CrossRef]
- Wood, L.F.; Leech, A.J.; Ohman, D.E. Cell Wall-Inhibitory Antibiotics Activate the Alginate Biosynthesis Operon in Pseudomonas aeruginosa: Roles of Σ22 (AlgT) and the AlgW and Prc Proteases. Mol. Microbiol. 2006, 62, 412–426. [Google Scholar] [CrossRef]
- Martin, D.W.; Schurr, M.J.; Yu, H.; Deretic, V. Analysis of Promoters Controlled by the Putative Sigma Factor AlgU Regulating Conversion to Mucoidy in Pseudomonas aeruginosa: Relationship to Sigma E and Stress Response. J. Bacteriol. 1994, 176, 6688–6696. [Google Scholar] [CrossRef]
- Li, S.; Lou, X.; Xu, Y.; Teng, X.; Liu, R.; Zhang, Q.; Wu, W.; Wang, Y.; Bartlam, M. Structural Basis for the Recognition of MucA by MucB and AlgU in Pseudomonas aeruginosa. FEBS J. 2019, 286, 4982–4994. [Google Scholar] [CrossRef]
- Damron, F.H.; Yu, H.D. Pseudomonas aeruginosa MucD Regulates the Alginate Pathway through Activation of MucA Degradation via MucP Proteolytic Activity. J. Bacteriol. 2011, 193, 286–291. [Google Scholar] [CrossRef]
- Mathee, K.; Ciofu, O.; Sternberg, C.; Lindum, P.W.; Campbell, J.I.A.; Jensen, P.; Johnsen, A.H.; Givskov, M.; Ohman, D.E.; Søren, M.; et al. Mucoid Conversion of Pseudomonas aeruginosa by Hydrogen Peroxide: A Mechanism for Virulence Activation in the Cystic Fibrosis Lung. Microbiology 1999, 145, 1349–1357. [Google Scholar] [CrossRef]
- Damron, F.H.; Napper, J.; Teter, M.A.; Yu, H.D. Lipotoxin F of Pseudomonas aeruginosa Is an AlgU-Dependent and Alginate-Independent Outer Membrane Protein Involved in Resistance to Oxidative Stress and Adhesion to A549 Human Lung Epithelia. Microbiology 2009, 155, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Crespo, A.; Pedraz, L.; Van Der Hofstadt, M.; Gomila, G.; Torrents, E. Regulation of Ribonucleotide Synthesis by the Pseudomonas aeruginosa Two-Component System AlgR in Response to Oxidative Stress. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Boucher, J.C.; Martinez-Salazar, J.; Schurr, M.J.; Mudd, M.H.; Yu, H.; Deretic, V. Two Distinct Loci Affecting Conversion to Mucoidy in Pseudomonas aeruginosa in Cystic Fibrosis Encode Homologs of the Serine Protease HtrA. J. Bacteriol. 1996, 178, 511–523. [Google Scholar] [CrossRef]
- Orlandi, V.T.; Bolognese, F.; Chiodaroli, L.; Tolker-Nielsen, T.; Barbieri, P. Pigments Influence the Tolerance of Pseudomonas aeruginosa PAO1 to Photodynamically Induced Oxidative Stress. Microbiology 2015, 161, 2298–2309. [Google Scholar] [CrossRef]
- Thippakorn, C.; Isarankura-Na-Ayudhya, C.; Pannengpetch, S.; Isarankura-Na-Ayudhya, P.; Schaduangrat, N.; Nantasenamat, C.; Prachayasittikul, V. Oxidative Responses and Defense Mechanism of Hyperpigmented P. aeruginosa as Characterized by Proteomics and Metabolomics. EXCLI J. 2018, 17, 544–562. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Renga Thavasi, T.; Jayalakshmi, S. Phenazine Pigments from Pseudomonas aeruginosa and Their Application as Antibacterial Agent and Food Colourants. Res. J. Microbiol. 2008, 3, 122–128. [Google Scholar] [CrossRef]
- Jayaseelan, S.; Ramaswamy, D.; Dharmaraj, S. Pyocyanin: Production, Applications, Challenges and New Insights. World J. Microbiol. Biotechnol. 2014, 30, 1159–1168. [Google Scholar] [CrossRef]
- Visca, P.; Imperi, F.; Lamont, I.L. Pyoverdine Siderophores: From Biogenesis to Biosignificance. Trends Microbiol. 2007, 15, 22–30. [Google Scholar] [CrossRef]
- Rodríguez-Rojas, A.; Mena, A.; Martín, S.; Borrell, N.; Oliver, A.; Blázquez, J. Inactivation of the HmgA Gene of Pseudomonas aeruginosa Leads to Pyomelanin Hyperproduction, Stress Resistance and Increased Persistence in Chronic Lung Infection. Microbiology 2009, 155, 1050–1057. [Google Scholar] [CrossRef]
- Meirelles, L.A.; Newman, D.K. Both Toxic and Beneficial Effects of Pyocyanin Contribute to the Lifecycle of Pseudomonas aeruginosa. Mol. Microbiol. 2018, 110, 995–1010. [Google Scholar] [CrossRef]
- Hassett, D.J.; Charniga, L.; Bean, K.; Ohman, D.E.; Cohen, M.S. Response of Pseudomonas aeruginosa to Pyocyanin: Mechanisms of Resistance, Antioxidant Defenses, and Demonstration of a Manganese-Cofactored Superoxide Dismutase. Infect. Immun. 1992, 60, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, N.; Sharma, P.; Capalash, N. Pyocyanin Stimulates Quorum Sensing-Mediated Tolerance to Oxidative Stress and Increases Persister Cell Populations in Acinetobacter Baumannii. Infect. Immun. 2014, 82, 3417–3425. [Google Scholar] [CrossRef]
- Dolan, S.K.; Kohlstedt, M.; Trigg, S.; Ramirez, P.V.; Kaminski, C.F.; Wittmann, C.; Welch, M. Contextual Flexibility in Pseudomonas aeruginosa Central Carbon Metabolism during Growth in Single Carbon Sources. mBio 2020, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.; Dohnt, K.; Tielen, P.; Jahn, D.; Becker, J.; Wittmann, C. Robustness and Plasticity of Metabolic Pathway Flux among Uropathogenic Isolates of Pseudomonas aeruginosa. PLoS ONE 2014, 9, e88368. [Google Scholar] [CrossRef] [PubMed]
- Chavarría, M.; Nikel, P.I.; Pérez-Pantoja, D.; de Lorenzo, V. The Entner–Doudoroff Pathway Empowers Pseudomonas Putida KT2440 with a High Tolerance to Oxidative Stress. Environ. Microbiol. 2013, 15, 1772–1785. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Jung, J.; Jang, I.-A.; Madsen, E.L.; Park, W. Role of Glyoxylate Shunt in Oxidative Stress Response. J. Biol. Chem. 2016, 291, 11928–11938. [Google Scholar] [CrossRef]
- Ha, S.; Shin, B.; Park, W. Lack of Glyoxylate Shunt Dysregulates Iron Homeostasis in Pseudomonas aeruginosa. Microbiology 2018, 164, 587–599. [Google Scholar] [CrossRef]
- Koedooder, C.; Guéneuguès, A.; Van Geersdaële, R.; Vergé, V.; Bouget, F.-Y.; Labreuche, Y.; Obernosterer, I.; Blain, S. The Role of the Glyoxylate Shunt in the Acclimation to Iron Limitation in Marine Heterotrophic Bacteria. Front. Mar. Sci. 2018, 5, 435. [Google Scholar] [CrossRef]
- Morita, T.; Kawamoto, H.; Mizota, T.; Inada, T.; Aiba, H. Enolase in the RNA Degradosome Plays a Crucial Role in the Rapid Decay of Glucose Transporter MRNA in the Response to Phosphosugar Stress in Escherichia coli. Mol. Microbiol. 2004, 54, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.; Chen, F.; Liu, Y.; Zhao, Q.; Chen, R.; Pan, X.; Liu, C.; Cheng, Z.; Jin, S.; Jin, Y.; et al. Pseudomonas aeruginosa Enolase Influences Bacterial Tolerance to Oxidative Stresses and Virulence. Front. Microbiol. 2016, 7, 1999. [Google Scholar] [CrossRef] [PubMed]
- Corona, F.; Martínez, J.L.; Nikel, P.I. The Global Regulator Crc Orchestrates the Metabolic Robustness Underlying Oxidative Stress Resistance in Pseudomonas aeruginosa. Environ. Microbiol. 2019, 21, 898–912. [Google Scholar] [CrossRef] [PubMed]
- Picard, F.; Dressaire, C.; Girbal, L.; Cocaign-Bousquet, M. Examination of Post-Transcriptional Regulations in Prokaryotes by Integrative Biology. Comptes Rendus Biol. 2009, 332, 958–973. [Google Scholar] [CrossRef]
- Thongdee, N.; Jaroensuk, J.; Atichartpongkul, S.; Chittrakanwong, J.; Chooyoung, K.; Srimahaeak, T.; Chaiyen, P.; Vattanaviboon, P.; Mongkolsuk, S.; Fuangthong, M. TrmB, a TRNA M7G46 Methyltransferase, Plays a Role in Hydrogen Peroxide Resistance and Positively Modulates the Translation of KatA and KatB MRNAs in Pseudomonas aeruginosa. Nucleic Acids Res. 2019, 47, 9271–9281. [Google Scholar] [CrossRef]
- Jaroensuk, J.; Atichartpongkul, S.; Chionh, Y.H.; Wong, Y.H.; Liew, C.W.; McBee, M.E.; Thongdee, N.; Prestwich, E.G.; DeMott, M.S.; Mongkolsuk, S.; et al. Methylation at Position 32 of TRNA Catalyzed by TrmJ Alters Oxidative Stress Response in Pseudomonas aeruginosa. Nucleic Acids Res. 2016, 44, 10834–10848. [Google Scholar] [CrossRef] [PubMed]
- Summer, S.; Smirnova, A.; Gabriele, A.; Toth, U.; Fasemore, A.M.; Förstner, K.U.; Kuhn, L.; Chicher, J.; Hammann, P.; Mitulović, G.; et al. YBEY Is an Essential Biogenesis Factor for Mitochondrial Ribosomes. Nucleic Acids Res. 2020, 48, 9762–9786. [Google Scholar] [CrossRef]
- Xia, Y.; Weng, Y.; Xu, C.; Wang, D.; Pan, X.; Tian, Z.; Xia, B.; Li, H.; Chen, R.; Liu, C.; et al. Endoribonuclease YbeY Is Essential for RNA Processing and Virulence in Pseudomonas aeruginosa. mBio 2020, 11. [Google Scholar] [CrossRef]
- Gómez-Lozano, M.; Marvig, R.L.; Tulstrup, M.V.; Molin, S. Expression of Antisense Small RNAs in Response to Stress in Pseudomonas aeruginosa. BMC Genom. 2014, 15, 783. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, B.; Gruber, J.D.; Zhang, Y.-M.; Davies, C. Acyl Carrier Protein 3 Is Involved in Oxidative Stress Response in Pseudomonas aeruginosa. Front. Microbiol. 2018, 9, 2244. [Google Scholar] [CrossRef] [PubMed]
- Koskenkorva-Frank, T.S.; Kallio, P.T. Induction of Pseudomonas aeruginosa Fhp and FhpR by Reactive Oxygen Species. Can. J. Microbiol. 2009, 55, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Goldová, J.; Ulrych, A.; Hercík, K.; Branny, P. A Eukaryotic-Type Signalling System of Pseudomonas aeruginosa Contributes to Oxidative Stress Resistance, Intracellular Survival and Virulence. BMC Genom. 2011, 12, 437. [Google Scholar] [CrossRef]
- Moyano, A.J.; Tobares, R.A.; Rizzi, Y.S.; Krapp, A.R.; Mondotte, J.A.; Bocco, J.L.; Saleh, M.-C.; Carrillo, N.; Smania, A.M. A Long-Chain Flavodoxin Protects Pseudomonas aeruginosa from Oxidative Stress and Host Bacterial Clearance. PLoS Genet. 2014, 10, e1004163. [Google Scholar] [CrossRef]
- Panmanee, W.; Charoenlap, N.; Atichartpongkul, S.; Mahavihakanont, A.; Whiteside, M.D.; Winsor, G.; Brinkman, F.S.L.; Mongkolsuk, S.; Hassett, D.J. The OxyR-Regulated PhnW Gene Encoding 2-Aminoethylphosphonate:Pyruvate Aminotransferase Helps Protect Pseudomonas aeruginosa from Tert-Butyl Hydroperoxide. PLoS ONE 2017, 12, e0189066. [Google Scholar] [CrossRef]
- Ciofu, O.; Mandsberg, L.F.; Wang, H.; Høiby, N. Phenotypes Selected during Chronic Lung Infection in Cystic Fibrosis Patients: Implications for the Treatment of Pseudomonas aeruginosa Biofilm Infections. FEMS Immunol. Med. Microbiol. 2012, 65, 215–225. [Google Scholar] [CrossRef]
- Fernández-Barat, L.; Ciofu, O.; Kragh, K.N.; Pressler, T.; Johansen, U.; Motos, A.; Torres, A.; Hoiby, N. Phenotypic Shift in Pseudomonas aeruginosa Populations from Cystic Fibrosis Lungs after 2-Week Antipseudomonal Treatment. J. Cyst. Fibros. 2017, 16, 222–229. [Google Scholar] [CrossRef]
- Ciofu, O.; Riis, B.; Pressler, T.; Poulsen, H.E.; Høiby, N. Occurrence of Hypermutable Pseudomonas Aeruginosa in Cystic Fibrosis Patients Is Associated with the Oxidative Stress Caused by Chronic Lung Inflammation. Antimicrob. Agents Chemother. 2005, 49, 2276–2282. [Google Scholar] [CrossRef]
- Colque, C.A.; Albarracín Orio, A.G.; Feliziani, S.; Marvig, R.L.; Tobares, A.R.; Johansen, H.K.; Molin, S.; Smania, A.M. Hypermutator Pseudomonas aeruginosa Exploits Multiple Genetic Pathways To Develop Multidrug Resistance during Long-Term Infections in the Airways of Cystic Fibrosis Patients. Antimicrob. Agents Chemother. 2020, 64, e02142-19. [Google Scholar] [CrossRef]
- Bae, H.-W.; Cho, Y.-H. Mutational Analysis of Pseudomonas aeruginosa OxyR to Define the Regions Required for Peroxide Resistance and Acute Virulence. Res. Microbiol. 2012, 163, 55–63. [Google Scholar] [CrossRef]
- Wei, Q.; Le Minh, P.N.; Dötsch, A.; Hildebrand, F.; Panmanee, W.; Elfarash, A.; Schulz, S.; Plaisance, S.; Charlier, D.; Hassett, D.; et al. Global Regulation of Gene Expression by OxyR in an Important Human Opportunistic Pathogen. Nucleic Acids Res. 2012, 40, 4320–4333. [Google Scholar] [CrossRef]
- Papp-Szabò, E.; Firtel, M.; Josephy, P.D. Comparison of the Sensitivities of Salmonella Typhimurium OxyR and KatG Mutants to Killing by Human Neutrophils. Infect. Immun. 1994, 62, 2662–2668. [Google Scholar] [CrossRef]
- Storz, G.; Tartaglia, L.A. OxyR: A Regulator of Antioxidant Genes. J. Nutr. 1992, 122, 627–630. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sun, M.; Cheng, Y.; Yang, R.; Wen, Y.; Chen, Z.; Li, J. OxyR Is a Key Regulator in Response to Oxidative Stress in Streptomyces Avermitilis. Microbiology 2016, 162, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, K.; Ohniwa, R.L.; Kim, J.; Maruyama, A.; Ohta, T.; Takeyasu, K. Bacterial Nucleoid Dynamics: Oxidative Stress Response in Staphylococcus aureus. Genes Cells 2006, 11, 409–423. [Google Scholar] [CrossRef] [PubMed]
- Dubbs, J.M.; Mongkolsuk, S. Peroxide-Sensing Transcriptional Regulators in Bacteria. J. Bacteriol. 2012, 194, 5495–5503. [Google Scholar] [CrossRef] [PubMed]
- Vinckx, T.; Wei, Q.; Matthijs, S.; Cornelis, P. The Pseudomonas aeruginosa Oxidative Stress Regulator OxyR Influences Production of Pyocyanin and Rhamnolipids: Protective Role of Pyocyanin. Microbiology 2010, 156, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Vinckx, T.; Matthijs, S.; Cornelis, P. Loss of the Oxidative Stress Regulator OxyR in Pseudomonas aeruginosa PAO1 Impairs Growth under Iron-Limited Conditions. FEMS Microbiol. Lett. 2008, 288, 258–265. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sun, M.; Lyu, M.; Wen, Y.; Song, Y.; Li, J.; Chen, Z. Organic Peroxide-Sensing Repressor OhrR Regulates Organic Hydroperoxide Stress Resistance and Avermectin Production in Streptomyces avermitilis. Front. Microbiol. 2018, 9, 1398. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-Y.; Shin, J.-H.; Roe, J.-H. Dual Role of OhrR as a Repressor and an Activator in Response to Organic Hydroperoxides in Streptomyces Coelicolor. J. Bacteriol. 2007, 189, 6284–6292. [Google Scholar] [CrossRef] [PubMed]
- Atichartpongkul, S.; Fuangthong, M.; Vattanaviboon, P.; Mongkolsuk, S. Analyses of the Regulatory Mechanism and Physiological Roles of Pseudomonas aeruginosa OhrR, a Transcription Regulator and a Sensor of Organic Hydroperoxides. J. Bacteriol. 2010, 192, 2093–2101. [Google Scholar] [CrossRef]
- Atichartpongkul, S.; Vattanaviboon, P.; Wisitkamol, R.; Jaroensuk, J.; Mongkolsuk, S.; Fuangthong, M. Regulation of Organic Hydroperoxide Stress Response by Two OhrR Homologs in Pseudomonas aeruginosa. PLoS ONE 2016, 11, e0161982. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, U.A.; Hassett, D.J.; Vasil, M.L. Genetic and Physiological Characterization Of ohr, Encoding a Protein Involved in Organic Hydroperoxide Resistance in Pseudomonas aeruginosa. J. Bacteriol. 2001, 183, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Lan, L.; Murray, T.S.; Kazmierczak, B.I.; He, C. Pseudomonas aeruginosa OspR Is an Oxidative Stress Sensing Regulator That Affects Pigment Production, Antibiotic Resistance and Dissemination during Infection. Mol. Microbiol. 2010, 75, 76–91. [Google Scholar] [CrossRef]
- Crack, J.C.; Green, J.; Hutchings, M.I.; Thomson, A.J.; Le Brun, N.E. Bacterial Iron-Sulfur Regulatory Proteins As Biological Sensor-Switches. Antioxid. Redox Signal. 2012, 17, 1215–1231. [Google Scholar] [CrossRef]
- Choi, Y.-S.; Shin, D.-H.; Chung, I.-Y.; Kim, S.-H.; Heo, Y.-J.; Cho, Y.-H. Identification of Pseudomonas aeruginosa Genes Crucial for Hydrogen Peroxide Resistance. J. Microbiol. Biotechnol. 2007, 17, 1344–1352. [Google Scholar]
- Saninjuk, K.; Romsang, A.; Duang-nkern, J.; Vattanaviboon, P.; Mongkolsuk, S. Transcriptional Regulation of the Pseudomonas aeruginosa Iron-Sulfur Cluster Assembly Pathway by Binding of IscR to Multiple Sites. PLoS ONE 2019, 14, e0218385. [Google Scholar] [CrossRef]
- Miller, H.K.; Auerbuch, V. Bacterial Iron-Sulfur Cluster Sensors in Mammalian Pathogens. Metallomics 2015, 7, 943–956. [Google Scholar] [CrossRef]
- Kim, S.-H.; Lee, B.-Y.; Lau, G.W.; Cho, Y.-H. IscR Modulates Catalase A (KatA) Activity, Peroxide Resistance and Full Virulence of Pseudomonas aeruginosa PA14. J. Microbiol. Biotechnol. 2009, 19, 1520–1526. [Google Scholar] [CrossRef]
- Romsang, A.; Duangnkern, J.; Wirathorn, W.; Vattanaviboon, P.; Mongkolsuk, S. Pseudomonas aeruginosa IscR-Regulated Ferredoxin NADP(+) Reductase Gene (FprB) Functions in Iron-Sulfur Cluster Biogenesis and Multiple Stress Response. PLoS ONE 2015, 10, e0134374. [Google Scholar] [CrossRef]
- Romsang, A.; Duangnkern, J.; Saninjuk, K.; Vattanaviboon, P.; Mongkolsuk, S. Pseudomonas aeruginosa NfuA: Gene Regulation and Its Physiological Roles in Sustaining Growth under Stress and Anaerobic Conditions and Maintaining Bacterial Virulence. PLoS ONE 2018, 13, e0202151. [Google Scholar] [CrossRef]
- Boonma, S.; Romsang, A.; Duangnkern, J.; Atichartpongkul, S.; Trinachartvanit, W.; Vattanaviboon, P.; Mongkolsuk, S. The FinR-Regulated Essential Gene FprA, Encoding Ferredoxin NADP+ Reductase: Roles in Superoxide-Mediated Stress Protection and Virulence of Pseudomonas aeruginosa. PLoS ONE 2017, 12, e0172071. [Google Scholar] [CrossRef]
- Prince, R.W.; Cox, C.D.; Vasil, M.L. Coordinate Regulation of Siderophore and Exotoxin A Production: Molecular Cloning and Sequencing of the Pseudomonas aeruginosa Fur Gene. J. Bacteriol. 1993, 175, 2589–2598. [Google Scholar] [CrossRef]
- Barton, H.A.; Johnson, Z.; Cox, C.D.; Vasil, A.I.; Vasil, M.L. Ferric Uptake Regulator Mutants of Pseudomonas aeruginosa with Distinct Alterations in the Iron-Dependent Repression of Exotoxin A and Siderophores in Aerobic and Microaerobic Environments. Mol. Microbiol. 1996, 21, 1001–1017. [Google Scholar] [CrossRef]
- Pérard, J.; Covès, J.; Castellan, M.; Solard, C.; Savard, M.; Miras, R.; Galop, S.; Signor, L.; Crouzy, S.; Michaud-Soret, I.; et al. Quaternary Structure of Fur Proteins, a New Subfamily of Tetrameric Proteins. Biochemistry 2016, 55, 1503–1515. [Google Scholar] [CrossRef] [PubMed]
- Wilderman, P.J.; Sowa, N.A.; FitzGerald, D.J.; FitzGerald, P.C.; Gottesman, S.; Ochsner, U.A.; Vasil, M.L. Identification of Tandem Duplicate Regulatory Small RNAs in Pseudomonas aeruginosa Involved in Iron Homeostasis. Proc. Natl. Acad. Sci. USA 2004, 101, 9792–9797. [Google Scholar] [CrossRef] [PubMed]
- Pasqua, M.; Visaggio, D.; Lo Sciuto, A.; Genah, S.; Banin, E.; Visca, P.; Imperi, F. Ferric Uptake Regulator Fur Is Conditionally Essential in Pseudomonas aeruginosa. J. Bacteriol. 2017, 199, e00472-17. [Google Scholar] [CrossRef] [PubMed]
- Ha, U.; Jin, S. Expression of the SoxR Gene of Pseudomonas aeruginosa Is Inducible during Infection of Burn Wounds in Mice and Is Required To Cause Efficient Bacteremia. Infect. Immun. 1999, 67, 5324–5331. [Google Scholar] [CrossRef]
- Koo, M.-S.; Lee, J.-H.; Rah, S.-Y.; Yeo, W.-S.; Lee, J.-W.; Lee, K.-L.; Koh, Y.-S.; Kang, S.-O.; Roe, J.-H. A Reducing System of the Superoxide Sensor SoxR in Escherichia coli. EMBO J. 2003, 22, 2614–2622. [Google Scholar] [CrossRef]
- Kobayashi, K. Sensing Mechanisms in the Redox-Regulated, [2Fe–2S] Cluster-Containing, Bacterial Transcriptional Factor SoxR. Acc. Chem. Res. 2017, 50, 1672–1678. [Google Scholar] [CrossRef]
- Singh, A.K.; Shin, J.-H.; Lee, K.-L.; Imlay, J.A.; Roe, J.-H. Comparative Study of SoxR Activation by Redox-Active Compounds. Mol. Microbiol. 2013, 90, 983–996. [Google Scholar] [CrossRef]
- Pomposiello, P.J.; Demple, B. Redox-Operated Genetic Switches: The SoxR and OxyR Transcription Factors. Trends Biotechnol. 2001, 19, 109–114. [Google Scholar] [CrossRef]
- Palma, M.; Zurita, J.; Ferreras, J.A.; Worgall, S.; Larone, D.H.; Shi, L.; Campagne, F.; Quadri, L.E.N. Pseudomonas aeruginosa SoxR Does Not Conform to the Archetypal Paradigm for SoxR-Dependent Regulation of the Bacterial Oxidative Stress Adaptive Response. Infect. Immun. 2005, 73, 2958–2966. [Google Scholar] [CrossRef] [PubMed]
- Aendekerk, S.; Ghysels, B.; Cornelis, P.; Baysse, C. Characterization of a New Efflux Pump, MexGHI-OpmD, from Pseudomonas aeruginosa That Confers Resistance to Vanadium. Microbiology 2002, 148, 2371–2381. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, L.E.P.; Price-Whelan, A.; Petersen, A.; Whiteley, M.; Newman, D.K. The Phenazine Pyocyanin Is a Terminal Signalling Factor in the Quorum Sensing Network of Pseudomonas aeruginosa. Mol. Microbiol. 2006, 61, 1308–1321. [Google Scholar] [CrossRef] [PubMed]
- Reen, F.J.; Haynes, J.M.; Mooij, M.J.; O’Gara, F. A Non-Classical LysR-Type Transcriptional Regulator PA2206 Is Required for an Effective Oxidative Stress Response in Pseudomonas aeruginosa. PLoS ONE 2013, 8, e54479. [Google Scholar] [CrossRef]
- Rungrassamee, W.; Ryan, K.C.; Maroney, M.J.; Pomposiello, P.J. The PqrR Transcriptional Repressor of Pseudomonas aeruginosa Transduces Redox Signals via an Iron-Containing Prosthetic Group. J. Bacteriol. 2009, 191, 6709–6721. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, C.; Pu, Q.; Wu, Q.; Tan, S.; Shao, X.; Zhang, W.; Xie, Y.; Li, R.; Yu, X.; et al. Pseudomonas aeruginosa Regulatory Protein AnvM Controls Pathogenicity in Anaerobic Environments and Impacts Host Defense. mBio 2019, 10, e01362-19. [Google Scholar] [CrossRef]
- Issa, K.H.B.; Phan, G.; Broutin, I. Functional Mechanism of the Efflux Pumps Transcription Regulators From Pseudomonas aeruginosa Based on 3D Structures. Front. Mol. Biosci. 2018, 5, 57. [Google Scholar] [CrossRef]
- Chen, H.; Hu, J.; Chen, P.R.; Lan, L.; Li, Z.; Hicks, L.M.; Dinner, A.R.; He, C. The Pseudomonas aeruginosa Multidrug Efflux Regulator MexR Uses an Oxidation-Sensing Mechanism. Proc. Natl. Acad. Sci. USA 2008, 105, 13586–13591. [Google Scholar] [CrossRef]
- Fraud, S.; Poole, K. Oxidative Stress Induction of the MexXY Multidrug Efflux Genes and Promotion of Aminoglycoside Resistance Development in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2011, 55, 1068–1074. [Google Scholar] [CrossRef]
- Fetar, H.; Gilmour, C.; Klinoski, R.; Daigle, D.M.; Dean, C.R.; Poole, K. MexEF-OprN Multidrug Efflux Operon of Pseudomonas aeruginosa: Regulation by the MexT Activator in Response to Nitrosative Stress and Chloramphenicol. Antimicrob. Agents Chemother. 2011, 55, 508–514. [Google Scholar] [CrossRef]
- Muller, J.F.; Stevens, A.M.; Craig, J.; Love, N.G. Transcriptome Analysis Reveals That Multidrug Efflux Genes Are Upregulated To Protect Pseudomonas aeruginosa from Pentachlorophenol Stress. Appl. Environ. Microbiol. 2007, 73, 4550–4558. [Google Scholar] [CrossRef] [PubMed]
- Mandsberg, L.F.; Ciofu, O.; Kirkby, N.; Christiansen, L.E.; Poulsen, H.E.; Høiby, N. Antibiotic Resistance in Pseudomonas aeruginosa Strains with Increased Mutation Frequency Due to Inactivation of the DNA Oxidative Repair System. Antimicrob. Agents Chemother. 2009, 53, 2483–2491. [Google Scholar] [CrossRef]
- Masuda, N.; Sakagawa, E.; Ohya, S.; Gotoh, N.; Tsujimoto, H.; Nishino, T. Substrate Specificities of MexAB-OprM, MexCD-OprJ, and MexXY-OprM Efflux Pumps in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 3322–3327. [Google Scholar] [CrossRef] [PubMed]
- Laborda, P.; Alcalde-Rico, M.; Blanco, P.; Martínez, J.L.; Hernando-Amado, S. Novel Inducers of the Expression of Multidrug Efflux Pumps That Trigger Pseudomonas aeruginosa Transient Antibiotic Resistance. Antimicrob. Agents Chemother. 2019, 63, 15. [Google Scholar] [CrossRef]
- Jin, M.; Liu, L.; Wang, D.; Yang, D.; Liu, W.; Yin, J.; Yang, Z.; Wang, H.; Qiu, Z.; Shen, Z.; et al. Chlorine Disinfection Promotes the Exchange of Antibiotic Resistance Genes across Bacterial Genera by Natural Transformation. ISME J. 2020, 14, 1847–1856. [Google Scholar] [CrossRef]
- Valentini, M.; Filloux, A. Biofilms and Cyclic Di-GMP (c-Di-GMP) Signaling: Lessons from Pseudomonas aeruginosa and Other Bacteria. J. Biol. Chem. 2016, 291, 12547–12555. [Google Scholar] [CrossRef]
- Strempel, N.; Nusser, M.; Neidig, A.; Brenner-Weiss, G.; Overhage, J. The Oxidative Stress Agent Hypochlorite Stimulates C-Di-GMP Synthesis and Biofilm Formation in Pseudomonas Aeruginosa. Front. Microbiol. 2017, 8, 2311. [Google Scholar] [CrossRef] [PubMed]
- Poudyal, B.; Sauer, K. The PA3177 Gene Encodes an Active Diguanylate Cyclase That Contributes to Biofilm Antimicrobial Tolerance but Not Biofilm Formation by Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2018, 62, e01049-18. [Google Scholar] [CrossRef]
- Xia, B.; Li, M.; Tian, Z.; Chen, G.; Liu, C.; Xia, Y.; Jin, Y.; Bai, F.; Cheng, Z.; Jin, S.; et al. Oligoribonuclease Contributes to Tolerance to Aminoglycoside and β-Lactam Antibiotics by Regulating KatA in Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 2019, 63, e00212-19. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Chen, G.; Liu, Y.; Jin, Y.; Cheng, Z.; Liu, Y.; Yang, L.; Jin, S.; Wu, W. Pseudomonas aeruginosa Oligoribonuclease Contributes to Tolerance to Ciprofloxacin by Regulating Pyocin Biosynthesis. Antimicrob. Agents Chemother. 2017, 61, e02256-16. [Google Scholar] [CrossRef] [PubMed]
- Goldman, S.R.; Sharp, J.S.; Vvedenskaya, I.O.; Livny, J.; Dove, S.L.; Nickels, B.E. NanoRNAs Prime Transcription Initiation In Vivo. Mol. Cell 2011, 42, 817–825. [Google Scholar] [CrossRef]
- Chen, G.; Zhao, Q.; Zhu, F.; Chen, R.; Jin, Y.; Liu, C.; Pan, X.; Jin, S.; Wu, W.; Cheng, Z. Oligoribonuclease Is Required for the Type III Secretion System and Pathogenesis of Pseudomonas Aeruginosa. Microbiol. Res. 2016, 188–189, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Orr, M.W.; Donaldson, G.P.; Severin, G.B.; Wang, J.; Sintim, H.O.; Waters, C.M.; Lee, V.T. Oligoribonuclease Is the Primary Degradative Enzyme for PGpG in Pseudomonas aeruginosa That Is Required for Cyclic-Di-GMP Turnover. Proc. Natl. Acad. Sci. USA 2015, 112, E5048–E5057. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, D.; Kumari, H.; Mathee, K. Pseudomonas aeruginosa AmpR: An Acute-Chronic Switch Regulator. Pathog. Dis. 2015, 73, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, N.; Nur, I.; Noor, R. Comparative Study of Oxidative Stress Response in Pseudomonas aeruginosa (Subp01) and Pseudomonas Fluorescence (Subp02) Upon External Stimulation by Hydrogen Peroxide. EC Microbiol. 2019, 15, 653–660. [Google Scholar]
- Kiewitz, C.; Tümmler, B. Sequence Diversity of Pseudomonas aeruginosa: Impact on Population Structure and Genome Evolution. J. Bacteriol. 2000, 182, 3125–3135. [Google Scholar] [CrossRef]
- Frimmersdorf, E.; Horatzek, S.; Pelnikevich, A.; Wiehlmann, L.; Schomburg, D. How Pseudomonas aeruginosa Adapts to Various Environments: A Metabolomic Approach. Environ. Microbiol. 2010, 12, 1734–1747. [Google Scholar] [CrossRef]
- Moradali, M.F.; Ghods, S.; Rehm, B.H.A. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 5 May 2021).
- Skindersoe, M.E.; Alhede, M.; Phipps, R.; Yang, L.; Jensen, P.O.; Rasmussen, T.B.; Bjarnsholt, T.; Tolker-Nielsen, T.; Høiby, N.; Givskov, M. Effects of Antibiotics on Quorum Sensing in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2008, 52, 3648–3663. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, D.J.; Keyser, R. Effects of Subinhibitory Concentrations of Macrolide Antibiotics on Pseudomonas aeruginosa. Chest 2004, 125, 62S–69S. [Google Scholar] [CrossRef]
- Puckett, S.; Trujillo, C.; Wang, Z.; Eoh, H.; Ioerger, T.R.; Krieger, I.; Sacchettini, J.; Schnappinger, D.; Rhee, K.Y.; Ehrt, S. Glyoxylate Detoxification Is an Essential Function of Malate Synthase Required for Carbon Assimilation in Mycobacterium Tuberculosis. Proc. Natl. Acad. Sci. USA 2017, 114, E2225–E2232. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Cruz Nizer, W.S.; Inkovskiy, V.; Versey, Z.; Strempel, N.; Cassol, E.; Overhage, J. Oxidative Stress Response in Pseudomonas aeruginosa. Pathogens 2021, 10, 1187. https://doi.org/10.3390/pathogens10091187
da Cruz Nizer WS, Inkovskiy V, Versey Z, Strempel N, Cassol E, Overhage J. Oxidative Stress Response in Pseudomonas aeruginosa. Pathogens. 2021; 10(9):1187. https://doi.org/10.3390/pathogens10091187
Chicago/Turabian Styleda Cruz Nizer, Waleska Stephanie, Vasily Inkovskiy, Zoya Versey, Nikola Strempel, Edana Cassol, and Joerg Overhage. 2021. "Oxidative Stress Response in Pseudomonas aeruginosa" Pathogens 10, no. 9: 1187. https://doi.org/10.3390/pathogens10091187
APA Styleda Cruz Nizer, W. S., Inkovskiy, V., Versey, Z., Strempel, N., Cassol, E., & Overhage, J. (2021). Oxidative Stress Response in Pseudomonas aeruginosa. Pathogens, 10(9), 1187. https://doi.org/10.3390/pathogens10091187