
Staphylococcus aureus and Cystic Fibrosis—A Close Relationship. What Can We Learn from Sequencing Studies?


Abstract
1. Pathogenesis of S. aureus with Respect to Cystic Fibrosis
1.1. Adaptation to CF Airways
1.2. Clinical Impact
1.3. S. aureus—Fundamentals of Pathogenicity
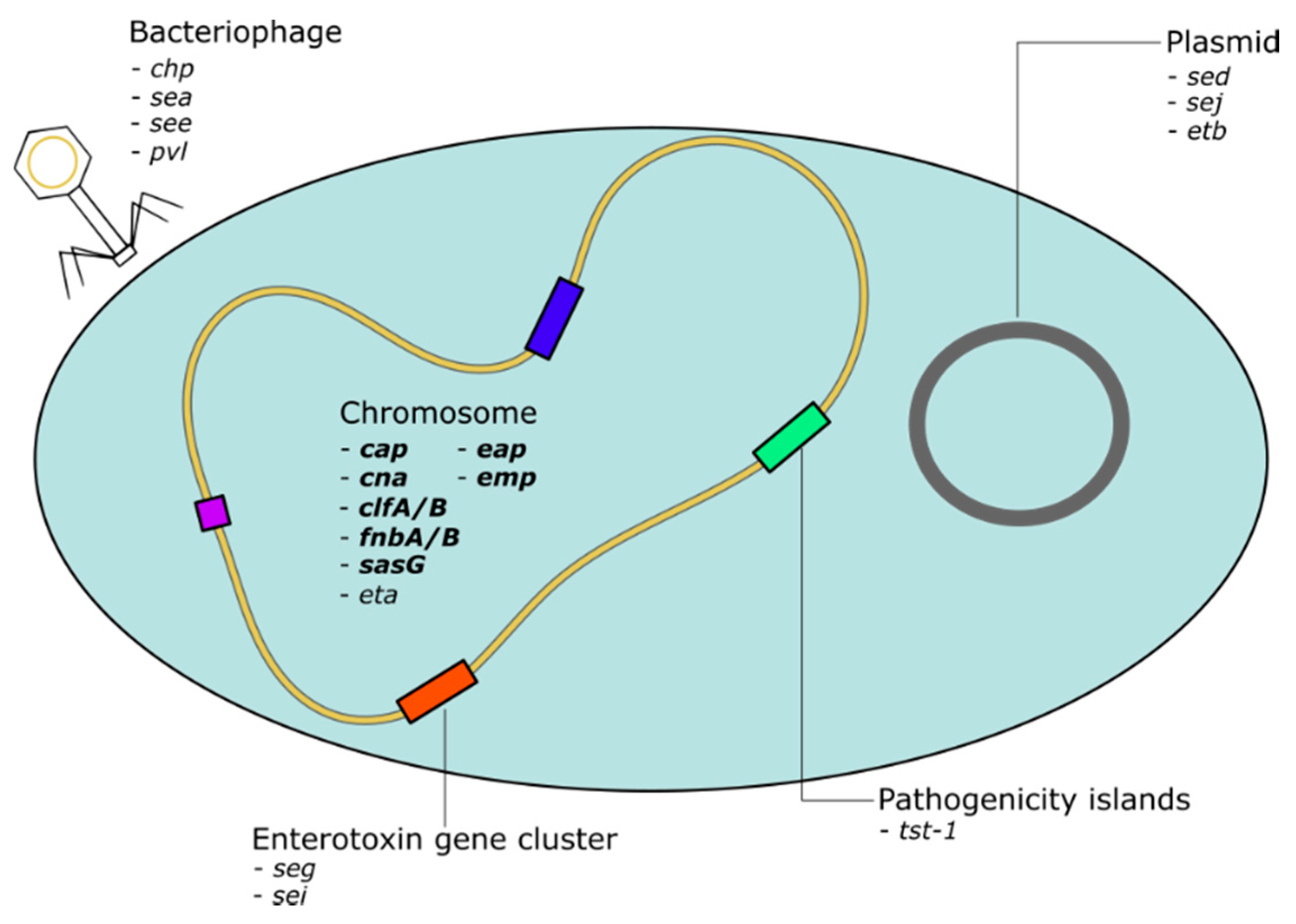
1.4. S. aureus Virulence Genome
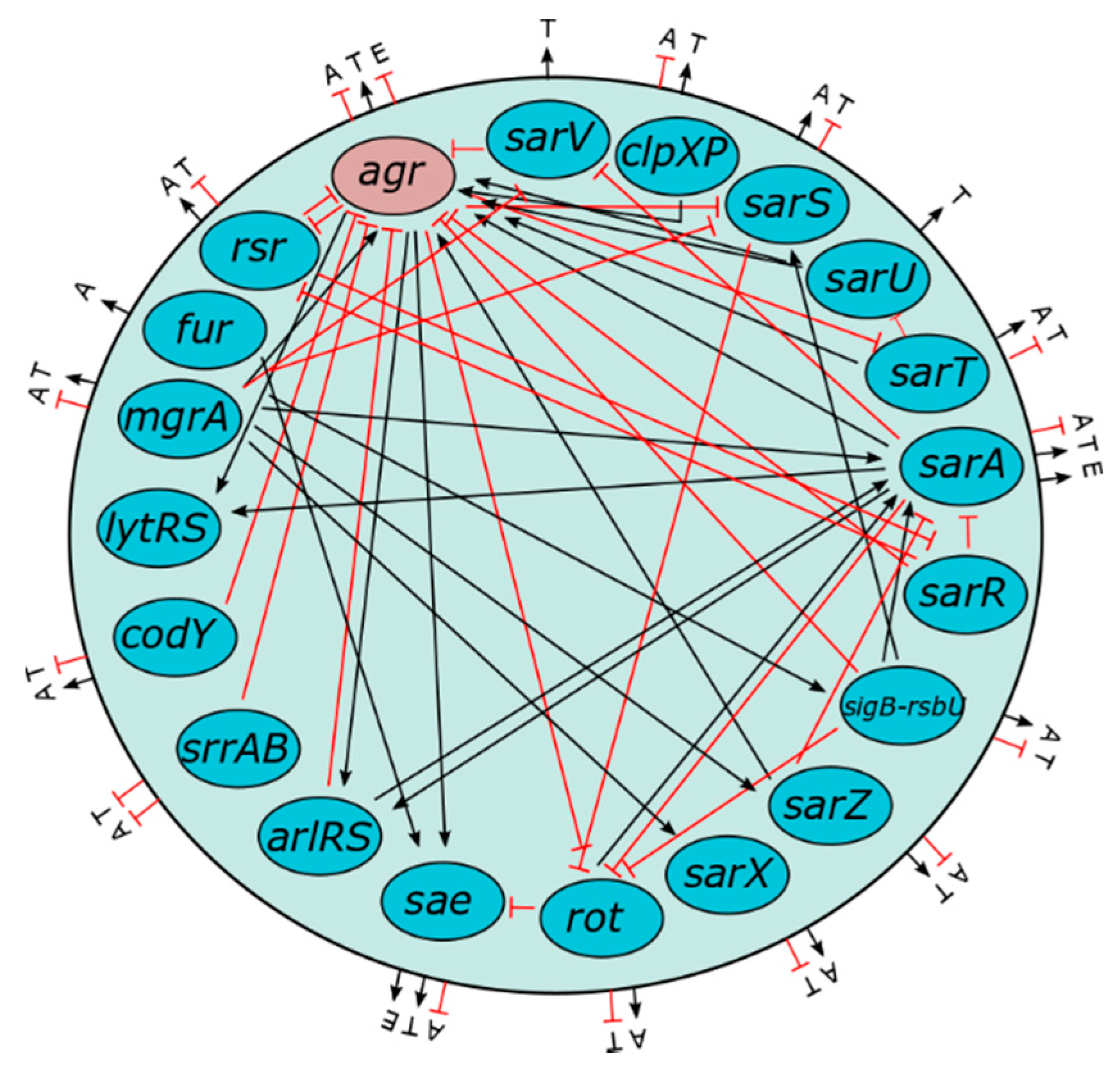
1.5. Role of Agr in Virulence Gene Regulation Via Quorum Sensing
1.6. Role of Horizontal Gene Transfer in the Spread of Virulence Genes
1.7. Impact of Varying Genotypes and Single Genes on S. aureus Pathogenicity
- The considered virulence gene should be associated with pathogenic clones or strains;
- The inactivation of the gene should measurably decrease virulence, and
- The reactivation of the gene should restore pathogenicity.
2. Genotyping of S. aureus in Cystic Fibrosis
2.1. Adaptation of S. aureus towards Persistence in Cystic Fibrosis Using WGS
2.2. S. aureus Population Dynamics in CF
2.3. MRSA in Cystic Fibrosis—Antibiotic Resistance, Persistence and Patho-Adaptation
2.4. Biofilm Formation—An Adaptive Mechanism of S. aureus in Cystic Fibrosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Authors | Number of Patients | Length of Study [Years] | Number of S. aureus Isolates for WGS | Sequencing Techniques | S. aureus Methicillin Resistance [%] | Most Important Outcome |
|---|---|---|---|---|---|---|
| Antonelli et al., 2016 [122] | 1 | N/A | 1 | WGS SCCmec-typing | 100 | - the MRSA isolate was linezolid resistant and carried a cfr gene. The strain belonged to the ST5-MRSA-II lineage spa-type t306 |
| Ankrum et al., 2017 [99] | 115 | 1.8 | 301 | - WGS - cgMLST -SCCmec typing | 70 | - definition of relatedness of strains by WGS: same strains: less than 71 SNPs very closely related strains: between 72 and </= 123 SNPs closely related strains: between 124 and </=156 SNPs distantly related strains: between 157 and </= 1514 SNPs unrelated strains: between 5957 and 21,644 SNPs- prediction tool PPFS2 was successful in predicting MRSA phenotype- Sequential isolates of patients did not follow a molecular clock (multiple strains at the same time that were not isolated?) |
| Azarian et al., 2019 [101] | 4 | 3.8 | 85 | - WGS - cgMLST - spa-typing | 100 | - peristence of a dominant lineage- intrahost diversity: variation in antibiotic resistance, mutations in eleven genes of three patients, clpX, odHA, fmtC, mfd among others- rates of molecular evolution between 2.21 and 8.64 nucleotide polymorphisms per year- gene-wide diversifying selection of spa |
| Bernardy et al., 2019 [134] | 50 | N/A | 65 | - WGS | - sequences are provided online - no further analysis performed | |
| Bernardy et al., 2020 [135] | 50 | N/A | 64 | - WGS - MLST - spa-types - agr-types | 61 | - variety of genotypes, not a specific “CF-clone”, CC5 and CC8 most common - CC8 clones carried PVL - between patients no transmission - all isolates carried alpha- and beta-toxins and many other virulence genes - isolates group together in terms of polysaccharide production - genetic association based on the status of coinfection with P. aeruginosa- close relatedness of longitudinal isolates sharing similar phenotypes |
| Boudet et al., 2021 [124] | 2 | 7 | 17 | - WGS | 100 | - highly diversified clonally related populations - resistance to linezolid due to a G2576T substitution in a variable number of 23S rRNA gene copies - intrasample heterogeneity of linezolid susceptible/resistant strains |
| Gabryszewski et al., 2019 [133] | 3 | 4 | 12 | - WGS | 100 | - association of persistent MRSA infection with staphyloccocal metabolic change- altered carbon metabolism: adaption by selective use of tricarboxylic acid cycle and biofilm formation- increased transcription of specific metabolic genes, most significantly fumC associated with decreased induction of proinflammatory cytokines |
| Haim et al., 2021 [112] | 4 | 0.7 | 20 | - spa-typing (Sanger) - MLST (Sanger) - WGS | 100 | - intrahost diversity- genetic differences in sequential isolates in both core and accessory genomes - independent mutations in thyA, htrA, rspJ and gyrA- non-synonymous mutations in genes associated with antimicrobial resistance, virulence, iron scavenging and oxidative stress resistance |
| Herzog et al., 2019 [103] | 1 | 14 | 7 | - spa-typing (Sanger) - WGS/cgMLST | - inverse expression of nucleases and agr during long-term adaption- high nuclease activity facilitates survival with NETs- mutation in agr most likely responsible for increased nuclease | |
| Lange et al., 2020 [100] | 195 | 1.75 | - | - spa-typing (Sanger) (3180) - multiplex PCR for adhesin genes: clfA/B,cna, eap, emp,fnbA/B, sasG/H, sdrC/D/E toxin genes: eta,etb, hlg, pvl, sea-j,tst immune evasion genes: chip, cap5/8 | - association of agr-type 1 and 4 with clinical status- virulence gene count varies with agr-type- loss of virulence genes during persistence- increased virulence gene plasticity in the presence of environmental stress (antibiotic therapy/inflammation) | |
| Langhanki et al., 2018 [102] | 1 | 13 | 75 | - spa-typing (Sanger) - MLST - WGS | - several lineages were present, but only two lineages co-existed for extended periods - genomic elements are transferred between lineages- increase in genomic changes - adaptation of successful lineages and elimination of inferior lineages | |
| Long et al., 2014 [128] | 1 | 0.4 | 6 | - WGS | 100 | - 2 SNPs in PBP2a leading to ceftaroline resistance- mutation in thyA leading to the SCV phenotype |
| Long et al., 2021 [93] | 246 | 2.2 | 1382 | - WGS -wgMLST - SCCmec-typing (WGS) | 28 | - frequent co-infection by multiple, genetically distinct S. aureus strains often differing in antibiotic susceptibility- identification of common set of S. aureus genes frequently mutated during long-term adaptation (rpoB,fmtB, agrA, rpsJ, rsbU, thyA, set9, walK, ebh)- contribution to antimicrobial resistance (rpsJ) and bacterial persistence (agrA, rsbU, ebh) or both (rpoB,thyA) - open pangenome with incoming new genes |
| Lopez-Collazo et al., 2015 [127] | 1 | 13 | 2 | - WGS SCCmec-typing (Sanger) - spa-typing (Sanger) | 100 | - reduction of bacterial chromosome - loss of one of three phages in the late isolate compared to the early isolate - decrease in immune response |
| McAdam et al., 2011 [91] | 1 | 2.2 | 3 | - WGS | - variation in phage content - genetic polymorphism in genes, which impact antibiotic resistance and global regulation of virulence - genetic variations correlated with phenotypic changes in terms of hemolytic activity, antibiotic sensitivity, growth rate | |
| Ormerod et al., 2015 [114] | 3 | 1 | 6 | - WGS - MLST | 0 | - all patients carried different strains - persistent infection in one patient - different virulence factor profiles in strains |
| Porterfield et al., 2021 [120] | 74 | N/A | 97 | - WGS | 100 | - most prevalent ST5 (HA-MRSA) and ST8 (CA-MRSA) - closely related MRSA strains only shared by family members - accessory gene content discriminates 3 clusters Cluster A: including ST5 and ST105, highly prevalent at all ages Cluster B: including ST8, younger patients Cluster C: only 6 distantly related strains - patients infected by cluster A had lower lung function and higher sputum mass compared to age matched patients infected by cluster B |
| Roch et al. 2019 [125] | 2 | N/A | 2 | - WGS | - underlying mutations for tedizolid resistance identified - G2576T mutation in rrl and new mutations in rplA and rspQ - functional experimental validation of the new mutations necessary | |
| Rolain et al. 2009 [126] | 1 | N/A | 1 | - WGS | 100 | - CF-Marseille clone with a hetero-glycopeptide Intermediate resistance - discovery of a new SCCmec type IV mosaic cassette and a new phage |
| Rouard et al., 2018 [123] | 1 | 5 | 7 | - spa typing (Sanger) - MLST (Sanger) - wgMLST | 100 | - G2576T mutation in all 5 rrl copies leading to linezolid resistance - correlation of MIC and mutated copies of rrl |
| Schwartbeck et al., 2016 [132] | 2 | 1.2 | 12 | - WGS - cgMLST | - 2.5% of patients with mucoid S. aureus isolates - closely related strains within an individual patient, but not between patients - 5bp deletion in the intergenic region of the ica operon responsible for the mucoid phenotype | |
| Tan et al. 2019 [6] | 3 | 6 | 6 | - WGS - MLST | - long-term persistence of the same clone in patients - identification of underlying genetic modifications that induce protein expression profiles and metabolic changes of late isolates compared to early isolates (biofilm, SCV, intracellular persistence) - changes were identified in e.g., agrC, saeR, thyA, gyrB, fakA | |
| Tomlinson et al. 2021 [136] | 1 | 14 | 7 | - WGS | - S. aureus induces an itaconate-dominated immunometabolic response in immune cells - itaconate inhibited S. aureus glycolysis and redirected carbon flux to the formation of polysaccharide biofilm - longitudinal isolates revealed numerous SNPs in virulence genes and regulators and genes involved in metabolism | |
| Westphal et al., 2020 [94] | 8 | 1.75 | 24 | - spa-typing (Sanger) (3893 isolates 183 patients) | - no association of spa-type and lung function - association of antibiotic therapy and age on carriage profile of spa-types during persistence- antibiotic treatment most likely prevents acquisition of new S. aureus clones - probability of dominant or related spa-types increases with age - related spa-types are mainly caused by deletions in the VNTR region - isolates with related spa-types belong to a clonal lineage as confirmed by WGS |
3. Summary and Conclusions
- There is not “a” particular CF clone, but CF patients are colonized and infected by various S. aureus clones, which are also common in both healthy and diseased humans.
- CF patients are infected by coexisting lineages; however, a dominant clone largely persists.
- There is limited transmission of clones within the CF population.
- The sequencing of long-term persistent sequential isolates reveals evolution and adaptation of isolates, some of them being convergent, especially as mutations in important virulence regulators such as agr and sigB, were observed. Additionally, mutations in thyA were often present especially in centres where patients are treated with trimethoprim-sulfamethoxazole.
- During persistence, S. aureus changes from an acute virulent to a chronic non-virulent isolate.
- S. aureus has an open pangenome with new incoming genes from coinfecting lineages.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Sullivan, B.P.; Freedman, S.D. Cystic fibrosis. Lancet 2009, 373, 1891–1904. [Google Scholar] [CrossRef]
- Elborn, J.S. Cystic fibrosis. Lancet 2016, 388, 2519–2531. [Google Scholar] [CrossRef]
- Cystic Fibrosis Foundation Patient Registry 2019 Annual Data Report, Bethesda, Maryland, 2020, Cystuc Fibrosis Foundation. Available online: https://www.cff.org/Research/Researcher-Resources/Patient-Registry/2019-Patient-Registry-Annual-Data-Report (accessed on 11 September 2021).
- Zolin, A.; Orenti, A.; Naehrlich, L.; Jung, A.; van Rens, J. European Cystic Fibrosis Society Patient Registry Annual Report 2018. 2020. Available online: https://www.ecfs.eu/sites/default/files/general-content-files/working-groups/ecfs-patient-registry/ECFSPR_Report_2018_v1.4 (accessed on 11 September 2021).
- Schwerdt, M.; Neumann, C.; Schwartbeck, B.; Kampmeier, S.; Herzog, S.; Görlich, D.; Dübbers, A.; Große-Onnebrink, J.; Kessler, C.; Küster, P.; et al. Staphylococcus aureus in the airways of cystic fibrosis patients—A retrospective long-term study. Int. J. Med. Microbiol. 2018, 308, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Coureuil, M.; Ramond, E.; Euphrasie, D.; Dupuis, M.; Tros, F.; Meyer, J.; Nemazanyy, I.; Chhuon, C.; Guerrera, I.C.; et al. Chronic Staphylococcus aureus Lung Infection Correlates with Proteogenomic and Metabolic Adaptations Leading to an Increased Intracellular Persistence. Clin. Infect. Dis. 2019, 69, 1937–1945. [Google Scholar] [CrossRef] [PubMed]
- Branger, C.; Gardye, C.; Lambert-Zechovsky, N. Persistence of Staphylococcus aureus strains among cystic fibrosis patients over extended periods of time. J. Med. Microbiol. 1996, 45, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Kahl, B.C.; Duebbers, A.; Lubritz, G.; Haeberle, J.; Koch, H.G.; Ritzerfeld, B.; Reilly, M.; Harms, E.; Proctor, R.A.; Herrmann, M.; et al. Population Dynamics of Persistent Staphylococcus aureus Isolated from the Airways of Cystic Fibrosis Patients during a 6-Year Prospective Study. J. Clin. Microbiol. 2003, 41, 4424–4427. [Google Scholar] [CrossRef] [PubMed]
- Delaisi, B.; Grosskopf, C.; Reignault, E.; Goehrs, J.M.; Navarro, J. International registry on mucoviscidosis: Comparison of the French data with the European data for 1995. Arch. Pediatrie Organe Off. Soc. Fr. Pediatrie 1998, 5, 384–388. [Google Scholar]
- Ahmed, M.I.; Mukherjee, S. Treatment for chronic methicillin-sensitive Staphylococcus aureus pulmonary infection in people with cystic fibrosis. Cochrane Database Syst. Rev. 2018, 7, CD011581. [Google Scholar] [CrossRef]
- Dasenbrook, E.C. Update on methicillin-resistant Staphylococcus aureus in cystic fibrosis. Curr. Opin. Pulm. Med. 2011, 17, 437–441. [Google Scholar] [CrossRef]
- Chmiel, J.F.; Aksamit, T.R.; Chotirmall, S.H.; Dasenbrook, E.C.; Elborn, J.S.; LiPuma, J.J.; Ranganathan, S.C.; Waters, V.J.; Ratjen, F.A. Antibiotic management of lung infections in cystic fibrosis. I. The microbiome, methicillin-resistant Staphylococcus aureus, gram-negative bacteria, and multiple infections. Ann. Am. Thorac. Soc. 2014, 11, 1120–1129. [Google Scholar] [CrossRef]
- Zobell, J.T.; Epps, K.L.; Young, D.C.; Montague, M.; Olson, J.; Ampofo, K.; Chin, M.J.; Marshall, B.C.; Dasenbrook, E. Utilization of antibiotics for methicillin-resistant Staphylococcus aureus infection in cystic fibrosis. Pediatr. Pulmonol. 2015, 50, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Pennoni, G.; Mencarini, V.; Palladino, N.; Peccini, L.; Principi, N. Antimicrobial Treatment of Staphylococcus aureus in Patients With Cystic Fibrosis. Front. Pharmacol. 2019, 10, 849. [Google Scholar] [CrossRef] [PubMed]
- Goerke, C.; Wolz, C. Adaptation of Staphylococcus aureus to the cystic fibrosis lung. Int. J. Med. Microbiol. 2010, 300, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Goerke, C.; Papenberg, S.M.; Dasbach, S.; Dietz, K.; Ziebach, R.; Kahl, B.C.; Wolz, C. Increased frequency of genomic alterations in Staphylococcus aureus during chronic infection is in part due to phage mobilization. J. Infect. Dis. 2004, 189, 724–734. [Google Scholar] [CrossRef] [PubMed]
- Priest, N.K.; Rudkin, J.K.; Feil, E.J.; Van Den Elsen, J.M.; Cheung, A.; Peacock, S.J.; Laabei, M.; Lucks, D.A.; Recker, M.; Massey, R.C. From genotype to phenotype: Can systems biology be used to predict Staphylococcus aureus virulence? Nat. Rev. Microbiol. 2012, 10, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Kahl, B.; Herrmann, M.; Everding, A.S.; Koch, H.G.; Becker, K.; Harms, E.; Proctor, R.A.; Peters, G. Persistent Infection with Small Colony Variant Strains of Staphylococcus aureus in Patients with Cystic Fibrosis. J. Infect. Dis. 1998, 177, 1023–1029. [Google Scholar] [CrossRef] [PubMed]
- Ziebandt, A.-K.; Kusch, H.; Degner, M.; Jaglitz, S.; Sibbald, M.J.J.B.; Arends, J.P.; Chlebowicz, M.A.; Albrecht, D.; Pantucek, R.; Doškar, J.; et al. Proteomics uncovers extreme heterogeneity in the Staphylococcus aureus exoproteome due to genomic plasticity and variant gene regulation. Proteomics 2010, 10, 1634–1644. [Google Scholar] [CrossRef] [PubMed]
- Liew, Y.K.; Hamat, R.A.; Van Belkum, A.; Chong, P.P.; Neela, V. Comparative Exoproteomics and Host Inflammatory Response in Staphylococcus aureus Skin and Soft Tissue Infections, Bacteremia, and Subclinical Colonization. Clin. Vaccine Immunol. 2015, 22, 593–603. [Google Scholar] [CrossRef]
- Goss, C.H.; Muhlebach, M.S. Review: Staphylococcus aureus and MRSA in cystic fibrosis. J. Cyst. Fibros. 2011, 10, 298–306. [Google Scholar] [CrossRef]
- Goerke, C.; Gressinger, M.; Endler, K.; Breitkopf, C.; Wardecki, K.; Stern, M.; Wolz, C.; Kahl, B.C. High phenotypic diversity in infecting but not in colonizing Staphylococcus aureus populations. Environ. Microbiol. 2007, 9, 3134–3142. [Google Scholar] [CrossRef]
- Besier, S.; Zander, J.; Siegel, E.; Saum, S.H.; Hunfeld, K.-P.; Ehrhart, A.; Brade, V.; Wichelhaus, T.A. Thymidine-Dependent Staphylococcus aureus Small-Colony Variants: Human Pathogens That Are Relevant Not Only in Cases of Cystic Fibrosis Lung Disease. J. Clin. Microbiol. 2008, 46, 3829–3832. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hirschhausen, N.; Block, D.; Bianconi, I.; Bragonzi, A.; Birtel, J.; Lee, J.; Dübbers, A.; Küster, P.; Kahl, J.; Peters, G.; et al. Extended Staphylococcus aureus persistence in cystic fibrosis is associated with bacterial adaptation. Int. J. Med. Microbiol. 2013, 303, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Hogardt, M.; Schubert, S.; Adler, K.; Götzfried, M.; Heesemann, J. Sequence variability and functional analysis of MutS of hypermutable Pseudomonas aeruginosa cystic fibrosis isolates. Int. J. Med. Microbiol. 2006, 296, 313–320. [Google Scholar] [CrossRef]
- Bragonzi, A.; Paroni, M.; Nonis, A.; Cramer, N.; Montanari, S.; Rejman, J.; DI Serio, M.S.; Döring, G.; Tümmler, B. Pseudomonas aeruginosa Microevolution during Cystic Fibrosis Lung Infection Establishes Clones with Adapted Virulence. Am. J. Respir. Crit. Care Med. 2009, 180, 138–145. [Google Scholar] [CrossRef]
- Wieneke, M.K.; Dach, F.; Neumann, C.; Görlich, D.; Kaese, L.; Thißen, T.; Dübbers, A.; Kessler, C.; Große-Onnebrink, J.; Küster, P.; et al. Association of Diverse Staphylococcus aureus Populations with Pseudomonas aeruginosa Coinfection and Inflammation in Cystic Fibrosis Airway Infection. mSphere 2021, 6, e0035821. [Google Scholar] [CrossRef]
- de Souza, D.C.; Cogo, L.L.; Palmeiro, J.K.; Dalla-Costa, L.M.; de Oliveira Tomaz, A.P.; Riedi, C.A.; Rosario Filho, N.A. Thymidine-auxotrophic Staphylococcus aureus small-colony variant bacteremia in a patient with cystic fibrosis. Pediatric Pulmonol. 2020, 55, 1388–1393. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.M.; Rourk, M.H.; Spock, A. Bacteremia in Patients with Cystic Fibrosis. Clin. Pediatr. 1980, 19, 746–748. [Google Scholar] [CrossRef]
- Kahl, B.C. Impact of Staphylococcus aureus on the pathogenesis of chronic cystic fibrosis lung disease. Int. J. Med. Microbiol. 2010, 300, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Goerke, C.; Campana, S.; Bayer, M.G.; Döring, G.; Botzenhart, K.; Wolz, C. Direct Quantitative Transcript Analysis of the agr Regulon of Staphylococcus aureus during Human Infection in Comparison to the Expression Profile In Vitro. Infect. Immun. 2000, 68, 1304–1311. [Google Scholar] [CrossRef]
- Wolter, D.J.; Emerson, J.C.; McNamara, S.; Buccat, A.M.; Qin, X.; Cochrane, E.; Houston, L.S.; Rogers, G.B.; Marsh, P.; Prehar, K.; et al. Staphylococcus aureus small-colony variants are independently associated with worse lung disease in children with cystic fibrosis. Clin. Infect. Dis. 2013, 57, 384–391. [Google Scholar] [CrossRef]
- Besier, S.; Smaczny, C.; von Mallinckrodt, C.; Krahl, A.; Ackermann, H.; Brade, V.; Wichelhaus, T.A. Prevalence and Clinical Significance of Staphylococcus aureus Small-Colony Variants in Cystic Fibrosis Lung Disease. J. Clin. Microbiol. 2007, 45, 168–172. [Google Scholar] [CrossRef]
- Wolter, D.J.; Onchiri, F.M.; Emerson, J.; Precit, M.R.; Lee, M.; McNamara, S.; Nay, L.; Blackledge, M.; Uluer, A.; Orenstein, D.M.; et al. Prevalence and clinical associations of Staphylococcus aureus small-colony variant respiratory infection in children with cystic fibrosis (SCVSA): A multicentre, observational study. Lancet Respir. Med. 2019, 7, 1027–1038. [Google Scholar] [CrossRef]
- Gangell, C.; Gard, S.; Douglas, T.; Park, J.; de Klerk, N.; Keil, T.; Brennan, S.; Ranganathan, S.; Robins-Browne, R.; Sly, P.D.; et al. Inflammatory Responses to Individual Microorganisms in the Lungs of Children With Cystic Fibrosis. Clin. Infect. Dis. 2011, 53, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Sagel, S.D.; Gibson, R.L.; Emerson, J.; McNamara, S.; Burns, J.L.; Wagener, J.S.; Ramsey, B.W. Impact of Pseudomonas and Staphylococcus Infection on Inflammation and Clinical Status in Young Children with Cystic Fibrosis. J. Pediatr. 2009, 154, 183–188.e3. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.S.; Hook, S.; Jamsen, K.M.; Nixon, G.M.; Carzino, R.; Carlin, J.; Robertson, C.F.; Grimwood, K. Lower Airway Inflammation in Infants with Cystic Fibrosis Detected by Newborn Screening. Pediatr. Pulmonol. 2005, 40, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Smyth, A.R.; Walters, S. Prophylactic anti-staphylococcal antibiotics for cystic fibrosis. Cochrane Database Syst Rev. 2003, CD001912. [Google Scholar] [CrossRef]
- Smyth, A.R.; Bell, S.; Bojcin, S.; Bryon, M.; Duff, A.; Flume, P.; Kashirskaya, N.; Munck, A.; Ratjen, F.; Schwarzenberg, S.J.; et al. European Cystic Fibrosis Society Standards of Care: Best Practice guidelines. J. Cyst. Fibros. 2014, 13, S23–S42. [Google Scholar] [CrossRef]
- Smyth, A. Prophylactic Antibiotics in Cystic Fibrosis: A Conviction without Evidence? Pediatr. Pulmonol. 2005, 40, 471–476. [Google Scholar] [CrossRef]
- Stone, A.; Saiman, L. Update on the epidemiology and management of Staphylococcus aureus, including methicillin-resistant Staphylococcus aureus, in patients with cystic fibrosis. Curr. Opin. Pulm. Med. 2007, 13, 515–521. [Google Scholar] [CrossRef]
- Junge, S.; Görlich, D.; Reijer, M.D.; Wiedemann, B.; Tümmler, B.; Ellemunter, H.; Dübbers, A.; Küster, P.; Ballmann, M.; Koerner-Rettberg, C.; et al. Factors Associated with Worse Lung Function in Cystic Fibrosis Patients with Persistent Staphylococcus aureus. PLoS ONE 2016, 11, e0166220. [Google Scholar] [CrossRef]
- Lindsay, J.A. Staphylococci: Evolving Genomes. Microbiol. Spectr. 2019, 7, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, J.; Moore, C.; Day, N.P.; Peacock, S.J.; Witney, A.A.; Stabler, R.; Husain, S.E.; Butcher, P.D.; Hinds, J. Microarrays Reveal that Each of the Ten Dominant Lineages of Staphylococcus aureus Has a Unique Combination of Surface-Associated and Regulatory Genes. J. Bacteriol. 2006, 188, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, J.A. Genomic variation and evolution of Staphylococcus aureus. Int. J. Med. Microbiol. 2010, 300, 98–103. [Google Scholar] [CrossRef]
- Melles, D.C.; Gorkink, R.F.J.; Boelens, H.A.M.; Snijders, S.V.; Peeters, J.K.; Moorhouse, M.J.; Van Der Spek, P.J.; Van Leeuwen, W.B.; Simons, G.; Verbrugh, H.A.; et al. Natural population dynamics and expansion of pathogenic clones of Staphylococcus aureus. J. Clin. Investig. 2004, 114, 1732–1740. [Google Scholar] [CrossRef]
- Grumann, D.; Nübel, U.; Bröker, B.M. Staphylococcus aureus toxins—Their functions and genetics. Infect. Genet. Evol. 2014, 21, 583–592. [Google Scholar] [CrossRef]
- Kusch, H.; Engelmann, S. Secrets of the secretome in Staphylococcus aureus. Int. J. Med. Microbiol. 2014, 304, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Recsei, P.; Kreiswirth, B.; O’Reilly, M.; Schlievert, P.; Gruss, A.; Novick, R. Regulation of exoprotein gene expression in Staphylococcus aureus by agr. Mol. Genet. Genom. 1986, 202, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Höök, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Genet. 2013, 12, 49–62. [Google Scholar] [CrossRef]
- Smeltzer, M.; Lee, C.Y.; Harik, N.; Hart, M.E. Molecular Basis of Pathogenicity. In Staphylococci in Human Disease, 2nd ed.; Crossley, K.B., Jefferson, K.K., Archer, G.I., Fowler, V.G., Jr., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Mukherjee, S.; Bassler, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Genet. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Novick, R.P.; Ross, H.F.; Projan, S.J.; Kornblum, J.; Kreiswirth, B.; Moghazeh, S. Synthesis of staphylococcal virulence factors is controlled by a regulatory RNA molecule. EMBO J. 1993, 12, 3967–3975. [Google Scholar] [CrossRef]
- Jarraud, S.; Mougel, C.; Thioulouse, J.; Lina, G.; Meugnier, H.; Forey, F.; Nesme, X.; Etienne, J.; Vandenesch, F. Relationships between Staphylococcus aureus Genetic Background, Virulence Factors, agr Groups (Alleles), and Human Disease. Infect. Immun. 2002, 70, 631–641. [Google Scholar] [CrossRef]
- Traber, K.; Lee, E.; Benson, S.; Corrigan, R.; Cantera, M.; Shopsin, B.; Novick, R.P. agr function in clinical Staphylococcus aureus isolates. Microbiology 2008, 154, 2265–2274. [Google Scholar] [CrossRef] [PubMed]
- Goerke, C.; Kümmel, M.; Dietz, K.; Wolz, C. Evaluation of Intraspecies Interference Due to agr Polymorphism in Staphylococcus aureus during Infection and Colonization. J. Infect. Dis. 2003, 188, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Goerke, C.; Wolz, C. Regulatory and genomic plasticity of Staphylococcus aureus during persistent colonization and infection. Int. J. Med. Microbiol. 2004, 294, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, J.; Holden, M.T.G. Understanding the rise of the superbug: Investigation of the evolution and genomic variation of Staphylococcus aureus. Funct. Integr. Genom. 2006, 6, 186–201. [Google Scholar] [CrossRef]
- Peacock, S.J.; Moore, C.; Justice, A.; Kantzanou, M.; Story, L.; Mackie, K.; O’Neill, G.; Day, N.P.J. Virulent Combinations of Adhesin and Toxin Genes in Natural Populations of Staphylococcus aureus. Infect. Immun. 2002, 70, 4987–4996. [Google Scholar] [CrossRef]
- McCarthy, A.J.; Lindsay, J.A. The distribution of plasmids that carry virulence and resistance genes in Staphylococcus aureus is lineage associated. BMC Microbiol. 2012, 12, 104. [Google Scholar] [CrossRef]
- Lindsay, J.A. Staphylococcus aureus genomics and the impact of horizontal gene transfer. Int. J. Med. Microbiol. 2014, 304, 103–109. [Google Scholar] [CrossRef]
- Knöppel, A.; Lind, P.; Lustig, U.; Näsvall, J.; Andersson, D.I. Minor Fitness Costs in an Experimental Model of Horizontal Gene Transfer in Bacteria. Mol. Biol. Evol. 2014, 31, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- de Haas, C.J.; Veldkamp, K.E.; Peschel, A.; Weerkamp, F.; Van Wamel, W.J.; Heezius, E.C.; Poppelier, M.J.; Van Kessel, K.P.; Van Strijp, J.A. Chemotaxis inhibitory protein of Staphylococcus aureus, a bacterial antiinflammatory agent. J. Exp. Med. 2004, 199, 687–695. [Google Scholar] [CrossRef]
- van Wamel, W.J.; Rooijakkers, S.H.; Ruyken, M.; van Kessel, K.P.; van Strijp, J.A. The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on beta-hemolysin-converting bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef]
- Ye, X.; Wang, X.; Fan, Y.; Peng, Y.; Li, L.; Li, S.; Huang, J.; Yao, Z.; Chen, S. Genotypic and Phenotypic Markers of Livestock-Associated Methicillin-Resistant Staphylococcus aureus CC9 in Humans. Appl. Environ. Microbiol. 2016, 82, 3892–3899. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.M.-L.; Lloyd, D.H.; Lindsay, J.A. Staphylococcus aureus host specificity: Comparative genomics of human versus animal isolates by multi-strain microarray. Microbiology 2008, 154, 1949–1959. [Google Scholar] [CrossRef]
- Goerke, C.; Köller, J.; Wolz, C. Ciprofloxacin and Trimethoprim Cause Phage Induction and Virulence Modulation in Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Ubeda, C.; Maiques, E.; Knecht, E.; Lasa, I.; Novick, R.; Penadés, J.R. Antibiotic-induced SOS response promotes horizontal dissemination of pathogenicity island-encoded virulence factors in staphylococci. Mol. Microbiol. 2005, 56, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Wagner, P.L.; Waldor, M.K. Bacteriophage Control of Bacterial Virulence. Infect. Immun. 2002, 70, 3985–3993. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; McCann, A.; Litrup, E.; Murphy, R.; Cormican, M.; Fanning, S.; Brown, D.; Guttman, D.S.; Brisse, S.; Achtman, M. Neutral Genomic Microevolution of a Recently Emerged Pathogen, Salmonella enterica Serovar Agona. PLoS Genet. 2013, 9, e1003471. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Falkow, S. Molecular Koch’s postulates applied to microbial pathogenicity. Rev. Infect. Dis. 1988, 10 (Suppl. 2), S274–S276. [Google Scholar] [CrossRef]
- Laabei, M.; Uhlemann, A.-C.; Lowy, F.D.; Austin, E.D.; Yokoyama, M.; Ouadi, K.; Feil, E.; Thorpe, H.A.; Williams, B.; Perkins, M.; et al. Evolutionary Trade-Offs Underlie the Multi-faceted Virulence of Staphylococcus aureus. PLoS Biol. 2015, 13, e1002229. [Google Scholar] [CrossRef]
- Gillet, Y.; Issartel, B.; Vanhems, P.; Fournet, J.-C.; Lina, G.; Bes, M.; Vandenesch, F.; Piémont, Y.; Brousse, N.; Floret, D.; et al. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet 2002, 359, 753–759. [Google Scholar] [CrossRef]
- Shallcross, L.J.; Fragaszy, E.; Johnson, A.M.; Hayward, A. The role of the Panton-Valentine leucocidin toxin in staphylococcal disease: A systematic review and meta-analysis. Lancet Infect. Dis. 2013, 13, 43–54. [Google Scholar] [CrossRef]
- Sharma, P.; Gupta, S.; Rolain, J.-M. Whole genome sequencing of bacteria in cystic fibrosis as a model for bacterial genome adaptation and evolution. Expert Rev. Anti-Infect. Ther. 2014, 12, 343–355. [Google Scholar] [CrossRef]
- Fournier, P.-E.; Drancourt, M.; Raoult, D. Bacterial genome sequencing and its use in infectious diseases. Lancet Infect. Dis. 2007, 7, 711–723. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed]
- Price, J.; Gordon, N.C.; Crook, D.; Llewelyn, M.; Paul, J. The usefulness of whole genome sequencing in the management of Staphylococcus aureus infections. Clin. Microbiol. Infect. 2013, 19, 784–789. [Google Scholar] [CrossRef]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of Methicillin-Resistant Staphylococcus aureus in a University Hospital Setting by Using Novel Software for spa Repeat Determination and Database Management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef]
- Kahl, B.C.; Mellmann, A.; Deiwick, S.; Peters, G.; Harmsen, D. Variation of the Polymorphic Region X of the Protein A Gene during Persistent Airway Infection of Cystic Fibrosis Patients Reflects Two Independent Mechanisms of Genetic Change in Staphylococcus aureus. J. Clin. Microbiol. 2005, 43, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Mellmann, A.; Weniger, T.; Berssenbrügge, C.; Rothgänger, J.; Sammeth, M.; Stoye, J.; Harmsen, D. Based Upon Repeat Pattern (BURP): An algorithm to characterize the long-term evolution of Staphylococcus aureus populations based on spa polymorphisms. BMC Microbiol. 2007, 7, 98. [Google Scholar] [CrossRef] [PubMed]
- Enright, M.C.; Spratt, B.G. Multilocus sequence typing. Trends Microbiol. 1999, 7, 482–487. [Google Scholar] [CrossRef]
- Enright, M.; Day, N.P.J.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus Sequence Typing for Characterization of Methicillin-Resistant and Methicillin-Susceptible Clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38. [Google Scholar] [CrossRef]
- Foster, T.J. The Staphylococcus aureus “superbug”. J. Clin. Investig. 2004, 114, 1693–1696. [Google Scholar] [CrossRef]
- Frénay, H.M.E.; Bunschoten, A.E.; Schouls, L.M.; Van Leeuwen, W.J.; Vandenbroucke-Grauls, C.M.J.E.; Verhoef, J.; Mooi, F.R. Molecular typing of methicillin-resistant Staphylococcus aureus on the basis of protein A gene polymorphism. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 60–64. [Google Scholar] [CrossRef]
- Robinson, D.; Enright, M. Multilocus sequence typing and the evolution of methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2004, 10, 92–97. [Google Scholar] [CrossRef]
- Mellmann, A.; Andersen, P.S.; Bletz, S.; Friedrich, A.W.; Kohl, T.A.; Lilje, B.; Niemann, S.; Prior, K.; Rossen, J.; Harmsen, D. High Interlaboratory Reproducibility and Accuracy of Next-Generation-Sequencing-Based Bacterial Genotyping in a Ring Trial. J. Clin. Microbiol. 2017, 55, 908–913. [Google Scholar] [CrossRef]
- Leopold, S.R.; Goering, R.V.; Witten, A.; Harmsen, D.; Mellmann, A. Bacterial whole-genome sequencing revisited: Portable, scalable, and standardized analysis for typing and detection of virulence and antibiotic resistance genes. J. Clin. Microbiol. 2014, 52, 2365–2370. [Google Scholar] [CrossRef] [PubMed]
- Blanc, D.S.; Magalhães, B.; Koenig, I.; Senn, L.; Grandbastien, B. Comparison of Whole Genome (wg-) and Core Genome (cg-) MLST (BioNumerics(TM)) Versus SNP Variant Calling for Epidemiological Investigation of Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 1729. [Google Scholar] [CrossRef] [PubMed]
- McAdam, P.R.; Holmes, A.; Templeton, K.; Fitzgerald, J.R. Adaptive Evolution of Staphylococcus aureus during Chronic Endobronchial Infection of a Cystic Fibrosis Patient. PLoS ONE 2011, 6, e24301. [Google Scholar] [CrossRef]
- Salgado-Pabón, W.; Herrera, A.; Vu, B.G.; Stach, C.S.; Merriman, J.A.; Spaulding, A.R.; Schlievert, P.M. Staphylococcus aureus beta-toxin production is common in strains with the beta-toxin gene inactivated by bacteriophage. J. Infect. Dis. 2014, 210, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Long, D.R.; Wolter, D.J.; Lee, M.; Precit, M.; McLean, K.; Holmes, E.; Penewit, K.; Waalkes, A.; Hoffman, L.R.; Salipante, S.J. Polyclonality, Shared Strains, and Convergent Evolution in Chronic Cystic Fibrosis Staphylococcus aureus Airway Infection. Am. J. Respir. Crit. Care Med. 2021, 203, 1127–1137. [Google Scholar] [CrossRef]
- Westphal, C.; Görlich, D.; Kampmeier, S.; Herzog, S.; Braun, N.; Hitschke, C.; Mellmann, A.; Peters, G.; Kahl, B.C.; Staphylococcal CF Study Group; et al. Antibiotic Treatment and Age Are Associated With Staphylococcus aureus Carriage Profiles During Persistence in the Airways of Cystic Fibrosis Patients. Front. Microbiol. 2020, 11, 230. [Google Scholar] [CrossRef] [PubMed]
- Besier, S.; Zander, J.; Kahl, B.C.; Kraiczy, P.; Brade, V.; Wichelhaus, T.A. The Thymidine-Dependent Small-Colony-Variant Phenotype Is Associated with Hypermutability and Antibiotic Resistance in Clinical Staphylococcus aureus Isolates. Antimicrob. Agents Chemother. 2008, 52, 2183–2189. [Google Scholar] [CrossRef]
- Oliver, A.; Cantón, R.; Campo, P.; Baquero, F.; Blazquez, J. High Frequency of Hypermutable Pseudomonas aeruginosa in Cystic Fibrosis Lung Infection. Science 2000, 288, 1251–1253. [Google Scholar] [CrossRef] [PubMed]
- Prunier, A.; Malbruny, B.; Laurans, M.; Brouard, J.; Duhamel, J.; Leclercq, R. High Rate of Macrolide Resistance in Staphylococcus aureus Strains from Patients with Cystic Fibrosis Reveals High Proportions of Hypermutable Strains. J. Infect. Dis. 2003, 187, 1709–1716. [Google Scholar] [CrossRef] [PubMed]
- Tkadlec, J.; Vařeková, E.; Pantůček, R.; Doškař, J.; Růžičková, V.; Botka, T.; Fila, L.; Melter, O. Characterization of Staphylococcus aureus Strains Isolated from Czech Cystic Fibrosis Patients: High Rate of Ribosomal Mutation Conferring Resistance to MLS(B) Antibiotics as a Result of Long-Term and Low-Dose Azithromycin Treatment. Microb. Drug. Resist. 2015, 21, 416–423. [Google Scholar] [CrossRef]
- Ankrum, A.; Hall, B.G. Population Dynamics of Staphylococcus aureus in Cystic Fibrosis Patients To Determine Transmission Events by Use of Whole-Genome Sequencing. J. Clin. Microbiol. 2017, 55, 2143–2152. [Google Scholar] [CrossRef] [PubMed]
- Lange, J.; Heidenreich, K.; Higelin, K.; Dyck, K.; Marx, V.; Reichel, C.; Wamel, W.V.; Reijer, M.D.; Görlich, D.; Kahl, B.C. Staphylococcus aureus Pathogenicity in Cystic Fibrosis Patients-Results from an Observational Prospective Multicenter Study Concerning Virulence Genes, Phylogeny, and Gene Plasticity. Toxins 2020, 12, 279. [Google Scholar] [CrossRef]
- Azarian, T.; Ridgway, J.P.; Yin, Z.; David, M. Long-Term Intrahost Evolution of Methicillin Resistant Staphylococcus aureus Among Cystic Fibrosis Patients With Respiratory Carriage. Front. Genet. 2019, 10, 546. [Google Scholar] [CrossRef]
- Langhanki, L.; Berger, P.; Treffon, J.; Catania, F.; Kahl, B.C.; Mellmann, A. In vivo competition and horizontal gene transfer among distinct Staphylococcus aureus lineages as major drivers for adaptational changes during long-term persistence in humans. BMC Microbiol. 2018, 18, 152. [Google Scholar] [CrossRef]
- Herzog, S.; Dach, F.; De Buhr, N.; Niemann, S.; Schlagowski, J.; Chaves-Moreno, D.; Neumann, C.; Goretzko, J.; Schwierzeck, V.; Mellmann, A.; et al. High Nuclease Activity of Long Persisting Staphylococcus aureus Isolates Within the Airways of Cystic Fibrosis Patients Protects Against NET-Mediated Killing. Front. Immunol. 2019, 10, 2552. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Berends, E.T.; Horswill, A.R.; Haste, N.M.; Monestier, M.; Nizet, V.; Von Köckritz-Blickwede, M. Nuclease Expression by Staphylococcus aureus Facilitates Escape from Neutrophil Extracellular Traps. J. Innate Immun. 2010, 2, 576–586. [Google Scholar] [CrossRef]
- Schilcher, K.; Andreoni, F.; Uchiyama, S.; Ogawa, T.; Schuepbach, R.; Zinkernagel, A.S. Increased Neutrophil Extracellular Trap–Mediated Staphylococcus aureus Clearance Through Inhibition of Nuclease Activity by Clindamycin and Immunoglobulin. J. Infect. Dis. 2014, 210, 473–482. [Google Scholar] [CrossRef]
- Giraudo, A.T.; Cheung, A.L.; Nagel, R. The sae locus of Staphylococcus aureus controls exoprotein synthesis at the transcriptional level. Arch. Microbiol. 1997, 168, 53–58. [Google Scholar] [CrossRef]
- Rogasch, K.; Rühmling, V.; Pané-Farré, J.; Hoöper, D.; Weinberg, C.; Fuchs, S.; Schmudde, M.; Bröker, B.; Wolz, C.; Hecker, M.; et al. Influence of the Two-Component System SaeRS on Global Gene Expression in Two Different Staphylococcus aureus Strains. J. Bacteriol. 2006, 188, 7742–7758. [Google Scholar] [CrossRef]
- Sabirova, J.S.; Hernalsteens, J.-P.; De Backer, S.; Xavier, B.B.; Moons, P.; Turlej-Rogacka, A.; De Greve, H.; Goossens, H.; Malhotra-Kumar, S. Fatty acid kinase A is an important determinant of biofilm formation in Staphylococcus aureus USA300. BMC Genom. 2015, 16, 861. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, I.; Kriegeskorte, A.; Fischer, A.; Deiwick, S.; Theimann, N.; Proctor, R.A.; Peters, G.; Herrmann, M.; Kahl, B.C. In vivo mutations of thymidylate synthase (thyA) are responsible for thymidine-dependency in clinical small colony variants (TD-SCVs) of Staphylococcus aureus. J. Bacteriol. 2008, 190, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Besier, S.; Ludwig, A.; Ohlsen, K.; Brade, V.; Wichelhaus, T.A. Molecular analysis of the thymidine-auxotrophic small colony variant phenotype of Staphylococcus aureus. Int. J. Med. Microbiol. 2007, 297, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Haim, M.S.; Zaheer, R.; Bharat, A.; Di Gregorio, S.; Di Conza, J.; Galanternik, L.; Lubovich, S.; Golding, G.R.; Graham, M.R.; Van Domselaar, G.; et al. Comparative genomics of ST5 and ST30 methicillin-resistant Staphylococcus aureus sequential isolates recovered from paediatric patients with cystic fibrosis. Microb. Genom. 2021, 7, 000510. [Google Scholar] [CrossRef]
- Garofalo, A.; Giai, C.; Lattar, S.; Gardella, N.; Mollerach, M.; Kahl, B.; Becker, K.; Prince, A.S.; Sordelli, D.O.; Gómez, M.I. The Length of the Staphylococcus aureus Protein A Polymorphic Region Regulates Inflammation: Impact on Acute and Chronic Infection. J. Infect. Dis. 2012, 206, 81–90. [Google Scholar] [CrossRef]
- Ormerod, K.L.; George, N.M.; Fraser, J.; Wainwright, C.; Hugenholtz, P. Comparative genomics of non-pseudomonal bacterial species colonising paediatric cystic fibrosis patients. PeerJ 2015, 3, e1223. [Google Scholar] [CrossRef]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef]
- Hanssen, A.M.; Ericson Sollid, J.U. SCCmec in staphylococci: Genes on the move. FEMS Immunol. Med. Microbiol. 2006, 46, 8–20. [Google Scholar] [CrossRef]
- Vandenesch, F.; Naimi, T.; Enright, M.; Lina, G.; Nimmo, G.R.; Heffernan, H.; Liassine, N.; Bes, M.; Greenland, T.; Reverdy, M.-E.; et al. Community-Acquired Methicillin-Resistant Staphylococcus aureus Carrying Panton-Valentine Leukocidin Genes: Worldwide Emergence. Emerg. Infect. Dis. 2003, 9, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Treffon, J.; Fotiadis, S.A.; Van Alen, S.; Becker, K.; Kahl, B.C. The Virulence Potential of Livestock-Associated Methicillin-Resistant Staphylococcus aureus Cultured from the Airways of Cystic Fibrosis Patients. Toxins 2020, 12, 360. [Google Scholar] [CrossRef] [PubMed]
- Dodémont, M.; Argudín, M.; Willekens, J.; Vanderhelst, E.; Piérard, D.; Deyi, V.M.; Hanssens, L.; Franckx, H.; Schelstraete, P.; Leroux-Roels, I.; et al. Emergence of livestock-associated MRSA isolated from cystic fibrosis patients: Result of a Belgian national survey. J. Cyst. Fibros. 2018, 18, 86–93. [Google Scholar] [CrossRef]
- Porterfield, H.S.; Maakestad, L.J.; LaMarche, M.M.; Thurman, A.L.; Kienenberger, Z.E.; Pitcher, N.J.; Hansen, A.R.; Zirbes, C.F.; Boyken, L.; Muyskens, B.L.; et al. MRSA strains with distinct accessory genes predominate at different ages in cystic fibrosis. Pediatr. Pulmonol. 2021, 56, 2868–2878. [Google Scholar] [CrossRef]
- Shen, J.; Wang, Y.; Schwarz, S. Presence and dissemination of the multiresistance gene cfr in Gram-positive and Gram-negative bacteria. J. Antimicrob. Chemother. 2013, 68, 1697–1706. [Google Scholar] [CrossRef]
- Antonelli, A.; D’Andrea, M.M.; Galano, A.; Borchi, B.; Brenciani, A.; Vaggelli, G.; Cavallo, A.; Bartoloni, A.; Giovanetti, E.; Rossolini, G.M. Linezolid-resistant cfr-positive MRSA, Italy. J. Antimicrob. Chemother. 2016, 71, 2349–2351. [Google Scholar] [CrossRef] [PubMed]
- Rouard, C.; Garnier, F.; Leraut, J.; Lepainteur, M.; Rahajamananav, L.; Languepin, J.; Ploy, M.-C.; Bourgeois-Nicolaos, N.; Doucet-Populaire, F. Emergence and Within-Host Genetic Evolution of Methicillin-Resistant Staphylococcus aureus Resistant to Linezolid in a Cystic Fibrosis Patient. Antimicrob. Agents Chemother. 2018, 62, e00720-18. [Google Scholar] [CrossRef] [PubMed]
- Boudet, A.; Jay, A.; Dunyach-Remy, C.; Chiron, R.; Lavigne, J.-P.; Marchandin, H. In-Host Emergence of Linezolid Resistance in a Complex Pattern of Toxic Shock Syndrome Toxin-1-Positive Methicillin-Resistant Staphylococcus aureus Colonization in Siblings with Cystic Fibrosis. Toxins 2021, 13, 317. [Google Scholar] [CrossRef] [PubMed]
- Roch, M.; Varela, M.C.; Taglialegna, A.; Rosato, A.E. Tedizolid is a promising antimicrobial option for the treatment of Staphylococcus aureus infections in cystic fibrosis patients. J. Antimicrob. Chemother. 2019, 75, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Rolain, J.-M.; François, P.; Hernandez, D.; Bittar, F.; Richet, H.; Fournous, G.; Mattenberger, Y.; Bosdure, E.; Stremler, N.; Dubus, J.-C.; et al. Genomic analysis of an emerging multiresistant Staphylococcus aureus strain rapidly spreading in cystic fibrosis patients revealed the presence of an antibiotic inducible bacteriophage. Biol. Direct 2009, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- López-Collazo, E.; Jurado, T.; de Dios Caballero, J.; Pérez-Vázquez, M.; Vindel, A.; Hernández-Jiménez, E.; Tamames, J.; Cubillos-Zapata, C.; Manrique, M.; Tobes, R.; et al. In vivo attenuation and genetic evolution of a ST247-SCCmecI MRSA clone after 13 years of pathogenic bronchopulmonary colonization in a patient with cystic fibrosis: Implications of the innate immune response. Mucosal. Immunol. 2015, 8, 362–371. [Google Scholar] [CrossRef]
- Long, S.W.; Olsen, R.J.; Mehta, S.C.; Palzkill, T.; Cernoch, P.L.; Perez, K.; Musick, W.L.; Rosato, A.E.; Musser, J.M. PBP2a Mutations Causing High-Level Ceftaroline Resistance in Clinical Methicillin-Resistant Staphylococcus aureus Isolates. Antimicrob. Agents Chemother. 2014, 58, 6668–6674. [Google Scholar] [CrossRef]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival Mechanisms of Clinically Relevant Microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef]
- McKenney, D.; Pouliot, K.L.; Wang, Y.; Murthy, V.; Ulrich, M.; Döring, G.; Lee, J.C.; Goldmann, D.A.; Pier, G.B. Broadly Protective Vaccine for Staphylococcus aureus Based on an in Vivo-Expressed Antigen. Science 1999, 284, 1523–1527. [Google Scholar] [CrossRef]
- Li, Z.; Kosorok, M.R.; Farrell, P.M.; Laxova, A.; West, S.E.; Green, C.G.; Collins, J.; Rock, M.J.; Splaingard, M.L. Longitudinal development of mucoid Pseudomonas aeruginosa infection and lung disease progression in children with cystic fibrosis. JAMA 2005, 293, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Schwartbeck, B.; Birtel, J.; Treffon, J.; Langhanki, L.; Mellmann, A.; Kale, D.; Kahl, J.; Hirschhausen, N.; Neumann, C.; Lee, J.; et al. Dynamic in vivo mutations within the ica operon during persistence of Staphylococcus aureus in the airways of cystic fibrosis patients. PLoS Pathog. 2016, 12, e1006024. [Google Scholar] [CrossRef]
- Gabryszewski, S.J.; Lung, T.W.F.; Annavajhala, M.; Tomlinson, K.; Riquelme, S.A.; Khan, I.; Noguera, L.; Wickersham, M.; Zhao, A.; Mulenos, A.M.; et al. Metabolic Adaptation in Methicillin-Resistant Staphylococcus aureus Pneumonia. Am. J. Respir. Cell Mol. Biol. 2019, 61, 185–197. [Google Scholar] [CrossRef]
- Bernardy, E.E.; Petit, R.A., III; Moller, A.G.; Blumenthal, J.A.; McAdam, A.J.; Priebe, G.P.; Chande, A.T.; Rishishwar, L.; Jordan, I.K.; Read, T.D.; et al. Whole-Genome Sequences of Staphylococcus aureus Isolates from Cystic Fibrosis Lung Infections. Microbiol. Resour. Announc. 2019, 8, e01564-18. [Google Scholar] [CrossRef] [PubMed]
- Bernardy, E.E.; Petit, R.A., III; Raghuram, V.; Alexander, A.M.; Read, T.D.; Goldberg, J.B. Genotypic and Phenotypic Diversity of Staphylococcus aureus Isolates from Cystic Fibrosis Patient Lung Infections and Their Interactions with Pseudomonas aeruginosa. mBio 2020, 11, e00735-20. [Google Scholar] [CrossRef]
- Lennartz, F.E.; Schwartbeck, B.; Dübbers, A.; Große-Onnebrink, J.; Kessler, C.; Küster, P.; Schültingkemper, H.; Peters, G.; Kahl, B.C. The prevalence of Staphylococcus aureus with mucoid phenotype in the airways of patients with cystic fibrosis—A prospective study. Int. J. Med Microbiol. 2019, 309, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, K.L.; Lung, T.W.F.; Dach, F.; Annavajhala, M.K.; Gabryszewski, S.J.; Groves, R.A.; Drikic, M.; Francoeur, N.J.; Sridhar, S.H.; Smith, M.L.; et al. Staphylococcus aureus induces an itaconate-dominated immunometabolic response that drives biofilm formation. Nat. Commun. 2021, 12, 1399. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rumpf, C.; Lange, J.; Schwartbeck, B.; Kahl, B.C. Staphylococcus aureus and Cystic Fibrosis—A Close Relationship. What Can We Learn from Sequencing Studies? Pathogens 2021, 10, 1177. https://doi.org/10.3390/pathogens10091177
Rumpf C, Lange J, Schwartbeck B, Kahl BC. Staphylococcus aureus and Cystic Fibrosis—A Close Relationship. What Can We Learn from Sequencing Studies? Pathogens. 2021; 10(9):1177. https://doi.org/10.3390/pathogens10091177
Chicago/Turabian StyleRumpf, Christine, Jonas Lange, Bianca Schwartbeck, and Barbara C. Kahl. 2021. "Staphylococcus aureus and Cystic Fibrosis—A Close Relationship. What Can We Learn from Sequencing Studies?" Pathogens 10, no. 9: 1177. https://doi.org/10.3390/pathogens10091177
APA StyleRumpf, C., Lange, J., Schwartbeck, B., & Kahl, B. C. (2021). Staphylococcus aureus and Cystic Fibrosis—A Close Relationship. What Can We Learn from Sequencing Studies? Pathogens, 10(9), 1177. https://doi.org/10.3390/pathogens10091177