
Chlamydia Psittaci ST24: Clonal Strains of One Health Importance Dominate in Australian Horse, Bird and Human Infections


 , ,
, ,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Hosts, Infecting Strains Diversity and Clinical Signs
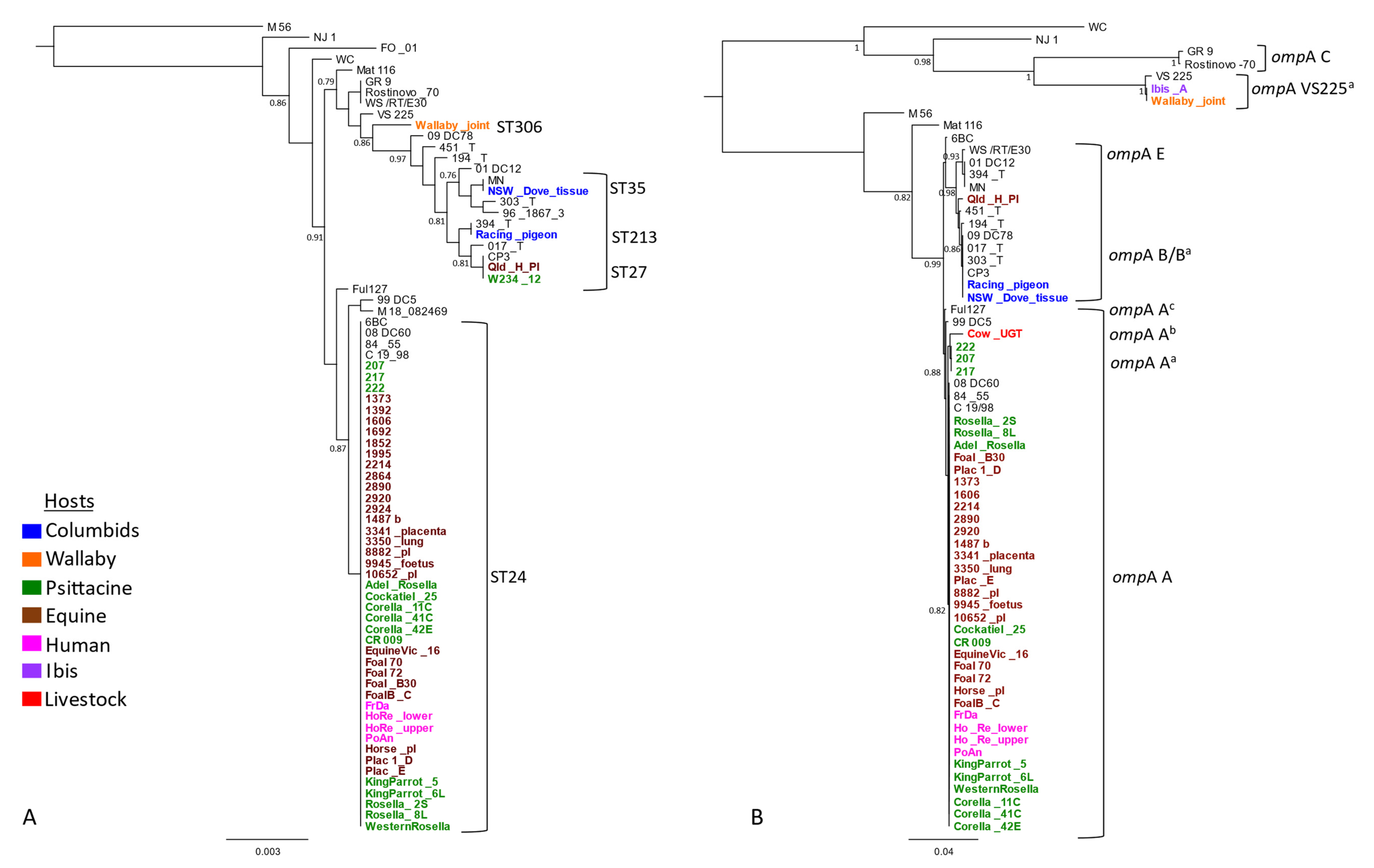
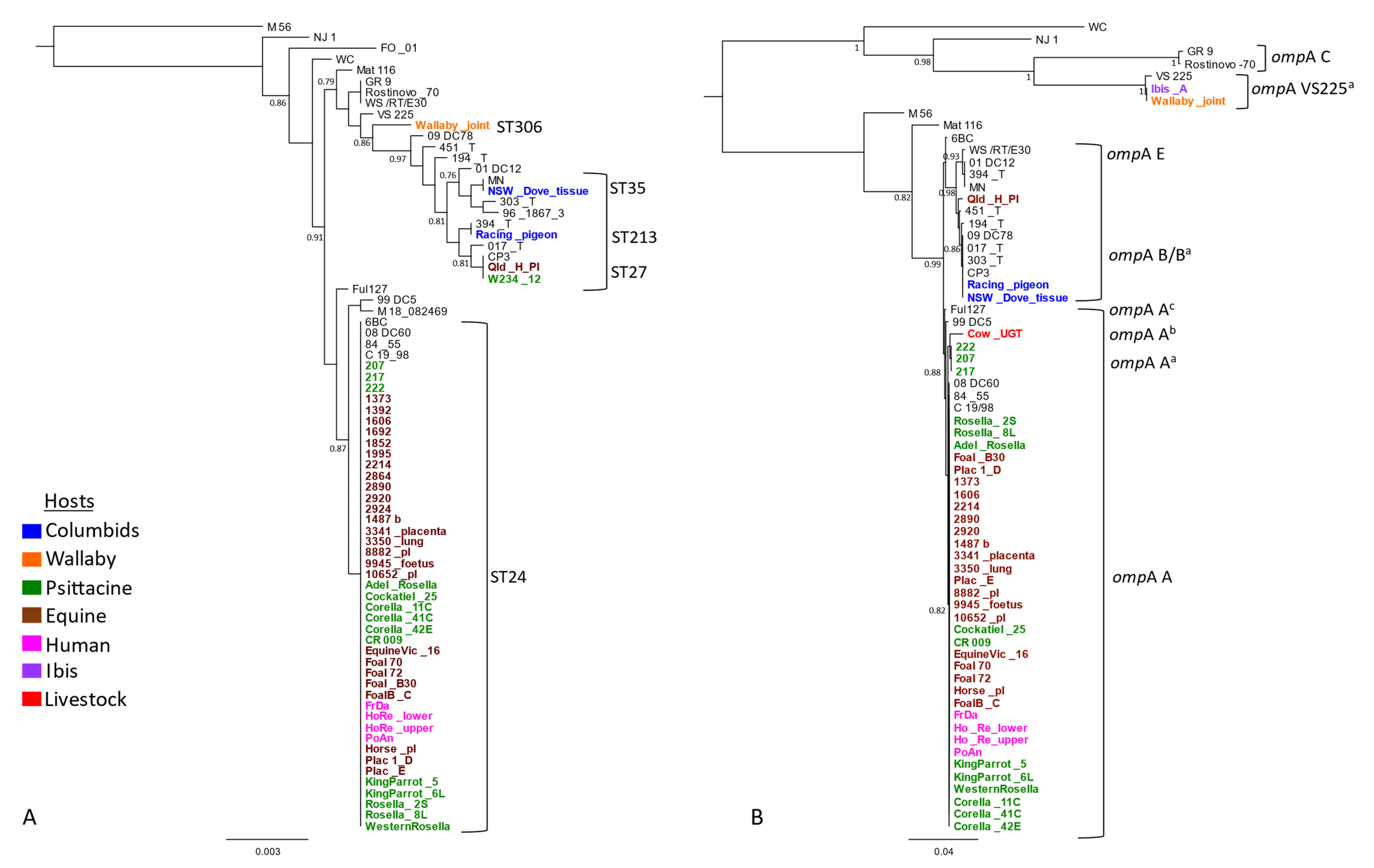
2.2. Phylogenetic Analyses of Detected C. psittaci Strains
3. Discussion
3.1. Uncovering C. psittaci Genetic Diversity
3.2. Roadmap to Understanding the Pathogenicity of Equine Abortogenic C. psittaci
3.3. Are We Missing Other Hosts?
3.4. Diagnostic Directions for C. psittaci Equine Abortion and Zoonosis
4. Materials and Methods
4.1. Sample Collection, DNA Extraction and C. psittaci Screening
4.2. Genotyping
4.3. Phylogenetic and Cluster Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knittler, M.R.; Sachse, K. Chlamydia psittaci: Update on an underestimated zoonotic agent. Pathog. Dis. 2015, 73, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Borel, N.; Polkinghorne, A.; Pospischil, A. A review on chlamydial diseases in animals: Still a challenge for pathologists? Vet. Pathol. 2018, 55, 374–390. [Google Scholar] [CrossRef] [Green Version]
- Zaręba-Marchewka, K.; Szymańska-Czerwińska, M.; Niemczuk, K. Chlamydiae—What’s new? J. Vet. Res. 2020, 64, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Branley, J.; Bachmann, N.L.; Jelocnik, M.; Myers, G.S.; Polkinghorne, A. Australian human and parrot Chlamydia psittaci strains cluster within the highly virulent 6BC clade of this important zoonotic pathogen. Sci. Rep. 2016, 6, 30019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemmerling, K.; Müller, U.; Mielenz, M.; Sauerwein, H. Chlamydophila species in dairy farms: Polymerase chain reaction prevalence, disease association, and risk factors identified in a cross-sectional study in western Germany. J. Dairy Sci. 2009, 92, 4347–4354. [Google Scholar] [CrossRef] [PubMed]
- Lenzko, H.; Moog, U.; Henning, K.; Lederbach, R.; Diller, R.; Menge, C.; Sachse, K.; Sprague, L.D. High frequency of chlamydial co-infections in clinically healthy sheep flocks. BMC Vet. Res. 2011, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Lagae, S.; Kalmar, I.; Laroucau, K.; Vorimore, F.; Vanrompay, D. Emerging Chlamydia psittaci infections in chickens and examination of transmission to humans. J. Med. Microbiol. 2014, 63, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Jelocnik, M.; Laurence, M.; Murdoch, F.R.; Polkinghorne, A. Detection of Chlamydiaceae in ocular swabs from Australian pre-export feedlot sheep. Aust. Vet. J. 2019, 97, 401–403. [Google Scholar] [CrossRef]
- Jelocnik, M.; Taylor-Brown, A.; O’Dea, C.; Anstey, S.; Bommana, S.; Masters, N.; Katouli, M.; Jenkins, C.; Polkinghorne, A. Detection of a range of genetically diverse chlamydiae in Australian domesticated and wild ungulates. Transbound. Emerg. Dis. 2019, 66, 1132–1137. [Google Scholar] [CrossRef] [PubMed]
- Anstey, S.I.; Quigley, B.L.; Polkinghorne, A.; Jelocnik, M. Chlamydial infection and on-farm risk factors in dairy cattle herds in South East Queensland. Aust. Vet. J. 2019, 97, 505–508. [Google Scholar] [CrossRef]
- Stokes, H.S.; Martens, J.M.; Walder, K.; Segal, Y.; Berg, M.L.; Bennett, A.T.D. Species, sex and geographic variation in chlamydial prevalence in abundant wild Australian parrots. Sci. Rep. 2020, 10, 20478. [Google Scholar] [CrossRef]
- Amery-Gale, J.; Legione, A.R.; Marenda, M.S.; Owens, J.; Eden, P.A.; Konsak-Ilievski, B.M.; Whiteley, P.L.; Dobson, E.C.; Browne, E.A.; Slocombe, R.F.; et al. Surveillance for Chlamydia spp. with multilocus sequence typing analysis in wild and captive birds in Victoria Australia. J. Wildl. Dis. 2020, 56, 16–26. [Google Scholar] [CrossRef]
- Jelocnik, M.; Jenkins, C.; O’Rourke, B.; Barnwell, J.; Polkinghorne, A. Molecular evidence to suggest pigeon-type Chlamydia psittaci in association with an equine foal loss. Transbound. Emerg. Dis. 2018, 65, 911–915. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.; Jelocnik, M.; Micallef, M.L.; Galea, F.; Taylor–Brown, A.; Bogema, D.R.; Liu, M.; O’Rourke, B.; Chicken, C.; Carrick, J.; et al. An epizootic of Chlamydia psittaci equine reproductive loss associated with suspected spillover from native Australian parrots. Emerg. Microbes Infect. 2018, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Burnet, F.M. A note of the occurrence of fatal psittacosis in parrots living in the wild state. Med. J. Aust. 1939, 1, 545–546. [Google Scholar] [CrossRef]
- Chan, J.; Doyle, B.; Branley, J.; Sheppeard, V.; Gabor, M.; Viney, K.; Quinn, H.; Janover, O.; McCready, M.; Heller, J. An outbreak of psittacosis at a veterinary school demonstrating a novel source of infection. One Health 2017, 3, 29–33. [Google Scholar] [CrossRef]
- Jelocnik, M.; Branley, J.; Heller, J.; Raidal, S.; Alderson, S.; Galea, F.; Gabor, M.; Polkinghorne, A. Multilocus sequence typing identifies an avian-like Chlamydia psittaci strain involved in equine placentitis and associated with subsequent human psittacosis. Emerg. Microbes Inf. 2017, 6, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anstey, S.; Lizárraga, D.; Nyari, S.; Chalmers, G.; Carrick, J.; Chicken, C.; Jenkins, C.; Perkins, N.; Timms, P.; Jelocnik, M. Epidemiology of Chlamydia psittaci infections in pregnant Thoroughbred mares and foals. Vet. J. 2021, 273, 105683. [Google Scholar] [CrossRef]
- Akter, R.; Sansom, F.M.; El-Hage, C.M.; Gilkerson, J.R.; Legione, A.R.; Devlin, J.M. A 25-year retrospective study of Chlamydia psittaci in association with equine reproductive loss in Australia. J. Med. Microbiol. 2021, 70. [Google Scholar] [CrossRef]
- Sachse, K.; Laroucau, K.; Hotzel, H.; Schubert, E.; Ehricht, R.; Slickers, P. Genotyping of Chlamydophila psittaci using a new DNA microarray assay based on sequence analysis of ompA genes. BMC Microbiol. 2008, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Pannekoek, Y.; Dickx, V.; Beeckman, D.S.; Jolley, K.A.; Keijzers, W.C.; Vretou, E.; Maiden, M.C.; Vanrompay, D.; van der Ende, A. Multi locus sequence typing of Chlamydia reveals an association between Chlamydia psittaci genotypes and host species. PLoS ONE 2010, 5, e14179. [Google Scholar] [CrossRef]
- Vorimore, F.; Aaziz, R.; de Barbeyrac, B.; Peuchant, O.; Szymańska-Czerwińska, M.; Herrmann, B.; Schnee, C.; Laroucau, K. A new SNP-based genotyping method for C. psittaci: Application to field samples for quick identification. Microorganisms 2021, 9, 625. [Google Scholar] [CrossRef] [PubMed]
- Read, T.D.; Joseph, S.J.; Didelot, X.; Liang, B.; Patel, L.; Dean, D. Comparative analysis of Chlamydia psittaci genomes reveals the recent emergence of a pathogenic lineage with a broad host range. mBio 2013, 4, e00604-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akter, R.; Stent, A.W.; Sansom, F.M.; Gilkerson, J.R.; Burden, C.; Devlin, J.M.; Legione, A.R.; El-Hage, C.M. Chlamydia psittaci: A suspected cause of reproductive loss in three Victorian horses. Aust. Vet. J. 2020, 98, 570–573. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, M.; Sarker, S.; Vaz, P.K.; Legione, A.R.; Devlin, J.M.; Macwhirter, P.L.; Whiteley, P.L.; Raidal, S.R. Disease surveillance in wild Victorian cacatuids reveals co-infection with multiple agents and detection of novel avian viruses. Vet. Microbiol. 2019, 235, 257–264. [Google Scholar] [CrossRef]
- Branley, J.M.; Weston, K.M.; England, J.; Dwyer, D.E.; Sorrell, T.C. Clinical features of endemic community-acquired psittacosis. New Microbes New infect. 2014, 2, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Mattmann, P.; Marti, H.; Borel, N.; Jelocnik, M.; Albini, S.; Vogler, B.R. Chlamydiaceae in wild, feral and domestic pigeons in Switzerland and insight into population dynamics by Chlamydia psittaci multilocus sequence typing. PLoS ONE 2019, 14, e0226088. [Google Scholar] [CrossRef] [Green Version]
- Favaroni, A.; Trinks, A.; Weber, M.; Hegemann, J.H.; Schnee, C. Pmp repertoires influence the different infectious potential of avian and mammalian Chlamydia psittaci strains. Front. Microbiol. 2021, 12, 656209. [Google Scholar] [CrossRef]
- Robbins, A.; Hanger, J.; Jelocnik, M.; Quigley, B.L.; Timms, P. Koala immunogenetics and chlamydial strain type are more directly involved in chlamydial disease progression in koalas from two south east Queensland koala populations than koala retrovirus subtypes. Sci. Rep. 2020, 10, 15013. [Google Scholar] [CrossRef]
- Peel, A.J.; Wells, K.; Giles, J.; Boyd, V.; Burroughs, A.; Edson, D.; Crameri, G.; Baker, M.L.; Field, H.; Wang, L.F.; et al. Synchronous shedding of multiple bat paramyxoviruses coincides with peak periods of Hendra virus spillover. Emerg. Microbes Infect. 2019, 8, 1314–1323. [Google Scholar] [CrossRef] [Green Version]
- Field, H.E. Evidence of Australian bat lyssavirus infection in diverse Australian bat taxa. Zoonoses Public Health 2018, 65, 742–748. [Google Scholar] [CrossRef] [Green Version]
- Barr, J.A.; Smith, C.; Marsh, G.A.; Field, H.; Wang, L.F. Evidence of bat origin for Menangle virus, a zoonotic paramyxovirus first isolated from diseased pigs. J. Gen. Virol. 2012, 93, 2590–2594. [Google Scholar] [CrossRef]
- Khusro, A.; Aarti, C.; Pliego, A.B.; Cipriano-Salazar, M. Hendra virus infection in horses: A review on emerging mystery paramyxovirus. J. Equine Vet. Sci. 2020, 91, 103149. [Google Scholar] [CrossRef] [PubMed]
- Williamson, K.M.; Wheeler, S.; Kerr, J.; Bennett, J.; Freeman, P.; Kohlhagen, J.; Peel, A.J.; Eby, P.; Merritt, T.; Housen, T.; et al. Hendra in the Hunter Valley. One Health 2020, 10, 100162. [Google Scholar] [CrossRef] [PubMed]
- Australian Government Department of Agriculture, Water and Environment. National Flying-Fox Monitoring Viewer. Available online: https://www.environment.gov.au/webgis-framework/apps/ffc-wide/ffc-wide.jsf (accessed on 4 June 2021).
- Fritschi, J.; Marti, H.; Seth-Smith, H.M.B.; Aeby, S.; Greub, G.; Meli, M.L.; Hofmann-Lehmann, R.; Mühldorfer, K.; Stokar-Regenscheit, N.; Wiederkehr, D.; et al. Prevalence and phylogeny of Chlamydiae and hemotropic mycoplasma species in captive and free-living bats. BMC Microbiol. 2020, 20, 182. [Google Scholar] [CrossRef]
- Plowright, R.K.; Reaser, J.K.; Locke, H.; Woodley, S.J.; Patz, J.A.; Becker, D.J.; Oppler, G.; Hudson, P.J.; Tabor, G.M. Land use-induced spillover: A call to action to safeguard environmental, animal, and human health. Lancet Planet. 2021, 5, e237–e245. [Google Scholar] [CrossRef]
- Jones, B.; Taylor, K.; Lucas, R.M.; Merritt, T.; Chicken, C.; Heller, J.; Carrick, J.; Givney, R.; Durrheim, D.N. Challenges in using serological methods to explore historical transmission risk of Chlamydia psittaci in a workforce with high exposure to equine chlamydiosis. Commun. Dis. Intell. 2019, 43. [Google Scholar] [CrossRef]
- Taylor, K.A.; Durrheim, D.; Heller, J.; O’Rourke, B.; Hope, K.; Merritt, T.; Freeman, P.; Chicken, C.; Carrick, J.; Branley, J.; et al. Equine chlamydiosis-An emerging infectious disease requiring a one health surveillance approach. Zoonoses Public Health 2018, 65, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Australian Institute of Health and Welfare. National Notifiable Diseases Surveillance System. Available online: http://www9.health.gov.au/cda/source/cda-index.cfm (accessed on 15 February 2021).
- Jelocnik, M.; Islam, M.M.; Madden, D.; Jenkins, C.; Branley, J.; Carver, S.; Polkinghorne, A. Development and evaluation of rapid novel isothermal amplification assays for important veterinary pathogens: Chlamydia psittaci and Chlamydia pecorum. PeerJ 2017, 5, e3799. [Google Scholar] [CrossRef] [Green Version]
- Reaser, J.K.; Witt, A.; Tabor, G.M.; Hudson, P.J.; Plowright, R.K. Ecological countermeasures for preventing zoonotic disease outbreaks: When ecological restoration is a human health imperative. Restor. Ecol. 2021, e13357. [Google Scholar] [CrossRef]
- De Garine-Wichatitsky, M.; Binot, A.; Ward, J.; Caron, A.; Perrotton, A.; Ross, H.; Tran Quoc, H.; Valls-Fox, H.; Gordon, I.J.; Promburom, P.; et al. “Health in” and “Health of” social-ecological systems: A practical framework for the management of healthy and resilient agricultural and natural ecosystems. Front. Public Health 2020, 8, 616328. [Google Scholar] [CrossRef]
- Madico, G.; Gursky, O.; Fairman, J.; Massari, P. Structural and immunological characterization of novel recombinant MOMP-based chlamydial antigens. Vaccines 2017, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Pantchev, A.; Sting, R.; Bauerfeind, R.; Tyczka, J.; Sachse, K. Detection of all Chlamydophila and Chlamydia spp. of veterinary interest using species-specific real-time PCR assays. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Jelocnik, M.; Polkinghorne, A.; Pannekoek, Y. Multilocus sequence typing (MLST) of Chlamydiales. Methods Mol. Biol. 2019, 2042, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Geneious Prime 2020.2.4. Available online: https://www.geneious.com (accessed on 3 February 2021).
- Chlamydiales MLST Database. Available online: https://pubmlst.org/organisms/chlamydiales (accessed on 3 February 2021).
- BLASTn. Available online: https://blast.ncbi.nlm.nih.gov (accessed on 20 March 2021).
- Tonkin-Hill, G.; Lees, J.A.; Bentley, S.D.; Frost, S.D.W.; Corander, J. Fast hierarchical Bayesian analysis of population structure. Nucleic Acids Res. 2019, 47, 5539–5549. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Hosts | Avian | Equine | Human | Wallaby | |
|---|---|---|---|---|---|
| Psittacine | Columbids | ||||
| No. of ST24-positive hosts | 19/20 (95%) | 0/2 (0%) | 44/45 (97.8%) | 5/5 (100%) | 0/1 (0%) |
| Total No. of hosts with clinical signs in ST24 infections | 11/19 (57.9%) | - | 44/44 (100%) | 4/5 (80%) | - |
| No. of other STs-positive hosts | 1/20 (5%) | 2/2(100%) | 1/45 (2.2%) | 0/6 (0%) | 1/1 (100%) |
| Total No. of hosts with clinical signs in other STs infections | 0/1 (0%) | 2/2(100%) | 1/1 (100%) | - | 1/1 (100%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anstey, S.I.; Kasimov, V.; Jenkins, C.; Legione, A.; Devlin, J.; Amery-Gale, J.; Gilkerson, J.; Hair, S.; Perkins, N.; Peel, A.J.; et al. Chlamydia Psittaci ST24: Clonal Strains of One Health Importance Dominate in Australian Horse, Bird and Human Infections. Pathogens 2021, 10, 1015. https://doi.org/10.3390/pathogens10081015
Anstey SI, Kasimov V, Jenkins C, Legione A, Devlin J, Amery-Gale J, Gilkerson J, Hair S, Perkins N, Peel AJ, et al. Chlamydia Psittaci ST24: Clonal Strains of One Health Importance Dominate in Australian Horse, Bird and Human Infections. Pathogens. 2021; 10(8):1015. https://doi.org/10.3390/pathogens10081015
Chicago/Turabian StyleAnstey, Susan I., Vasilli Kasimov, Cheryl Jenkins, Alistair Legione, Joanne Devlin, Jemima Amery-Gale, James Gilkerson, Sam Hair, Nigel Perkins, Alison J. Peel, and et al. 2021. "Chlamydia Psittaci ST24: Clonal Strains of One Health Importance Dominate in Australian Horse, Bird and Human Infections" Pathogens 10, no. 8: 1015. https://doi.org/10.3390/pathogens10081015
APA StyleAnstey, S. I., Kasimov, V., Jenkins, C., Legione, A., Devlin, J., Amery-Gale, J., Gilkerson, J., Hair, S., Perkins, N., Peel, A. J., Borel, N., Pannekoek, Y., Chaber, A.-L., Woolford, L., Timms, P., & Jelocnik, M. (2021). Chlamydia Psittaci ST24: Clonal Strains of One Health Importance Dominate in Australian Horse, Bird and Human Infections. Pathogens, 10(8), 1015. https://doi.org/10.3390/pathogens10081015